309 책임저자:김남득, 609-735, 부산시 금정구 장전동 산 30
부산대학교 약학대학 약학부 Tel: 051-510-2801, Fax: 051-513-6754 E-mail: [email protected]
접수일:2009년 10월 13일, 게재승인일:2009년 10월 23일
Correspondence to:Nam Deuk Kim
Department of Pharmacy, Pusan National University, San 30, Jangjeong- dong, Kcumjeong-gu, Busan 609-735, Korea
Tel: +82-51-510-2801, Fax: +82-51-513-6754 E-mail: [email protected]
Urushiol Induces Growth Inhibition and Apoptosis in MCF-7 Human Breast Cancer Cells
Seaho Kim1, So-Hyun Jeong1, Mohammad Akbar Hossain1, Min Young Kim1, Dong Hwan Kim1, Jin-Ah Kim1, Jeong-Hyun Yoon1, Chun Soo Na2 and Nam Deuk Kim1
1Department of Pharmacy (BK21 Program), College of Pharmacy, Molecular Inflammation Research Center for Aging Intervention (MRCA), Pusan National University, Busan 609-735, 2Lifetree Biotech Co., Ltd., Suwon 441-853, Korea
In this study, we investigated the effects of urushiol which was isolated from the sap of Korean lacquer tree (Rhus vernicifera Stokes), on the proliferation of MCF-7 human breast cancer cells. Forty-eight hour treatment of urushiol inhibited the growth of breast cancer cells and induced cell DNA fragmentation.
This urushiol-induced apoptosis in the MCF-7 cells was closely linked with the down-regulation of Bcl-2 protein expression and the cleavage of poly (ADP-ribose) polymerase. Urushiol also caused a marked increase in the level of p21WAF1/CIP1protein in a p53-dependent manner. Based on our data, urushiol can be considered as a good candidate for an effective chemotherapeutic agent inducing apoptosis of cancer cells, although further study will be needed to confirm this. (Cancer Prev Res 14, 309-314, 2009) Key Words: Urushiol, Human breast cancer cells, Apoptosis
INTRODUCTION
Urushiol is extracted from cortex of Rhus verniciflua Stokes and a mixture of various analogues.1∼5) This mixture is notorious for inducing hypersensitivity reaction as a main component of Western poison ivy (Toxicodendron radicans).6∼8) The potential therapeutic usefulness of urushiol has been overlooked although there are many immunological studies of urushiol. The cytotoxic effects of urushiol compounds have been studied in 29 different kinds of human cancer cell lines, including MCF-7 human breast cancer cells, and the value of 50% growth inhibition (GI50) has been reported to be ap- proximately 4.0μg/ml, showing cell line-specific cytotoxicity.9) However, only cytotoxic sulforhodamine B assay was conducted and, therefore, the underlying molecular mechanism of the cytotoxicity of urushiol was not fully elucidated.
Breast cancer is by far the most common cancer amongst women worldwide including South Korea.10) It is currently
treated by surgery, radiotherapy, and immunotherapy, and is frequently managed by adjuvant chemotherapies or hormonal therapies. Breast cancer, however, is highly resistant to chemotherapy, and there are still no effective means of cure for patients with advanced stages of the disease, especially in cases of hormone-independent cancer. Therefore, effective che- motherapeutic agents for breast cancer would have a significant impact on morbidity and mortality of breast cancer.
It is widely accepted that tumor cells treated with anticancer agents die from apoptosis11) and emerging evidences have demonstrated this. The phenomenon of apoptosis plays a necessary role as a protective mechanism against carcinogenesis by eliminating damaged cells or abnormally proliferating cells stimulated by various chemical agents.12∼14) The molecular mechanism of regulation of apoptosis is a complex process and involves a number of death- or survival-related cellular genes, such as oncogenes of c-Myc, p53, as well as p21WAF1/CIP1, Bcl-2, and Bax which are downstream target genes of p53.15∼17) In this study, we examined whether urushiol inhibits cell growth
Fig. 1. Growth inhibition of MCF-7 cells after treatment with urushiol. Cells were plated at 2×104 cells per 60-mm plate, and incubated for 24 h. The cells were treated with variable concentrations of urushiol for 48 h and the viability was measured by the metabolic-dye-based MTT assay. Viability of control cells was set at 100% and viability relative to the control is presented. Results are expressed as percentage of the vehicle treated control±SD of three separate experiments. The significance was determined by Student’s t-test (*p<0.05 or
**<0.01 vs. untreated control).
and induces apoptosis in MCF-7 breast cancer cells.
MATERIALS AND METHODS 1. Cell culture and urushiol solution
MCF-7, a human breast cancer cell line, was obtained from American Type Culture Collection (Manassa, VA, USA). The MCF-7 cells were cultured on DMEM supplemented with 10%
FBS, 50 mg/ml gentamicin (Sigma Chemical Co., St. Louis, MO, USA), and 135 mg/ml glutamine (Sigma) at 37°C incubator in a humidified atmosphere containing 5% CO2. The medium was changed three times per week. The number of cells was counted with a hematocytometer. Urushiol complex was obtained in a solution form from Lifetree Biotech Co., Ltd.
(Suwon, Korea). The aqueous solution was filtered through Whatman No. 2 filter paper, and then concentrated in vacuum to dryness. The yield of urushiol was about 3.5%. The powder was weighted and adjusted to a final concentration with DMSO (Sigma, St. Louis, MO, USA) and stored at −20°C until use.
The stock solution of urushiol was diluted with medium to the desired concentration prior to use. The maximal concentration of DMSO did not exceed 0.1% (v/v) in the treatment range (2∼10μg/ml), where there was no influence on the cell growth (data not shown).
2. Assessment of cell proliferation by MTT assay
The proliferation of MCF-7 cells was assessed by using the MTT assay which is based on the conversion of MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide]
to MTT-formazan by mitochondrial enzyme.18) In brief, these cells were cultured with 1 ml of medium per well in 24-multiwell plates at 37°C for 24 h under 5% CO2 in air.
The cells were then treated with desired concentrations of urushiol. Then, the MTT solution (5 mg/ml PBS) was added to the well with a concentration of 0.25 mg/ml. Following the incubation for 4 h, the mixture of medium and the MTT solution was carefully discarded, after which 1 ml EtOH- DMSO (1:1) was added to the well in order to extract the crystallized dye. The amount of blue dye formed was deter- mined by measuring the absorbance at 540 nm. Measurements were performed in triplicates.
3. Agarose gel analysis of DNA fragmentation Cells were harvested, rinsed twice in cold PBS, and resus-
pended in lysis buffer [5 mM Tris-HCl (pH 7.5), 5 mM ethylenediaminetetra acetic acid (EDTA), and 0.5% Triton X-100] at 4°C for 30 min. After centrifugating them at 27,000×g for 15 min, the supernatant was treated with RNase, followed by proteinase K digestion, phenol/chloroform/isoamyl alcohol (25:24:1) extraction and isopropanol precipitation.
DNA separated through a 1.5% agarose gel was stained with ethidium bromide (EtBr, Sigma) and visualized by ultraviolet light source.
4. Gel electrophoresis and western blotting
The cells were harvested, lysed, and protein concentrations were quantified using the Bio-Rad protein assay (Bio-Rad Lab., Hercules, CA, USA) by the procedure described by the ma- nufacturer. For the Western blot analysis, an equal amount of protein was subjected to electrophoresis on SDS-polyacrylamide gels and transferred to nitrocellulose membranes (Schleicher &
Schuell, Keene, NH, USA) by electroblotting. Blots were pro- bed with the desired antibodies for 1 h, incubated with diluted enzyme-linked secondary antibodies and then visualized by the enhanced chemiluminescence (ECL) according to the recommen- ded procedure (Amersham Pharmacia Biotech Inc., Piscataway, NJ, USA). The primary antibodies were purchased from Santa
Fig. 2. Representative microphotographs of urushiol treated MCF-7 cells. Cells were plated at 2×104 cells per 60-mm plate, incubated for 24 h and treated with urushiol for 48 h. (A) Control, (B) 2μg/ml, (C) 4μg/ml, (D) 6μg/ml, (E) 8μg/ml, (F) 10μg/ml.
The cells were photographed by phase contrast microscopy (original magnification 400×).
Cruz Biotechnology Inc. (Santa Cruz, CA, USA) and Calbiochem (Cambridge, MA, USA). Peroxidase-labeled secondary antibodies were purchased from Amersham Pharmaci Biotech.
5. Statistics
Data were expressed as the mean±SD of three separate experiments and analyzed by Student’s t-test. The means were
Fig. 3. Apoptosis inducing effects of urushiol on MCF-7 cells. (A) Cells were incubated with urushiol for 48 h. Internucleosomal DNA fragmentation was analyzed by 1% agarose gel electrophoresis in the presence of EtBr. (B) Induction of PARP cleavage in MCF-7 cells was detected by anti-parp antibody. (C) The cells were treated with variable concentration of urushiol for 48 h and total cell lysates were prepared and immunoblotted. Western blots were detected with antibodies against BCL-2, Bax, and β-actin, and ECL detection. A representative blot is shown from three independent experiments. C, Control.
considered significantly different at either *p<0.05 or **p
<0.01.
RESULTS
1. Cytotoxic and growth inhibitory effects of urushiol
The effect of urushiol on the viability of MCF-7 cells was assessed via MTT dye assay at various concentrations. As shown in Fig. 1, the cell viability was decreased in a concentration- dependent manner when the cells were exposed to the concentrations of urushiol greater than 4μg/ml for 48 h. The dose required for half-maximal inhibition (IC50) of MCF-7 cell growth was approximately 6μg/ml and the treatment with higher urushiol concentrations (8∼10 μg/ml) for 48 h showed more than 75% reduction of the cell viability. Microphoto- graphs of urushiol-treated MCF-7 cells also showed distinct morphological changes and cell viability (Fig. 2).
2. Induction of apoptosis in MCF-7 cells
We assessed the effects of urushiol on apoptosis induction in the MCF-7 cells using DNA fragmentation assay. As shown in Fig. 3A, agarose gel electrophoresis of DNA of urushiol treated-cells showed ladders of DNA fragmentation which is indicative of induced cell apoptosis. Polypeptide degradation, including poly (ADP-ribose) polymerase (PARP), was examined to see the possible involvement of apoptosis-associated protease during the growth inhibition of the breast cancer cells. PAPR cleavage was evident by the appearance of the p85 PARP cleavage fragment (Fig. 3B) and clearly observed in the 8μg/
ml of urushiol-treated cells which were undergoing apoptosis.
In addition, we studied the changes in the levels of Bax and Bcl-2 expression since apoptosis is known to be regulated by the alteration in the ratio of Bcl-2/Bax protein expression. The Western blotting results showed that the significant down- regulation of Bcl-2 expression and up-regulation of Bax ex-
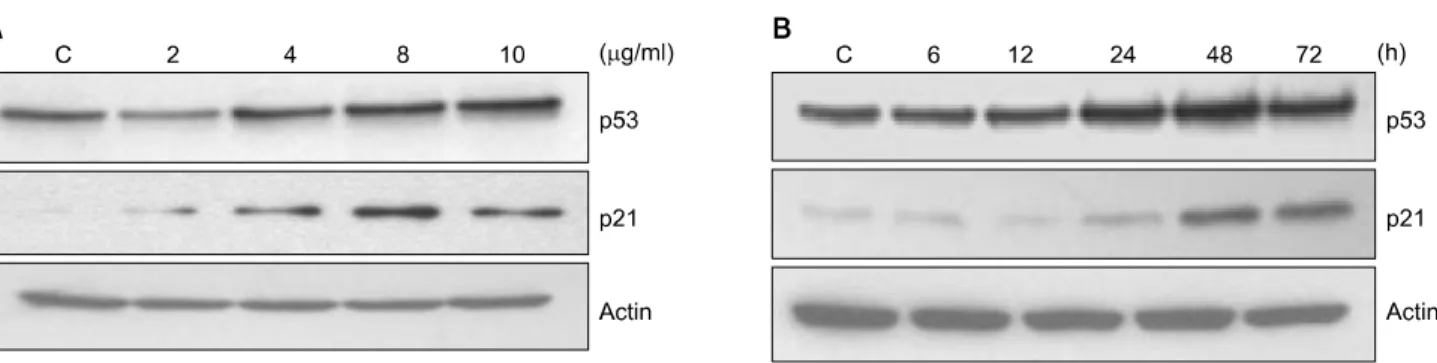
Fig. 4. Dose- and time-dependent effects of urushiol on the protein levels of p53 and p21 in MCF-7 cells. (A) To study dose-dependent effects of urushiol, the cells were treated with variable concentration of urushiol for 48 h. (B) To study time-dependent effects of urushiol, the cells were treated with 8 μg/ml urushiol for each time point. Total cell lysates were prepared and immunoblotted. Western blots were detected with antibodies against p53, p21, and β-actin, and ECL detection. A representative blot is shown from three independent experiments. C, control.
pression in the urushiol-treated MCF-7 cells (Fig. 3C).
3. Up-regulation of p53 and p21WAF1/CIP1 protein in MCF-7 cells treated with urushiol
The tumor suppressor gene, p53, has been shown to regulate a DNA damage-triggered G1 checkpoint by up-regulating the expression of p21WAF1/C1P1, a cyclin-dependent kinase (Cdk) in- hibitor19,20) Our results showed that urushiol treatment resulted in a dose-dependent increase in the p21WAF1/CIP1 protein level via p53-dependent manner (Fig. 4A). Besides, the treatment with 8μg/ml of urushiol for 48 h or 72 h resulted in time-dependent up-regulation of p21WAF1/CIP1 protein level via p53-dependent manner (Fig. 4B).
DISCUSSION AND CONCLUSION
Urushiol seems to have potential cytotoxic effects on the proliferation of human breast cancer cells via apoptosis. Uru- shiol reduced the viability of MCF-7 cells through the in- duction of apoptosis in a dose-dependent manner. By observing internucleosomal DNA fragmentation, we could confirm that urushiol induced apoptosis in MCF-7 cells.
Apoptosis is important regulatory mechanisms that removes unwanted cells during the development and thereby maintain the cell homeostasis. The molecular mechanisms by which anticancer agents induce apoptosis involve the activation of proapoptotic signaling or the inhibition of survival signaling.
Therefore, the balance between the survival and death signal transduction pathways is important in controlling apopto- sis.21,22)
It has been shown that the Bcl-2 family plays an important regulatory role in apoptosis, either as an inhibitor or as an activator. In particular, Bcl-2 has been reported to directly inhibit members of the caspase family, including caspases-3 and -9.23) Thus, it has been suggested that the ratio between the level of pro-apoptotic Bax protein and that of the anti- apoptotic factor Bcl-2 protein determines how a cell responds to an apoptotic signal.21) In this study, urushiol altered the levels of Bax and Bcl-2 expression in MCF-7 cells, resulting in an increase in the ratio of Bax/Bcl-2 (Fig. 3C).
p53 plays a key role in mediating the cell response to various stressors, mainly by inducing or repressing a number of genes involved in cell cycle arrest, senescence, apoptosis, DNA repair, and angiogenesis. p53 mutations are observed in a significant minority of breast tumors. In the remaining cases, the alte- ration of interactive components or target genes could con- tribute, to some extent, to the reduced ability of p53 efficiently managing stressful events. While the prognostic and predictive value of p53 is still in debate, there is an increasing interest for p53-based therapies.24) Furthermore, our study found that urushiol increased the expression of p21WAF1/CIP1 in a p53- dependent manner.
As a conclusion, urushiol demonstrated cytotoxicity and induced apoptosis in MCF-7 cells. Our data also suggest that the anti-proliferative effects of urushiol on MCF-7 cells appear to be mediated, at least partially, through the p53-dependent regulation of p21WAF1/CIP1expression, and by a reduction of Bcl-2. Therefore, the development of modified synthetic uru- shiol derivatives, provided by increased its antitumor activity, would represent a potential new class of chemotherapeutic
agents. Although our studies have commenced to elucidate the antitumor activities of urushiol, further study of urushiol would be needed to confirm its potential as a candidate for chemo- therapeutic agent.
ACKNOWLEDGEMENT
This work was supported for two years by Pusan National University Research Grant.
REFERENCES
1) Symes WF, Dawson CH. Poison ivy "urushiol". J Am Chem Soc 76, 2959-2963, 1954.
2) Markiewitz KH, Dawson CR. On the isolation of the aller- genically active components of the toxic principle of poison ivy. J Org Chem 30, 1610-1613, 1965.
3) Corbett MD, Billets S. Characterization of poison oak urushiol.
J Pharm Sci 64, 1715-1718, 1975.
4) Elsohly MA, Turner CE. GLC analysis of poison ivy and poison oak urushiol components in vegetable oil preparations.
J Pharm Sci 69, 587-589, 1980.
5) Niimura N, Miyakoshi T. Structural study of oriental lacquer films during the hardening process. Talanta 70, 146-152, 2006.
6) Kalish RS, Johnson KL. Enrichment and fuction of urushiol (poison ivy)-specific T lymphocytes in lesions of allergic contact dermatitis to urushiol. J Immunol 145, 3706-3713, 1990.
7) Kalish RS. Recent developments in the pathogenesis of allergic contact dermatitis. Arch Dermatol 127, 1558-1563, 1991.
8) Kalish RS, Wood JA, LaPorte A. Processing of urushiol (poi- son ivy) hapten by both endogenous and exogenous pathways for presentation to T cell in vitro. J Clin Invest 93, 2039- 2047, 1994.
9) Hong DH, Han SB, Lee CW, Park SH, Jeon YJ, Kim MJ, Kwak SS, Kim HM. Cytotoxicity of urushiols isolated from sap of Korean lacquer tree (Rhus vernicifera Stokes). Arch Pharm Res 22, 638-641, 1999.
10) Lee JH, Yim SH, Won YJ, Jung KW, Son BH, Lee HD, Lee ES, Yoo KY, Ahn SH, Shin HR. Population-based breast cancer statistics in Korea during 1993-2002: incidence, mortality, and survival. J Korean Med Sci 22, S11-16, 2007.
11) Brown JM, Wouters BG. Apoptosis: mediator or mode of cell
killing by anticancer agents? Drug Resist Updat 4, 135-136, 2001.
12) Fesik SW. Promoting apoptosis as a strategy for cancer drug discovery. Nat Rev Cancer 5, 876-885, 2005.
13) Li Y, Ahmed F, Ali S, Philip PA, Kucuk O, Sarkar FH.
Inactivation of nuclear factor kappaB by soy isoflavone genistein contributes to increased apoptosis induced by chemotherapeutic agents in human cancer cells. Cancer Res 65, 6934-6942, 2005.
14) Hsu YL, Kuo PL, Lin LT, Lin CC. Asiatic acid, a triterpene, induces apoptosis and cell cycle arrest through activation of extracellular signal-regulated kinase and p38 mitogen-activated protein kinase pathways in human breast cancer cells. J Pharmacol Exp Ther 313, 333-344, 2005.
15) Boise LA, Gonzaliez-Garcia M, Postema CE, Ding I, Linsten T, Turka LA, Mao X, Nunez G, Thompson CB. bcl-x, a Bcl-2 related gene that functions as a dominant regulator of apo- ptotic cell death. Cell 74, 597-608, 1993.
16) Oltval Z, Milliman CL, Korsmeyer SJ. Bcl-2 heterodimerized in vivo with a conserved homology, Bax, that accelerates pro- grammed cell death. Cell 74, 609-619, 1993.
17) Gottschalk AR, Boise LH, Thompsin CB, Quintans J. Iden- tification of immunosuppressant-induced apoptosis in murine B-cell line and its prevention by bcl-x but not Bcl-2. Proc Natl Acad Sci USA 91, 7350-7354, 1994.
18) Tada H, Shiho O, Kuroshima K, Koyoma M, Tsukamoto K.
An improved colorimetric assay for interleukin-2. J Immunol Methods 93, 157-165, 1986.
19) Dermers GW, Foster SA, Halbert CL, Galloway DA. Growth arrest by induction of p53 in DNA. Proc Natl Acad Sci USA 91, 4382-4386, 1994.
20) Hartwell LH, Kastan MB. Cell cycle control and cancer.
Science 266, 1821-1828, 1994.
21) Salomons GS, Brady HJ, Verwijs-Janssen M, van den Berg JD, Hart AA, van den Berg H, Behrendt H, Hahlen K, Smets LA. The Bax alpha:Bcl-2 ratio modulates the response to dexamethasone in leukaemic cells and is highly variable in childhood acute leukaemia. Int J Cancer 71, 959-965, 1997.
22) Choi HJ, Choi YH, Yee SB, Im E, Jung JH, Kim ND.
Ircinin-1 induces cell cycle arrest and apoptosis in SK-MEL-2 human melanoma cells. Mol Carcinog 44, 162-173, 2005.
23) Roy N, Deveraux QL, Takahashi R, Salvesen GS, Reed JC.
The c-IAP-1 and c-IAP-2 proteins are direct inhibitors of specific caspases. EMBO J 16, 6914-6925, 1997.
24) Lacroix M, Toillon RA, Leclercq G. p53 and breast cancer, an update. Endocr Relat Cancer 13, 293-325, 2006.