Identification and Characterization of an Agarase- and Xylanse-producing Catenovulum jejuensis A28-5 from Coastal Seawater of Jeju Island, Korea
Da Som Kim, Ga Ram Jeong, Chang Hwan Bae, Joo-Hong Yeo, and Won-Jae Chi*
Biological and Genetic Resources Assessment Division, National Institute of Biological Resources, Incheon 22689, Republic of Korea
Received: March 31, 2017 / Revised: June 8, 2017 / Accepted: June 8, 2017
Introduction
Plant biomass is the most abundant and widely spread source of bioproducts and biofuels. Hydrolysis of the plant biomass is the most important step for extract- ing marine plant-derived polysaccharides (e.g., agar, carrageenan, and alginate) and terrestrial plant-derived polysaccharides (e.g., cellulose, and hemicelluloses), which exist in enormous amounts in nature. During hydrolysis, distinctive oligosaccharides with various physiological activities are produced [1].
Agar is composed of agaropectin and agarose, which are the main constituents of the cell wall of macroalgae.
Agarose is a polymer that is linked by repeating units of 3,6-anhydro-α-L-galactose and β-D-galactose. Depend- ing on the method of degradation, agarooligosaccharide (produced by α-agarase or acid hydrolysis) and neoaga- rooligosaccharide (produced by β-agarase) are produced.
These oligosaccharides can be used in various indus- tries, including the food industry, for properties such as temperature-stability, intestinal regulation, and low cal- ories; the pharmaceutical industry, for their therapeutic and preventive effects against obesity and diabetes; and the cosmetic industry, for their whitening, moisturizing, and antioxidative effects [2].
Xylan is a hemicellulose, the main constituent that accounts for 20−40% of the cell wall constituents of ter- restrial plants, and has a complex structure composed of β-1,4-linked xylose monomers substituted with arabino- syl, acetyl, and glucuronosyl groups and can be hydro- Strain A28-5, which can degrade xylan and agar in solid medium, was isolated from a coastal seawater sample collected from Jeju Island, South Korea. This strain was found to be a gram-negative, Na+-requiring bacte- rial strain with a polar flagellum for motility. Additionally, the strain was tolerant to antibiotics such as ampicillin and thiostrepton. The G+C content of the genome was 43.96% and menaquinone-7 was found to be the predominant quinone. Major fatty acids constituting the cell wall of the strain were C16:1ω7c/iso-C15:0
2-OH (23.32%), C16:0 (21.83%), and C18:1ω7c (17.98%). The 16S rRNA gene sequence of the strain showed the highest similarity (98.94%) to that of Catenovulum agarivorans YM01, which was demonstrated by construct- ing a neighbor-joining phylogenetic tree. A28-5 was identified as a novel species of the genus Catenovulum via DNA-DNA hybridization with Catenovulum agarivorans YM01, and thus was named as Catenovulum jejuensis A28-5. The formation of tetramers and hexamers of xylooligosaccharides and (neo)agarooligosac- charides, respectively, were confirmed by thin-layer chromatography analysis using an enzyme reaction solution containing xylan or agarose with two crude enzymes prepared from the liquid culture of the strain.
Keywords: Agarase, xylanase, Catenovulum, characterization, xylooligosaccharide, agarooligosaccharide
*Corresponding author
Tel: +82-32-590-7113, Fax: +82-32-590-7472 E-mail: [email protected]
© 2017, The Korean Society for Microbiology and Biotechnology
lyzed by various xylanases. Xylanases are required in various processes such as in pulp-prebleaching to remove the hemicelluloses in pulp, in stimulation of digestion of animal feed, as food and baking additives, in processing of fruits and vegetables, in ethanol and xylitol production, and in paper manufacturing. Research on the properties of xylanases that are suitable for use in each industry is essential. Furthermore, xylooligosac- charides (XOSs), the xylan hydrolysates produced by xylanases, are known to have various physiological activities, which include antimicrobial, antioxidative, and anti-inflammatory activities; thus, it is believed that they can be used in various industries such as food, med- ical, agriculture, livestock, and cosmetic [3, 4].
The family Altermonadaceae within the class δ- proteobacteria are large group of marine, obligate aerobic heterotroph and requiring sodium to grow. They have genomes that contain several macromolecule-degradable genes and secondary metabolite synthetic gene clusters [5]. Catenovulum is a novel genus belonging to the class δ-proteobacteria. Since the identification of the novel species, C. agarivorans YM01, which was first reported by Yan et al. [6], a new different species C. maritimus Q1 belonging to this genus had been reported until now [7].
The aforementioned hydrolytic degradation methods using enzymes such as agarase and xylanase can selec- tively and specifically hydrolyze polysaccharides to pro- duce desired oligosaccharides. In particular, the hydrolysis of plant biomass by microbial enzymes has been accepted as an eco-friendly method that can replace chemical deg- radation methods, which produce harmful by-products [8]. Many researchers have been trying to isolate new microorganisms that produce more suitable enzymes [9− 12].
In this study, a new microorganism that produce useful enzymes capable of hydrolyzing the cell walls of marine and terrestrial plants were identified and the properties of the enzymes were identified to determine their use in the manufacture of plant biomass-derived oligosaccharides.
Materials and Methods
Isolation of microorganisms
Seawater was collected off the coast of Seogwipo-si,
Jeju Island and diluted to a series of concentrations (10-1−10-5). It was smeared on marine agar (MA) 2216 (MA, Difco, USA) and incubated at 37℃ for 2 d. Colonies formed on the this plate were inoculated on a new culture plate with media containing azurine-crosslinked(AZCL)- xylan (Megazyme, Ireland), and were incubated at 37℃ for 2 d. The colonies exhibiting activation of xylanase or agarase were selected for further studies. Xylanase acti- vation was confirmed from the blue coloration that appeared around the colonies after AZCL-xylan in the media was degraded, and agarase activation was con- firmed after staining with Lugol’s Iodine solution. Among the selected bacterial colonies, those showing activation of both xylanase and agarase simultaneously, were inoc- ulated on a new culture plate and incubated for 2 d.
Finally, one microbial strain was selected through the screening process based on the morphological character- istics such as shape, color, transparency, and size of the colony. The selected strain was named strain A28-5.
Sequencing of 16S rRNA gene sequence of strain A28-5 Strain A28-5 was cultured in marine broth (MB) under shaking conditions for 2 d and the bacteria was harvested by centrifugation at 20,000 ×g for 10 min.
Genomic DNA was extracted from the collected bacterial sample by using a Genomic DNA extraction kit from DyneBio Inc. (Korea). The 16S rRNA gene of the strain A28-5 was amplified using bacterial universal primers (27F and 1492R) [13]. The PCR conditions were (i) 30 s at 94℃, (ii) 30 cycles of 20 s at 98℃, 30 s at 55℃, and 1 min at 72℃, and (iii) 10 min at 72℃. PCR was performed in a Thermal Cycler Dice (Takara, Japan) and the amplified PCR products were analyzed on a 1%
agarose gel in TAE buffer. The DNA fragments were inserted into the pGEM-T easy vector (Promega, USA) and base sequence analysis was performed by Genotech Inc. (Daejeon, Korea). Base sequence homology analysis was performed on the 16S rRNA gene sequence of strain A28-5 with the data from GenBank database using the BlastN program of National Center for Biotechnology Information (NCBI). Additionally, homology with type strains was determined by comparing the 16S rRNA gene sequence of strain A28-5 with that of type strains from EzTaxon database (http://www.ezbiocloud.net/
eztaxon).
Phylogenetic analysis
For phylogenetic analysis, the 16S rRNA gene sequences of type strains provided by EzTaxon database were secured and phylogenetic associations with strain A28-5 were analyzed by creating a phylogenetic tree using PHYLIP (PHYLogeny Inference Package) pro- gram. The ClustalW program was used for multiple alignment between the secured base sequences, and the gaps at the 5′- and 3′-ends were edited by the GeneDoc program. The phylogenetic tree was produced by the neighbor joining (NJ) method [14]. The produced NJ phylogenetic tree was verified by making phylogenetic trees using the maximum parsimony (MP) and maxi- mum likelihood (ML) methods.
DNA-DNA hybridization
To investigate the possibility of strain A28-5 being a new species, DNA-DNA hybridization was performed on the type strains with high phylogenetic relationship, Catenovulum agarivorans YM01 and Gayadomonas joo- biniege G7, at the Korean Culture Center of Microorgan- isms. The method used for the DNA-DNA hybridization analysis is as follows: The bacterial strain was cultured on MA plate and the genomic DNA was extracted.
Micrococcus luteus KCCM 11326 was used as a negative standard bacterial strain. Probe DNA preparation and hybridization reaction were conducted using a DIG High Prime DNA Labeling and Detection Starter Kit II (Roche Applied Science, Germany) according to the man- ufacturer’s instructions. Hybridization signals were measured by the Quantity One Program (Bio-Rad, USA), and the values of type strains were calculated by determining the signal of strain A28-5 as 100%. The hybridization value for each bacterial strain was calcu- lated by performing three repetitive experiments.
Analysis of morphological and physiological characteris- tics of strain A28-5
Strain A28-5 was stained using a Gram stain kit (BD, USA), followed by observation under a microscope. The cells cultured on the plate for 2 d were also stained by 1% phosphotungstic acid (PTA) and were observed under a transmission electron microscope (TEM) (JEM1010, JEOL, Japan) to confirm flagella.
To investigate the growth characteristics of strain A28-5, growth was observed by culturing and incubating
the strain for 4 d on MA plate containing NaCl to final concentrations of 0−10% (intervals of 1%). To investigate the effect of varying pH on the growth of strain A28-5, solid media ranging from pH 5.5−10.0 (intervals of pH 0.5) were prepared and the strain was inoculated, fol- lowed by observing the growth by incubating it for 4 days. Enzyme activation was performed using argi- nine dihydrolase, urease, esculin hydrolase (β-glucosi- dase), gelatinase, and β-galactosidase using an API 20NE kit (Biomérieux, France) according to the manu- facturer’s instructions. However, as strain A28-5 was confirmed to be a NaCl-requiring strain, the medium was supplemented with NaCl to a final concentration of 2% and incubated at 37℃ for 2 d, followed by observing the growth.
To investigate the optimal carbon sources that can be used for cultivating strain A28-5, the strain was first inoculated into artificial sea water broth (ASW; 6.1 g Tris base, 12.3 g MgSO4, 0.74 g KCl, 0.13 g (NH4)2HPO4, 17.5 g NaCl, 0.14 g CaCl2, per liter; pH 7.2) containing 0.5% of each carbon source and cultured under shaking conditions at 200 rpm and 37℃ for 4 days. The tested carbon sources were glucose, sucrose, N-acetyl-glucos- amine, carboxylmethyl cellulose (CMC), arabinose, malt- ose, starch, agar, lactose, xylose, inositol, mannitol, sorbitol, xylan, galactose, and glycine.
To investigate the antibiotic sensitivity of strain A28- 5, the bacteria was evenly smeared on MA plate and a paper disc containing 30 μl of each antibiotic were placed at the centre of each plate. The antibiotics tested were chloramphenicol (25 μg/ml), kanamycin (50 μg/ml), ampicillin (50 μg/ml), apramycin (50 μg/ml), thiostrepton (50 μg/ml), nalidixic acid (25 μg/ml), neomycin (30 μg/
ml), and tetracycline (10 μg/ml), and a zone of clearance appearing due to inhibition of growth was observed by incubating at 37℃ for 1 d. C. agarivorans YM01 was used as a comparison strain, on which the same analy- ses were simultaneously performed as described above.
Analysis of biochemical characteristics of strain A28-5 To analyze the biochemical characteristics of strain A28-5, the bacteria grown on MA plate at 37℃ for 2 d was harvested. Analyses of main quinones, fatty acids, and G+C content were conducted at the Korean Culture Center of Microorganisms (KCCM, Korea). The method used for each analysis is as follows: Main quinones were
analyzed by reverse phase HPLC using a C-18 column and the fatty acids present in the bacteria were methyl- esterified following the method previously described by Miller and Berger [15]. The G+C content in the genomic DNA was analyzed by the method of Mesbah et al., which uses reverse phase HPLC [16]. C. agarivorans YM01 was used as a comparison strain, on which similar analyses were performed simultaneously.
Degradation of xylan by strain A28-5
Strain A28-5 was cultured on MA plate containing 0.15% AZCL-xylan at 37℃ for 2 d. In AZCL-xylan-con- taining media, xylan degradation was confirmed by the change in color. Additionally, to confirm that strain A28- 5 secretes the xylan-degrading enzyme extracellularly, it was inoculated into MB and incubated at 37℃ for 2 d, followed by centrifugation at 20,000 ×g for 30 min and the supernatant was collected by discarding the bacte- rial pellet. The supernatant was filtered using a 0.22-μm syringe filter, and 10 μl of the supernatant was spotted on MA plate containing 0.15% AZCL-xylan and was incubated at 37℃ for 12 h, then the degradation of AZCL-xylan was observed.
Degradation of agar by strain A28-5
Strain A28-5 was cultured on MA at 37℃ for 2 d. The plate was then stained with Lugol’s Iodine solution (1%
I2, 2% KI), followed by washing excess Lugol’s Iodine solution with distilled water. Agar degradability around the bacterial colonies was observed due to the property that the Lugol’s Iodine solution stains only polysaccha- rides. The supernatant without bacterial cells was obtained as described in the previous experiment, and 10 μl of this supernatant was spotted on MA plate and incubated at 37℃ for 12 h; agar degradation was observed by staining with the Lugol's Iodine solution.
MA used for this experiment contained AZCL-xylan so that not only agar, but also xylan, degradability could be confirmed simultaneously.
Preparation of xylanase and agarase
Strain A28-5 was inoculated into 200 ml MB contain- ing 0.1% birchwood xylan and incubated at 37℃ for 2 d, followed by centrifugation at 20,000 ×g for 30 min, and the supernatant was collected by discarding the bacte- rial pellet. The supernatant was filtered using a 0.22 μm
syringe filter (Millipore, USA) and was concentrated using an Amicon ultra centrifugal filter (Millipore, USA), and 2 ml of xylanase was obtained. Agarase was prepared similar to xylanase, using an agar-containing culture medium.
Thin layer chromatography (TLC) analysis
To analyze the xylan hydrolysates produced by the action of xylanases, secreted by the bacterial strain A28- 5, 100 μl of the prepared xylanase was added to 1×
phosphate-buffered saline (PBS) solution containing 0.4% birchwood xylan and allowed to react at 37℃ for 24 h. At 12 h interval during the enzyme reaction, 20 μl of reaction solution was sampled and applied to a Silica gel 60 plate (Merck, USA) using xylobiose and xylo- tetraose (Megazyme, Ireland) as standards. A solution of n-butanol: acetic acid:distilled water (2:1:2) was used as a developing solvent, and a solution of ethanol:sulfuric acid (9:1) was used as a coloring reagent. After develop- ment, the coloring reagents were evenly sprayed and visualized by heating at 120℃.
To analyze hydrolysis products of agar by agarase secreted by strain A28-5, 100 μl of the agarase crude enzyme solution was added to 1×PBS buffer containing 0.4% agarose and reacted at 45℃ for 24 h. At this time, the temperature was set as the minimum temperature at which the agarose did not solidify after melting. The analytical method was the same as that of xylanase hydrolysis products. However, purely purified neoagaro- biose, neoagarotetraose, and neoagarohexaose were used as standard to compare the size of hydrolysates [17].
Results
Identification of strain A28-5
A marine microbial strain showing highest xylan and agar degradabilities was finally selected and designated as strain A28-5 (Fig. 1A and B). The homology search based on the 16S rRNA gene sequence showed the fol- lowing results for strain A28-5−98.94% homology with C. agarivorans YM01 (GQ262000), 95.78% with G.
joonbiniege G7 (AMRX01000001), 95% with Catenovulum maritimus Q1 (NR_146038), 91.02% with Algicola sag- amiensis B-10-31 (AB063324), and 90.95% with Echini- monas agarilytica KMM6351 (JX072970).
NJ phylogenetic tree was constructed based on the
16S rRNA gene sequence homology search results. The phylogenetic analysis showed that strain A28-5 is most closely related to C. agarivorans YM01 (Fig. 1D). Phylo- genetic trees constructed by the Maximum-Parsimony (MP) and the Maximum-Likehood (ML) methods showed a similar relationship as that of the NJ phylogenetic tree (data not shown). Based on the results of 16S rRNA gene sequence homology and phylogenetic analysis, it is possi-
ble that strain A28-5 is of the same genus and species as C. agarivorans YM01 or has evolved into a different spe- cies. The DNA hybridization analysis between strain A28-5 and the strains with high 16S rRNA gene sequence and phylogenetic associations showed the hybridization value of 61.83% with C. agarivorans YM01 (Fig. 1C). Generally, it is considered to be a new subspe- cies when the DNA-DNA hybridization value is in the Fig. 1. Isolation of strain A28-5. (A) Xylanase activity. Strain A28-5 was cultured on MA containing AZCL-xylan at 37℃ for 2 d.
(B) Agarase activity. Strain A28-5 was cultured on MA at 37℃ for 2 d. The plate was stained with Lugol’s iodine solution. (C) DNA- DNA hybridization. The values obtained from self hybridization of strain A28-5 were set at 100%. (1), strain A28-5; (2), Catenovulum agarivorans YM01; (3), Micrococcus luteus KCCM11326 as the negative control. (D) Neighbor joining phylogenetic analysis. Phyloge- netic tree was constructed based on the 16S rRNA gene sequences of strain A28-5 and its genetically related strains. Distances was determined according to the Kimura-two model and bootstrap values (>50%) based on 1,000 replicates are listed as percentages at nodes. Nucleotide sequence accession numbers are given in parentheses. Scale bar, 0.01 substitutions per 100 nucleotides.
range of 70−90%, the same species when the value is 90% or higher, or a different species when the value 70%
or lower [18]. According to these criteria, the DNA hybridization value of 61.83% classifies strain A28-5 as a different species in the same genus as that of C. agariv- orans YM01. G. joonbiniege G7, the strain with the sec- ond highest 16S rRNA gene sequence homology after C.
agarivorans YM01, showed a DNA hybridization value of 13.56% (data not shown). As this value is lower than the cut-off value (i.e., 20%), it may indicate a different genus, it is thought that these two bacterial strains may have evolved into different genera. Thus, in the present study, strain A28-5 can be classified as a novel species of the genus Catenovulum from the results of 16S rRNA gene sequence analysis, phylogenetic tree analysis, and DNA-DNA hybridization analysis.
Morphological and physiological characteristics of strain A28-5
Strain A28-5 was confirmed to be gram-negative by Gram staining. The motility of strain A28-5 was con- firmed by observing flagella using Transmission Election Microscope (TEM) after negative staining (data not shown).
Strain A28-5 grew only in the presence of NaCl (1−4%)
and not in the medium without NaCl. This indicates that strain A28-5 requires NaCl for growth. pH 7.0−9.0 was observed to be optimal for the growth of the strain, whereas the growth was not observed below or above these pH conditions.
Strain A28-5 grew in the medium containing glucose, sucrose, N-acetyl-glucosamine, arabinose, maltose, starch, agar, lactose, xylose, mannitol, xylan, and galactose.
However, the growth was not observed in the medium containing CMC, inositol, sorbitol, or glycine. This char- acteristic was similar to that of the type strain, C. aga- rivorans YM01. In case of antibiotic sensitivity, C.
agarivorans YM01 showed resistance towards ampicillin only, whereas Catenovulum sp. A28-5 showed resistance towards two antibiotics—ampicillin and thiostreptone whereas it was sensitive towards other antibiotics including chloramphenicol, kanamycin, apramycin, nali- dixic acid, neomycin, and tetracycline, and this sensitiv- ity was similar to that of C. agarivorans YM01. For all the above experiments, the growth of the comparison strain, C. agarivorans YM01, was observed in similar conditions. The characteristics of strain A28-5, C. aga- rivorans YM01 and C. maritimus Q1 are compared and shown in Table 1. Strain A28-5 can be distinguished from the two type strains by several characteristics such
Table 1.Distinguishable characteristics of strain A28-5, Catenovulum agarivorans YM01, Catenovulum maritimus Q1. Strains:
1, A28-5 (this study); 2, Catenovulum agarivorans YM01 (this study); 3, Catenovulum maritimus Q1 (data from Li et al. [7]). The G+C content (mol%), isoprenoid quinine and motility results of Catenovulum agarivorans YM01 are from Yan et al. [6].
Characteristics 1 2 3
Source Sea water, Jeju Island, Korea Sea water, Qingdao, China Porphyra yezoensis Ueda, Weihai, China
G+C content (mol%) 43.96 44.8 37.9
Motility Polar flagella Peritrichous flagella Peritrichous flagella
Colony color Pale yellow Pale yellow Pale white
Growth at:
pH 7.0−9.0 7.5−9.5 6.0−8.5
NaCl concentration (%) 1−4 1−3 0.5−5.5
Utilization of:
Sorbitol − + −
Enzyme activity
Arginine dihydrolase − + −
Nitrate to nitrite − + −
Resistant to:
Ampicillin (50 μg/ml) + - ND
Isoprenoid quinone MK-7 MK-7 Q-8
Symbols : +, positive reaction; −, negative reaction; MK-7, menaquinone-7; Q-8, ubiquinone-8; ND, not determined.
as the cells morphology, G+C content, enzyme activity and growth condition.
Biochemical characteristics of strain A28-5
The G+C content in the genomic DNA of strain A28-5 was analyzed to be 43.96% which is similar to that of the comparison strain, C. agarivorans YM01 (44.8%) (Table 1). Additionally, menaquinone-7 (MK-7) was observed as the predominant quinone in the strain A28-5, which is the similar characteristic of the type strain C. agariv- orans YM01, and it is specifically present in species of the class δ-proteobacteria (Table 1).
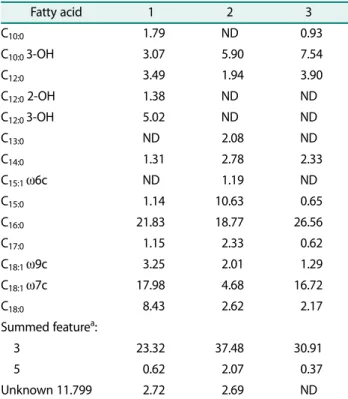
The fatty acids present in the cell wall of strain A28-5 were analyzed and were composed of straight-chain fatty acids (50.42%), unsaturated fatty acid (21.95%)], branched fatty acid (0.52%), Summed feature (23.94%), and unknown (3.16%). The cellular fatty acids of C. aga- rivorans YM01, the phylogenetically associated compari- son strain, were simultaneously analyzed using the
same method. In particular, three types of fatty acids (C10:0, C12:0 2-OH, and C12:0 3-OH) were found only in strain A28-5 but not in C. agarivorans YM01, while two types of fatty acids (C13:0 andC15:1 ω6c) were found only in C. agarivorans YM01 but not in strain A28-5. Five types of fatty acids (C15:0, C18:0,C18:1 ω7c, summed fea- ture 3, and summed feature 5) clearly showed differ- ences between the two strains. Such differences in fatty acid content and type between the two strains indicate that the two strains have evolved into different species.
Although strain A28-5 and C. maritimus Q1 share the same major cellular fatty acids (C16:0, C18:1 ω7c, and summed feature 3), other quantitative differences in the fatty acids indicated that this isolate may represent a different species from C. maritmus Q1. The cellular fatty acid composition of strain A28-5, C. agarivorans YM01, and C. maritimus Q1 is shown in Table 2.
Based on the results of molecular, biological, morpho- logical, physiological, and biochemical analyses, strain A28-5 is thought to be a novel species of the genus Catenovulum. Therefore, in the present study, strain A28-5 was named as Catenovulum jejuensis A28-5.
Degradation of xylan by Catenovulum jejuensis A28-5 As shown in Fig. 1A, the insoluble AZCL-xylan parti- cles were solubilized to form a blue coloration around the bacterial colonies. This blue coloration indicates that strain A28-5 secretes xylan-degrading enzymes extracel- lularly. Agar degradation ability was observed in liquid culture medium containing agar as a carbon source, but xylan degradation was not observed, which means that xylan degrading enzyme produced in C. jejuensis A28-5 is induced only in the presence of xylan. Therefore, in order to produce xylan-degrading enzyme using the strain, it is considered that xylan should be prepared and contained in the medium.
TLC analysis of the reaction solution containing birch- wood xylan and xylanase obtained from the broth in which C. jejuensis A28-5 was grown, showed that xylanase secreted by C. jejuensis A28-5 degrades xylan to produce tetramers and hexamers as major degrada- tion products (Fig. 2A). This TLC pattern is consistent with the hydrolysis characteristics of the common endo- type xylanase. Therefore, it is possible to produce useful oligosaccharides such as tetramer and hexamer from xylan using the enzymes of the strain.
Table 2. Cellular fatty acid composition of strain A28-5, Catenovulum agarivorans YM01 and Catenovulm maritimus Q1. Strains: 1, A28-5 (this study); 2, Catenovulum agarivorans YM01 (this study); 3, Catenovulum maritimus Q1 (data from Li et al. [7]).
Fatty acid 1 2 3
C10:0 1.79 ND 0.93
C10:0 3-OH 3.07 5.90 7.54
C12:0 3.49 1.94 3.90
C12:0 2-OH 1.38 ND ND
C12:0 3-OH 5.02 ND ND
C13:0 ND 2.08 ND
C14:0 1.31 2.78 2.33
C15:1 ω6c ND 1.19 ND
C15:0 1.14 10.63 0.65
C16:0 21.83 18.77 26.56
C17:0 1.15 2.33 0.62
C18:1 ω9c 3.25 2.01 1.29
C18:1 ω7c 17.98 4.68 16.72
C18:0 8.43 2.62 2.17
Summed featurea:
3 23.32 37.48 30.91
5 0.62 2.07 0.37
Unknown 11.799 2.72 2.69 ND
ND, not detected; Sum 3, C16:1 ω7c/iso-C15:0 2-OH; 5, C18:2 ω6c, 9c/Anteiso-C18:0.
Degradation of agar by Catenovulum jejuensis A28-5 The strain was incubated at 37℃ for 2 days and stained with Lugol’s Iodine solution to confirm agar deg- radation around the cells (Fig. 1B). Agar degradation was observed in the supernatant from which the cells were removed by incubation in MB liquid medium con- taining agar. It was found that C. jejuensis A28-5 secreted an agarase enzyme outside the cell. In the medium containing xylan as a carbon source, weak agar degradability was observed. This means that the agar- degrading enzyme is produced by C. jejuensis A28-5 at low levels even in the absence of agar. However, in order to induce the production of more agarase from the strain, agar should be used as a carbon source. As a result of TLC analysis of enzyme reaction with agarose using agarase prepared from C. jejuensis A28-5 liquid culture, it was possible to know that the agarase secreted by C. jejuensis A28-5 degrades agarose to produce (neo)agarotetraose (major product) and (neo)agarohexaose (minor product) (Fig. 2B). This TLC pattern is consistent with hydrolysis characteristics of common endo-type agarases. Therefore, it is possible to produce tetramer and hexamer (neo)agaro-oligosaccharides from agar using the enzyme of the strain.
Discussion
The first important step for a successful biotechnologi- cal application is to isolate a microorganism that pro- duces industrially useful xylanase and agarase and to reveal the characteristics of the enzymes produced by the strain. In particular, oligosaccharide production through polymer degradation by xylanase and agarase has been recently receiving attention, and specifically activated enzymes are required for the production of tar- geted oligosaccharides. In the present study, a marine microorganism that secretes xylanase and agarase extracellularly was isolated and identified as a novel species of the genus Catenovulum by morphological, physiochemical, and genetic analyses, and the degrada- tion mechanisms of the enzymes produced by this strain for polymers of xylan and agar were revealed. Through xylan hydrolysis using xylanase prepared from the liquid medium in which C. jejuensis A28-5 was grown, the tetramers and hexamers of XOSs were obtained, and through agarose hydrolysis using agarase, tetramers and hexamers of (neo)agarooligosaccharides were obtained.
Based on enzyme activities in the liquid medium, it is expected that the genome of strain A28-5 contains many genes encoding xylanase and agarase. In fact, in C. aga- rivorans YM01, the type strain belonging to the same genus as strain A28-5, after being identified as a new agar-degrading Catenovulum genus in 2011, a genome sequence of 4.87 Mb was determined by Shi et al., and a total of 15 genes encoding agarase were identified in the genome [19]. Among these, it was shown that 2 genes encode α-agarase and 13 genes encode β-agarase. Pre- ceding this, in Alteromonas sp. strain S89, which was identified as an agarase-producing marine microbial strain and whose genome sequence was determined, 6 agarase coding genes were discovered [20]. Recently, 2 β- agarases were discovered in Agarivorans gilvus WH0801 [21]. As compared to these microorganisms, C. agariv- orans YM01 encodes relatively many agarases. When cultured on an agar plate, the agar degradation zone around the colonies of C. jejuensis A28-5 was observed to be similar to that of C. agarivorans YM01. This indicates that the strain can be expected to encode various aga- rases similar to C. agarivorans YM01. Currently, we are sequencing the entire genome of C. jejuensis A28-5 using Fig. 2.Thin layer chromatography analysis. (A) Degradation
of birchwood xylan. Crude enzyme of strain A28-5 was reacted with birchwood xylan at 40℃. S1 : xylobiose (X2); S2 : xylo- tetraose (X4); 1 and 2 : reactant of crude enzyme and birch- wood xylan. Arrows indicate tetramer and hexamer, respectively.
(B) Degradation of agarose. Crude enzyme of strain A28-5 was reacted with agarose at 40℃. S : neoagarobiose (NG2), neoag- arotetraose (NG4), neoagarohexaose (NG6); 1 and 2 : reactant of crude enzyme and agarose. Arrows indicate (neo)agaro- tetraose and (neo)agarohexaose, respectively.
the PacBio sequencing technology. Once the genome is sequenced, we will be able to compare C. jejuensis A28-5 with type strains at the genetic level. In addition, by securing the xylanase and agarase genes, it is expected that the mass production of these enzymes will be possi- ble via genetic engineering techniques.
In the present study, a novel species of the genus Catenovulum, which secretes xylanase and agarase, was identified from the coastal seawater of Jeju Island, Korea, and its characteristics were revealed. Addition- ally, the hydrolytic properties of the enzymes produced by the strain were analyzed by TLC. The present study provides important information on the enzymes that can produce various oligosaccharides by decomposing plant cell wall components.
GenBanck accession number and strain deposit
The Genbank accession number for the 16S rRNA gene sequence of strain A28-5 is KC433404. The strain A28-5 was deposited in KACC (KACC92062).
Acknowledgments
This work was supported by a grant from the National Institute of Biological Resources (NIBR), funded by the Ministry of Environment (MOE) of the Republic of Korea (NIBR201629101).
References
1. Chi WJ, Chang YK, Hong SK. 2012. Agar degradation by microorganisms and agar-degrading enzymes. Appl. Microbiol.
Biotechnol. 94: 917-930.
2. Kobayashi R, Takisada M, Suzuki T, Kirimura K, Usami S. 1997.
Neoagarobiose as a novel moisturizer with whitening effect.
Biosci. Biotechnol. Biochem. 61: 162-163.
3. Christakopoulos P, Katapodis P, Kalogeris E, Kekos D, Macris BJ, Stamatis H, et al. 2003. Antimicrobial activity of acidic xylo-oligo- saccharides produced by family 10 and 11 endoxylanases. Int. J.
Biol. Macromol. 31: 171-175.
4. Kallel F, Driss D, Chaabouni SE, Ghorbel R. 2015. Biological activi- ties of xylooligosaccharides generated from garlic straw xylan by purified xylanase from Bacillus mojavensis UEB-FK. Appl. Biochem.
Biotechnol. 175: 950-964.
5. Mario LP, Francisco R-V. 2014. The family Alteromonadaceae. The Prokaryotes. pp. 69-92. Springer-Verlag Berlin Heidelberg, New York.
6. Yan S, Yu M, Wang Y, Shen C, Zhang XH. 2011. Catenovulum aga- rivorans gen. nov., sp. nov., a peritrichously flagellated, chain- forming, agar-hydrolysing gammaproteobacterium from seawa-
ter. Int. J. Syst. Evol. Microbiol. 61: 2866-2873.
7. Li DQ, Zhou YX, Liu T, Chen GJ, Du ZJ. 2015. Catenovulum mariti- mus sp. nov., a novel agarolytic gammaproteobacterium isolated from the marine alga Porphyra yezoensis Ueda (AST58-103), and emended description of the genus Catenovulum. Antonie Van Leeuwenhoek. 108: 427-434.
8. Biely P. 1985. Microbial xylanolytic systems. Trends Biotechnol. 11:
286-290.
9. Chi WJ, Park JS, Kwak MJ, Kim JF, Chang YK, Hong SK. 2013. Isola- tion and characterization of a novel agar-degrading marine bac- terium, Gayadomonas joobiniege gen, nov, sp. nov., from the Southern Sea, Korea. J. Microbiol. Biotechnol. 23: 1509-1518.
10. Kim JH, Choi BH, Jo M, Kim SC, Lee PC. 2014. Flavobacterium faecale sp. nov., an agarase-producing species isolated from stools of Antarctic penguins. Int. J. Syst. Evol. Microbiol. 64: 2884- 2890.
11. Amel BD, Nawel B, Khelifa B, Mohammed G, Manon J, Salima KG, et al. 2016. Characterization of a purified thermostable xylanase from Caldicoprobacter algeriensis sp. nov. strain TH7C1(T). Carbo- hydr. Res. 419: 60-68.
12. Morais CG, Lara CA, Oliveira ES, Peter G, Dlauchy D, Rosa CA.
2015. Spencermartinsiella silvicola sp. nov., a yeast species iso- lated from rotting wood. Int. J. Syst. Evol. Microbiol. doi:10.1099/
ijsem.0.000764.
13. Lane DJ. 1991. 16S/23S rRNA sequencing. pp. 115-175. In Stacke- brandt E, Goodfellow M (eds.), Nucleic acid techniques in bacte- rial systematics. Chichester, United Kingdom: John Wiley and Sons.
14. Saitou N, Nei M. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4:
406-425.
15. Miller L, Berger T. 1985. Bacterial identification by gas chroma- tography of whole cell fatty acid. Hewlett-Packard Application note. pp. 228-241.
16. Mesbah M, Premachandran U, Whitman WB. 1989. Precise mea- surement of the G+C content of deoxyribonucleic acid by high- performance liquid chromatography. Int. J. Syst. Bacteriol. 39:
159-167.
17. Temuujin U, Chi WJ, Lee SY, Chang YK, Hong SK. 2011. Overex- pression and biochemical characterization of DagA from Strepto- myces coelicolor A3(2): an endo-type β-agarase producing neoagarotetraose and neoagarohexaose. Appl. Microbiol. Bio- technol. 92: 749-759.
18. Van Trappen S, Tan TL, Yang J, Mergaert J, Swings J. 2004. Alter- monas stellipolaris sp. nov., a novel, budding, prosthecate bacte- rium from Antarctic seas, and emended description of the genus Alteromonas. Int. J. Syst. Evol. Microbiol. 54: 1157-1163.
19. Shi X, Yu M, Yan S, Dong S, Zhang XH. 2012. Genome sequence of the thermostable-agarase-producing marine bacterium Caten- ovulum agarivorans YM01(T), which reveals the presence of a series of agarase-encoding genes. J. Bacteriol. 194: 5484.
20. Oh C, De Zoysa M, Kwon YK, Heo SJ, Affan A, Jung WK, et al. 2011.
Complete genome sequence of the agarase-producing marine
bacterium strain s89, representing a novel species of the genus Alteromonas. J. Bacteriol. 193: 5538.
21. Zhang P, Rui J, Du Z, Xue C, Li X, Mao X. 2016. Complete genome
sequence of Agarivorans giluvs WH0801(T), and agarase-produc- ing bacterium isolated from seaweed. J. Biotechnol. 219: 22-23.
국문초록
제주 연안해수로부터 한천 분해 효소 및 자일란 분해 효소를 생산하는 Catenovulum jejuensis A28-5의 동정 및 특성 규명 김다솜, 정가람, 배창환, 여주홍, 지원재*
국립생물자원관생물자원활용부유용자원분석과
Strain A28-5는대한민국제주도연안의해수샘플로부터고체배지내 xylan과 agar를분해하는균주로분리되었다. Strain A28- 5는그람음성균으로한개의 polar flagella로운동성을갖는 Na+이온요구성균주로분석되었다. 또한 ampilcillin과 thiostreptone 등의항생제에내성을보였다. Genome 내 G+C content는 43.96%이고, Menaquinone-7 (MK-7)을 predominant quinone으로 함유하고있었다. Strain A28-5의세포벽을구성하는주요지방산은 C16:1 ω7c/iso-C15:0 2-OH (23.32%), C16:0(21.83%), C18:1ω7c (17.98%)였다. strain A28-5의 16S rRNA gene sequence는 Catenovulum agarivorans YM01와가장높은상동성(98.94%)을보 였으며, Neighbor-Joining phylogenetic tree 제작을통해서 Catenovulum agarivorans YM01와가장높은근연관계를보이는 것을증명하였다. Catenovulum agarivorans YM01과의 DNA-DNA hybridization 분석을통하여 A28-5을 Catenovulum 속의 신종으로분류하여Catenovulum jejuensis A28-5로명명하였다. 이균주의액체배양으로부터준비된두종류의조효소를이용한 xylan 또는 agarose 와의효소반응액을 Thin layer chromatography로분석하여각각 tetramer와 hexamer의 xylooligosaccharides 와 (neo)agarooligosacchardes가생산되는것을확인하였다.