99 책임저자:박옥진, 305-811, 대전시 유성구 전민동 461-6
한남대학교 식품영양학과
Tel: 042-629-8793, Fax: 042-629-8789 E-mail: [email protected]
접수일:2009년 6월 3일, 게재승인일:2009년 6월 18일
Correspondence to:Ock Jin Park
Department of Food and Nutrition, Hannam University, 461-6, Jeonmin-dong, Yusung-gu, Daejeon 305-811, Korea
Tel: +82-42-629-8793, Fax: +82-42-629-8789 E-mail: [email protected]
Regulation of AMP-activated Kinase and
Cyclooxygenase-2 by Estrogen in Non-cancerous, Tumor-promoter Stimulated Non-cancerous or
Breast Cancer Cells
Song Yi Park1, Yun-Kyoung Lee2, Young-Min Kim1, Won Sup Lee3 and Ock Jin Park2 Departments of 1Biological Sciences, 2Food and Nutrition, Hannam University, Daejeon 305-811,
3Department of Internal Medicine, Gyeong-Sang National University Hospital, Jinju 660-702, Korea
Estrogen has been suggested as a promoter of breast cancer for its cell proliferative effect and antiapoptotic properties, and moreover, recent epidemiological data suggest that females are at higher risk for lung cancers than males, and it is attributed to the greater exposure to the genotoxicity of estrogen metabolites in the form of hydroxyestradiols. Our previous studies have revealed that AMP-activated protein kinase (AMPK) activation is strongly related to the inhibition of cancer cell growth. We examined the regulation of AMPK and cyclooxygenase-2 (COX-2) by exogenous estrogen in non-cancerous normal cells, TPA (12-O-tetradecanoylphorbol-13-acetate)-treated normal cells and MCF-7 breast cancer cells to elucidate the effects of estrogen treatment on AMPK and COX-2. The exogenous estrogen strongly stimulated COX-2 expression in TPA treated cox-2 +/+ cells and MCF-7 cells, but such stimulation was not observed with non-TPA treated normal cells. Estrogen activated AMPK in all three kinds of cells, and in TPA-treated and cancerous cells the activation of AMPK with estrogen could not abolish COX-2 expression. These results indicate that the cell type is an important defining factor of AMPK and COX-2 regulation by estrogen, and this may have some significance to unravel the possible link between estrogen and cancers. (Cancer Prev Res 14, 99-103, 2009)
Key Words: Estrogen, TPA treatment, AMPK-COX-2 pathway, Carcinogenesis, MCF-7 breast cancer cells
INTRODUCTION
The greater exposure to estrogen is known to play an im- portant role in the development of breast tumors.1) The concept that estrogens are the group of hormones capable of promoting cell proliferation makes these hormones as strong candidates for major risk factors of breast carcinogenesis.2) Recent epidemio- logical studies have revealed that non-smoking women may be at greater risk for lung cancer than non-smoking men, possibly due to the greater exposure to hydroxyestradiols, a carcinogenic
metabolite of estrogen.3) Gene expression alteration by estrogen is controlled by cell- and gene-specific interactions, along with several regulatory compound factors. Among various epigenetic factors, estrogen receptors are the elements responsible for vari- ous gene expression. Once estrogen binds to the receptor (particularly b-receptor), the binding of the receptor to the cog- nate estrogen-responsive element (ERE) occurs and this event would modulate target gene expression.4) Although many at- tempts to delineating various epigenetic interactions caused by estrogen exposure, the nature of epigenetic events connecting estrogen to carcinogenesis has not been clarified. Among vari-
Fig. 1. Effects of 17-β-estradiol on proliferation of MCF-7 breast cancer cells. Cells were exposed to 17-β-estradiol for 24 or 48 hr and cell survival was measured by MTT assay.
ous candidates of target gene modulation by estrogen, the con- tribution of COX-2 alteration by estrogen can draw much attention.5)
COX-2 is one of two distinct isoenzymes COX-1 and COX-2, and one of mediators of inflammation.6) Inflammation is known as a key factor in carcinogenesis, especially in stages of promotion and/or progression.7) It was found that the prod- uct of COX-2 pathway or COX-2 itself is capable of increasing cell proliferation, and enhancing anti-apoptotic potential of tu- mor cells.8) Furthermore, COX-2 may play a critical role in cancer development by modulating key steps of cancer cell adaptation, such as angiogenesis and migration.9) Our labo- ratory has reported that certain dietary factors including sele- nium and tea catechin modulate cancer cell proliferation by re- ducing COX-2 expression through a recently emerging carcino- gensis master controller, AMPK.10,11) The various observations such as the overexpression of COX-2 or the elevated prosta- glandin levels in solid malignancies, the potential of COX-2 to increase cell proliferation and decrease apoptosis or to induce angiogenesis, the abrogation of experimental tumorigenesis in cox-2 knock out animals or application of certain prostaglandin product to enhance skin tumorigenesis reveals a close link be- tween inflammation and carcinogenesis.12) Therefore, the rela- tionships among estrogen exposure, inflammatory conditions el- evating COX-2 and cancer control need to be scrutinized.10) Furthermore, the involvement of AMPK in inflammation and tumorigenesis merits precise investigation. Currently, AMPK draws attention for its possible role of the primary target for the prevention and treatment of cancers, since AMPK has been shown to be tightly connected to tumor suppressor genes such as LKB and TSC2 (tuberous sclerosis complex 2).13)
Present study was undertaken to unravel the epigenetic mechanism of carcinogensis induced by estrogen or possibly es- trogen metabolites. Carcinogenic effects of estrogen through modulating AMPK or COX-2 were examined, and estrogen in the form of 17-β-estradiol was applied to normal cells under TPA treatment, compared to the treatment of estrogen to can- cerous cells.
MATERIALS AND METHODS 1. Cell culture
The MCF-7 human breast cancer cell line was purchased from ATCC (Gaithersburg, MD). The cells of cox-2+/+ and
cox-2−/− MEFs were generously provided by Dr. Z. Dong (Hormel Institute, Austin, MN). The cells were cultured in RPMI1640 medium containing 10% FBS, 100 mg/l streptomy- cin, and 100 U/ml penicillin.
2. Cell proliferation assay
Cell proliferation was assessed by the MTT method. Cells were seeded in 24-well plates containing the test compounds for indicated dose dependently, and then incubated with 30μl MTT solution (5 mg/ml in PBS) for 2 h at 37oC. Optical den- sities of the solutions, in each well, were determined by an ELISA reader.
3. Western blotting
Cells were lysed with ice-cold lysis buffer, including 50 mM Tris-HCl, pH 7.4, 1% NP-40, 0.25% sodium deoxycholate, 150 mM NaCl, 1 mM EDTA, 1 mM PMSF, 1 mM sodium orthovanadate, 1 mM NAF, 1μg/ml aprotinin, 1μg/ml leu- peptin, and 1μg/ml pepstatin. Solubilized proteins were centri- fuged at 13,000 xg in a microfuge (4oC) for 5 min, and super- natant protein was collected. The cell lysates were separated on SDS-PAGE and then electro-transferred onto a nitro- cellulose membrane (Scheicher & Schuell, Dassel, Germany).
Antibodies (p-Acc, p-AMPK, COX-2, and β-actin) were pur- chased from Cell Signaling Technology (Danvers, MA, USA).
Fig. 2. Effects of 17-β-estradiol on COX-2 expression and AMPK activation in MCF-7 breast cancer cells. Cells were exposed to 17-β-estradiol for 24 hr and 48 hr, and p-AMPK and COX-2 levels were measured.
Fig. 3. Effects of 17-β-estradiol on COX-2 expression and AMPK activation in cox-2 +/+ and negative cell. Cells were exposed to 17-β-estradiol for 6 hr, and p-AMPK and COX-2 levels were measured.
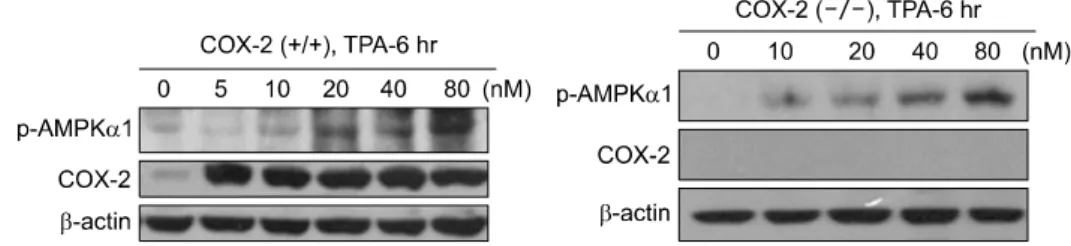
Fig. 4. Effects of TPA on COX-2 expression and AMPK activation in cox-2 +/+ and negative cell. Cells were exposed to TPA for 6hr and p-AMPK and COX-2 levels were measured.
RESULTS
1. Effects of 17 β-estradiol on MCF-7 cell growth
The evaluation of the effect of estrogen on the growth of MCF-7 cells is shown in Fig. 1. A dose-dependent growth stimulation was observed when the cancer cells were treated with 17 β-estradiol (0∼200 nM) for 24 or 48 hr.
2. Effects of 17 β-estradiol on COX-2 ex- pression and AMPK activation in MCF-7 cells
As shown in Fig. 2 estrogen stimulated COX-2 expression both of the incubation periods of 24 and 48 hr. A striking activation of AMPK by estrogen was found at 100∼200 nM (24 hr incubation) and 25∼200 nM concentrations of estrogen treatment. The activated patterns of COX-2 and AMPK by estrogen were different, since AMPK was activated only at the higher concentration in 24 hr treatment, and similar in 48 hr treatment.
3. Effects of 17 β-estradiol on COX-2 ex- pression and AMPK activation in cox-2 +/+
and cox-2−/− cells
To examine the effect of estrogen on COX-2 and AMPK regulation in normal cells possessing cox-2 gene or lacking cox-2
gene, AMPK and the activation of COX-2 were detected in these cells treated with 0∼200 nM 17 β-estradiol (Fig. 3).
In both of cox-2 +/+ and cox-2−/− cells, AMPK activation occurred showing a slight dose dependency. It was found that the activation of AMPK did not appear to be related to COX-2 expression in cox-2 +/+ cells. It seems that estrogen possesses AMPK activating ability exhibiting a strong response at higher concentrations, and COX-2 stimulatory effect only at the high- est concentration in noncancerous cox-2 +/+ cells.
4. Effects of TPA on COX-2 expression and AMPK activation in cox-2 +/+ and cox-2−/− cells
A strong activation of COX-2 was observed in TPA-treated cox-2 +/+ cells (Fig. 4). This activation did not accompany the activation of AMPK in cox-2 +/+ cells, although a slight activation of AMPK at the concentrations between 20∼80 nM
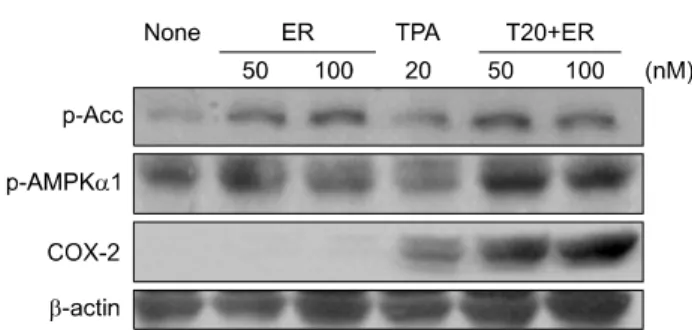
Fig. 5. Effects of 17-β-estradiol on COX-2 expression and AMPK activation in cancer promoter TPA treated cox-2 +/+
cells. Cells were exposed to 17-β-estradiol in the presence of 20 nM of TPA for 6 hr, Western blotting was performed as described in Materials and Methods.
TPA was noticeable. A similar activation of AMPK was found with TPA treatment in cox-2−/−cells as in cox-2+/+ cells (20∼80 nM TPA treatment).
5. Effects of 17-β estradiol on COX-2 ex- pression and AMPK activation in cancer promoter TPA treated cox-2 +/+ cells
As shown in Fig. 5, the combination treatment of TPA and estrogen resulted in a stronger response of COX-2 compared to estrogen alone or TPA alone. At the same time, AMPK acti- vation seemed to be in the order of TPA alone<estrogen alone
<TPA and estrogen.
DISCUSSION AND CONCLUSION
Estrogen has been suggested as a promoter of breast cancer for its cell proliferative effect and an anti-apoptotic agent.14,15) The previous research establishing link between estrogen and breast carcinogenesis revealed that the phenotypic and genomic changes triggered by estrogen exposure that lead normal cells to tumorigenesis.16) Generally, COX-2 and other proteins in the same pathway play an important role as tumor growth modulators. The accumulating evidence presented there are in- verse relationship between COX-2 inhibition and breast cancer incidence.17) A number of studies supports that overexpression of COX-2 is linked to breast tumors. The exact molecular mechanisms involving COX-2 responsible for induction of tu- morigenesis have not unraveled currently. Most likely ex- planation would be interaction of prostanoids with induction of proliferatory responses such as growth stimulatory kinases or acceleration of anti-apoptotic pathways.18) The activation of
nuclear receptors such as NFkB may be triggered by prosta- noids resulting in pro-mitogenic responses.
In this study, we have assessed the possible link between estrogen, and COX-2 expression, AMPK and the alteration of cancer cell growth. We applied the three kinds of cell sys- tems-noncancerous cells, tumor promoter TPA treated normal cells and breast cancer cells. This study clearly showed that the alteration of COX-2 expression by estrogen was found only in cancer cells or TPA cells, and this provides persuasive evidence that estrogen may control cancer cell proliferation via COX-2 expression. Estrogen showed the ability to activate AMPK in all three noncancerous or cancer cells. However, the activation of AMPK was not accompanied with the suppression of COX-2 in TPA-treated or cancerous cells, indicating that AMPK might not act as an anti-cancer target in the systems treated with estrogen. The importance of AMPK-COX-2 signaling pathway in controlling cancer cells has been emphasized in pre- vious publication.11)
We have investigated the potential of estrogen to inducing COX-2 in three kinds of cell systems of noncancerous normal cells, tumor-promoter treated cells and cancerous cells. And we have tested the possible link between cancer cell growth control and AMPK-COX-2 signaling pathway. It was found that estro- gen has an ability to increase COX-2 expression in cancer cells or tumor-promoter treated cells. In contrast to other phytoes- trogen, the stimulatory activity of estrogen on AMPK was not related to the control of COX-2 expression. The unique ability of estrogen to activate AMPK in a situation of cell growth stimulating requires further study. One of the possible ex- planations of this unexpected finding is that although estradiol has a potential to phosphorylate AMPK in all kinds of cells including cancer cells, a strong growth stimulatory effect of es- trogen possibly via signals such as akt might hinder the ability of AMPK to suppress COX-2 in cancer cells.
ACKNOWLEDGEMENT
This work was supported by Hannam University Fund 2009.
REFERENCES
1) Helferich WG, Andrade JE, Hoagland MS. Phytoestrogens and breast cancer: a complex story. Inflammopharmacology 16, 219-226, 2008.
2) Chen GG, Zeng Q, Tse GM. Estrogen and its receptors in cancer. Med Res Rev 28, 954-974, 2008.
3) Ho CC, Ling YC, Chang LW, Tsai HT, Tsai MH, Lin P.
17-Beta estradiol and hydroxyestradiols interact via the NF-kappa B pathway to elevate cyclooxygenase 2 expression and prostaglandin E2 secretion in human bronchial epithelial cells. Toxicol Sci 104, 294-302, 2008.
4) Zhang Y, Zhou G, Wang H, Zhang X, Wei F, Cai Y, Yin D. Transcriptional upregulation of breast cancer resistance protein by 17beta-estradiol in ERalpha-positive MCF-7 breast cancer cells. Oncology 71, 446-455, 2006.
5) Akarasereenont P, Techatraisak K, Thaworn A, Chotewutta- korn S. The induction of cyclooxygenase-2 by 17beta-estradiol in endothelial cells is mediated through protein kinase C.
Inflamm Res 49, 460-465, 2000.
6) Chung HY, Cesari M, Anton S, Marzetti E, Giovannini S, Seo AY, Carter C, Yu BP, Leeuwenburgh C. Molecular in- flammation: underpinnings of aging and age-related diseases.
Ageing Res Rev 8, 18-30, 2009.
7) Fitzpatrick FA. Inflammation, carcinogenesis and cancer. Int Immunopharmacol 1, 1651-1667, 2001.
8) Fecker LF, Stockfleth E, Nindl I, Ulrich C, Forschner T, Eberle J. The role of apoptosis in therapy and prophylaxis of epithelial tumours by nonsteroidal anti-inflammatory drugs (NSAIDs). Br J Dermatol 156, 25-33, 2007.
9) Greenhough A, Smartt HJ, Moore AE, Roberts HR, Williams AC, Paraskeva C, Kaidi A. The COX-2/PGE2 pathway: key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis 30, 377-386, 2009.
10) Modugno F, Ness RB, Chen C, Weiss NS. Inflammation and endometrial cancer: a hypothesis. Cancer Epidemiol Biomarkers Prev 14, 2840-2847, 2005.
11) Hwang JT, Kim YM, Surh YJ, Baik HW, Lee SK, Ha J, Park OJ. Selenium regulates cyclooxygenase-2 and ex- tracellular signal-regulated kinase signaling pathways by acti- vating AMP-activated protein kinase in colon cancer cells.
Cancer Res 66, 10057-10063, 2006.
12) Rundhaug JE, Fischer SM. Cyclo-oxygenase-2 plays a critical role in UV-induced skin carcinogenesis. Photochem Photobiol 84, 322-329, 2008.
13) Hardie DG. New roles for the LKB1→AMPK pathway. Curr Opin Cell Biol 17, 167-173, 2005.
14) Nenci I, Marchetti E, Querzoli P. Commentary on human mammary preneoplasia. The estrogen receptor-promotion hypothesis. J Steriod Biochem 30, 105-106, 1988.
15) Perillo B, Sasso A, Abbondanza C, Palumbo G. 17 beta-estra- diol inhibits apoptosis in MCF-7 cells, including bcl-2 ex- pression via two estrogen-responsive elements present in the coding sequence. Mol Cell Biol 20, 2890-2901, 2000.
16) Huang Y, Fernandez SV, Goodwin S, Russo PA, Russo IH, Sutter TR, Russo J. Epithelial to mesenchymal transition in human breast epithelial cells transformed by 17beta-estradiol.
Cancer Res 67, 11147-11157, 2007.
17) Mazhar D, Ang R, Waxman J. COX inhibitors and breast cancer. Br J Cancer 94, 346-350, 2006.
18) Schams D, Berisha B. Steroids as local regulators of ovarian activity in domestic animals. Domest Anim Endocrinol 23, 53-65, 2002.