Introduction
Dodam herbal formula (Dodam) is a well-known Korean traditional prescription, which has been widely used in oriental medicine for treatment of heart and brain disorders1). However, the mechanism by which Dodam prevents functional deterioration of brain cells after acute stroke is still unclear.
Cellular oxidative damage is implicated a wide array of clinical disorders. Increased oxidative stress has been implicated as the leading cause for neuronal cell death in many neurodegenerative disorders including stroke2). Due to the high consumption of oxygen, high levels of polyunsaturated fatty acids in neural cell membranes and the relatively low levels of antioxidants, the brain is particularly vulnerable to oxidative insults3,4).
Mitochondria are essential cellular organelles that play central roles in energy metabolism and apoptosis. The mitochondrial respiratory chain is one of the most important
sites of ROS production under physiological conditions, and it has been long suspected that mitochondrial ROS play an important role in apoptosis. Distortion of the neuronal oxidative metabolism correlating with suppressed activity of the respiratory chain enzymes has been demonstrated for a number of neurological diseases. Decreases in mitochondrial respiratory chain complex activities as well as mitochondrial-derived oxidative stress have been implicated in a number of diseases such as those related to neurodegenerative and chemical-induced cells injuries. It is now generally accepted that massive neuronal death due to oxidative stress is a common characteristic of brain whether in stroke1,5) or in neurodegenerative diseases including Alzheimer’s disease6), Parkinson’s disease7,8), and traumatic brain injury9). Complex I could be a critical site of mitochondrial ROS production and relatively small level of inhibition is sufficient to increase ROS generation10,11). Inhibition of the mitochondrial respiratory chain by rotenone has been widely used to study the role of the mitochondrial respiratory chain in apoptosis12,13). Rotenone is a naturally occurring insecticide isolated from plant species belonging to genera Derris and Lonchorcarpus.
The cytotoxicity of rotenone is irreversible binding and inactivation of NADH dehydrogenase complex (generally
Protective Effects of Dodam Water Extract (Dodam) Against Rotenone-Induced Neurotoxicity in Neuro-2A Cells
Myung Ja Youn, Seong Yeol Park, Channy Park, Jin Kyung Kim, Yunha Kim, Eun Sook Kim, Byung Soon Moon1, Hong Seob So, Raekil Park*
Vestibulocochlear Research Center & Department of Microbiology, School of Medicine, 1 : School of Oriental Medicine, Wonkwang University
Dodam formula (Dodam) has been used for neurodegenerative disease in Oriental medicine. Dodam is capable of protecting diverse kinds of cells from damage caused by a variety of toxic stimuli. In the present study, we investigated the underlying protective mechanism of Dodam on rotenone-induced cytotoxicity in rat neuroblastoma Neuro-2A cells. Treatment with Neuro-2A cells with rotenone caused the loss of cell viability, and condensation and fragmentation of nuclei, which was associated with the elevation of ROS level, and lipid peroxidation, the increase in Bax/Bcl-2 ratio. Rotenone induced mitochondrial dysfunction characterized by mitochondrial membrane potential loss and cytochrome-c release. These phenotypes induced by rotenone were reversed by pretreatment with Dodam. Our results suggested that major features of rotenone-induced neurotoxicity are partially mediated by mitochondrial dysfunction and oxidative stress, and that Dodam markedly protects Neuro-2A cells from oxidative injury. These data indicated that Dodam might provide a useful therapeutic strategy in treatment of the neurodegenerative diseases caused by oxidative injuries.
Key words : Dodam, Rotenone Neurotoxicity, Mitochondrial dysfunction, Neuro-2A
* To whom correspondence should be addressed at : Raekil Park, VestibuloCochlear Research Center and Department of Microbiology, Wonkwang University School of Medicine, 344-2 Shinyong-dong, Iksan,
․E-mail : [email protected], ․Tel : 063-850-6777
․Received : 2008/03/20 ․Accepted : 2007/04/01
known as complex I) of the mitochondrial electron transport chain. Inhibition of mitochondrial respiratory chain complex I by rotenone had been found to induce cell death in a variety of cells. Rotenone is considered to exert its cytotoxicity through induction of apoptosis14,15).
The present study, we describe the protective effect of Dodam on neuronal cell death induced by an inhibitor of mitochondrial electron transport, rotenone. We used Neuro-2A cells as a model to investigate the role of oxidative stress to neuronal apoptosis induced by rotenone. Our data demonstrate that rotenone induced mitochondrial dysfunction with ROS production, which results in apoptotic death of Neuro-2A cells.
Materials and Methods
1. Reagents
Dulbecco’s modified Eagle’s medium (DMEM) was from Gibco Life Technologies (USA). Fetal bovine serum (FBS) was purchased from Hyclone. 3-[4, 5-dimethylthiazol-2-yl]-2, 5-dephenyl tetrazolium bromide (MTT), trypsin, streptomycin, penicillin, and rotenone were obtained from Sigma. 2’
7’-Dichlorofluorescein diacetate (DCFH-DA), JC-1, and hydroethidium (HE) were purchased from Molecular Probes (Eugene, OR, USA). Antibodies for Bcl-2, Bax, cytochrome-c, VDAC and β-actin were purchased from Santa Cruz Biotechnology Inc. Peroxidase-linked secondary antibody was bought from Amersham Life Science. All other chemicals used were of the highest grade available commercially.
2. Preparation of Dodam Water Extract
The water extract of Dodam herbal formula (Dodam)was prepared as bellows. The dried prescription of herbs, provided by Wonkwang Oriental Medical Hospital, was boiled with distilled water for 3 h. Total extract was centrifuged at 5,000 rpm for 30 min and the supernatant of extract was lyophilized and kept at 4℃. Lyophilized extract was weighed and dissolved in dimethyl sulfoxide (DMSO). The ingredients of 49.2 g Dodam herbal formula included 8 g Tuber Pinelliae, 4 g Pericarpium Citri Nobilis, 4 g Poria, 4 g Rhizoma arisaematis, 4 g Fructus Ponciri, 6 g Radix Linderae, 4 g Rhizoma Cnidii, 4 g Radix Angelicae Dahuricae, 4 g Batryticatus Bambycis, 4 g Radix Platycodi, 2 g Rhizoma Zingiberisa Siccatum, 1.2 g Radix Glycyrrhizae. The Dodam water extract was identified and authenticated by College of Oriental Medicine, Wonkwang University (Iksan, Jeonbuk, Korea).
3. Cell culture
Neuro-2A cells (American Type Culture Collection, USA)
were maintained in DMEM supplemented with 10% (v/v) heat-inactivated fetal calf serum and 100 units/ml penicillin/streptomycin. Cells were kept at 37℃ in humidified 5% CO2 and 95% air. Experiments were carried out 24 - 48 h after cells were seeded onto plates or dishes.
4. MTT assay for cell viability
Cell viability was quantified by MTT assay. Briefly, cells were plated in 48-well culture plates at the density of 5 × 104 cells/well and allowed to adhere at 37℃ for 12 h. Thereafter, medium was replaced with fresh medium and cells were incubated with rotenone in the presence or absence of Dodam. After 24 h, 50 ml MTT (5 mg/mL in PBS) were added and cells incubated for an additional 4 h at 37℃. The absorbance was then measured at 595 nm using a Digiscan Micro plate Reader (Assays Hitech, Kornenburg, Austria). Wells without cells were used as blanks and were subtracted as background from each sample. Results were expressed as a percentage of control.
5. Apoptosis assay
To assess apoptosis, the nuclei of Neuro-2A cells were stained with DAPI. Cells were fixed in PBS containing 3.7%
paraformaldehyde for 15 min. After fixation, cells were washed twice with PBS and then treated with DAPI (1: 5,000 dilutions) in PBS at room temperature for 15 min. After three washes, cells were observed under fluorescence microscope. To prepare genomic DNA, Neuro-2A cells (4 x 106, incubated for 24 h with or without rotenone, alone, or in presence of Dodam were detached from 6 cm culture dishes and the cell suspension was centrifuged at 100 g for 10 min16). The cell pellet was then washed twice with ice-cold PBS and genomic DNA was isolated using a DNA isolation kit (Promega, Basel, Switzerland) according the manufacturer’s instructions. The DNA samples was analyzedon a 1.5% agarose gel containing ethidium bromide (1 mg/mL) in TBE buffer (100 mM Tris, 90 mM boric acid, 1 mM EDTA) and run for 90 min at 70 V.
After electrophoresis, the DNA was visualized under UV light and photographed.
6. Caspase activity assay
To measure the activity of caspase-3 in Neuro-2A cells, a colorimetric assay was used according to the instruction of manufacturer. Briefly, cells were treated with indicated conditions and at the indicated times, cells were washed once in cold PBS and lysed in a buffer containing 50 mM HEPES (pH 7.4), 100 mM NaCl, 0.5% CHAPS (pH 7.4), 1 mM DTT, and 0.1 mM EDTA. Cells lysate was incubated with peptide substrate in assay buffer for 2 h at 37°C. Cleavage of
fluorogenic substrates, including 100 mM Ac-DEVD-AMC by caspase-3 was measured at an excitation of 380 nm and an emission of 460 nm. Results are represented as the percent change of the activity compared to the untreated control.
7. Determination of Mitochondrial Membrane Potential (MMP) Mitochondrial membrane potential was quantified using the ratio metric probe JC-1 (Molecular Probes, USA). In healthy Neuro-2A cells, the intact membrane allows the lipophilic dye JC-1 to enter into the mitochondria where it accumulated and aggregated, producing an intense orange signal. In apoptotic cells, where the mitochondrial membrane potential has collapsed, the monomeric JC-1 remains in the cytosol and appears green. Neuro-2A cells were treated for 15 h with 500 nM rotenone in the absence or presence of Dodam. Cells were incubated for 30 min with 10 mg/ml JC-1 at 37℃ under 5%
CO2, and then washed twice with 0.1 M PBS. The proportion of aggregated vs. monomeric JC-1 probe was visualized under UV light and photographed.
8. Mitochondrial complex I activity (NADH–ubiquinone oxidoreductase assay)
Mitochondria complex I activity was determined by the method of Ian et al.,12), which followed the decrease in NADH absorbance at 340 nm that occurred when ubiquinone (CoQ1) was reduced to form ubiquinol17). The reaction was initiated by adding CoQ1 (50 mM) to the reaction mixture, at 30℃. After 5 min, rotenone was added and the reaction was further registered for 5 min. Complex I activities were expressed in nanomoles per minute per milligram of protein and represent the rotenone sensitive rates.
9. Western blot analysis
Cytosolic protein extracts was prepared as previously described18). Briefly, cells were collected by centrifugation at 300 x g for 5 min at 4°C and washed with ice-cold PBS. The cell pellet was then resuspended in 500 ml of lysis buffer (20 mM Hepes- KOH, pH 7.5, 210 mM sucrose, 70 mM mannitol;
1.5 mM MgCl2, 10 mM KCl2, 10 ug/ml leupeptin, and 10 mM digitonin). After 10 min incubation at 25°C, the sample was spun at 14,000 x g for 15 min, and the supernatant containing cytosolic proteins was stored at -70°C until analysis by polyacrylamide gel electrophoresis (SDS-PAGE). The protein extract wassubjected to standard SDS-PAGE, transferred onto polyvinylidene difluoride membranes (Millipore), and probed with appropriate antibodies as described individually in the figure legends. The bound primary antibody was detected using appropriate horseradish peroxidase-conjugated secondary
antibody, and protein was visualized using an enhanced chemiluminescence detection kit. β-actin was used as an internal control to confirm that the amounts of protein load were equal.
10. Detection of cytochrome-c release
Cytochrome-c released from mitochondria was determined by Western blot analysis. Cytosolic and mitochondrial fractions were prepared as below, but the fractionation buffer contained 250 mM sucrose, 10 mM KCl, 1.5 mM MgCl2, 1 mM dithiothreitol, 1 mM EDTA, 1 mM EGTA, 20 mM HEPES, pH 7.0, and protease inhibitors (1 mM sodium orthovanadate, 10 mg/ml leupeptin, 10 mg/ml aprotinin, and 1 mM phenylmethylsulfonyl fluoride). The mitochondrial pellet was resuspended in the same buffer, sonicated, and centrifuged to obtain the soluble fraction. Equal amounts of protein were separated on a 10% SDS polyacrylamide gel and transblotted onto polyvinylidene difluoride-nitrocellulose filters. Specific cytochrome-c band was detected by using anti-cytochrome-c antibody (1:500) followed by enhanced chemiluminescence-based detection.
11. Measurement of intracellular ROS
ROS generation in cells was measured by using fluorescent probes 2’ 7’-Dichlorofluorescein diacetate (DCFH-DA, Molecular Probes, Eugene, OR, USA) and hydroethidium (HE). Cells in 96-well plates were incubated for 24 h, reacted with 10 mM DCF-DA at 37 ℃for 30 min, and washed twice with PBS. Fluorescence intensity of DCF was measured by using microplate-reader (Fluoroskan Ascent Thermo Labsystems) at excitation wavelength 485 nm and emission wavelength 538 nm. Blue fluorescent dihydroethidium is oxidized to red fluorescent ethidium in the presence of superoxide. Therefore, the ethidium fluorescence intensity is proportional to the amount of superoxide formed intracellulary. Cells were washed twice with PBS and incubated in Krebs Ringer HEPES solution containing 5 mM HE at 37℃ for 30 min. The fluorescence was measured by flow cytometry (FACSCalibur, BD Biosciences).
12. Lipid peroxidation (MDA-TBA assay)
Lipid peroxidation assay was based on that described by Ohkawa et al.,19) with some modifications reported by Ford et al.,20). Briefly, 50 ml of PBS suspension of harvested cells (1 - 5 × 106 cells) were added 50 ml of 3.2% SDS in PBS, and incubated for 10 min at room temperature. Following incubation, 150 ml of 20% acetic acid (pH 3.5) and 150 ml of 0.8% TBA (thiobarbituric acid) in 0.05 N NaOH were added and the mixture was then boiled for 1 h. The mixture was
cooled and extracted with 1 ml each of n-butanol-pyridine mixture (1:3, v/v) to avoid turbidity. The upper layer (approximately 240 ml) of each sample was aspirated, and the absorbance was measured at 532 nm. Concentration of 2-TBA was determined using the extinction coefficient of 1.56×105 M-1cm-1. The results were expressed as mol of malondialdehyde (MDA) per mg-protein. Protein was determined by the method of Lowry using bovine serum albumin as a standard.
13. Statistical analysis
The data given in the text are expressed as mean ± S.D.
Statistical comparison was carried out with three or more groups using one-way analysis of variance (ANOVA). A difference was considered to be significant at P< 0.05.
Results
1. Treatment with Dodam protected Neuro-2A cells from Rotenone-induced cytotoxicity
We tested the cytoprotective effects of Dodam on rotenone-induced toxicity in Neuro-2A cells. First, we evaluated the cytotoxicity of rotenone in Neuro-2A cells. As shown in Fig. 1A, cells were exposed to increasing doses of rotenone for 24 h and cell viability was determined MTT assay. Cell viability was markedly decreased 24 h after rotenone treatment in a dose-dependent manner. However, when cells were pretreated with Dodam (0.125 - 1.5 mg/ml), rotenone-induced cytotoxicity was significantly attenuated(Fig.
1B). The protective effect of Dodam was dose-dependent manner with a maximal effects being reached around 1 mg/ml. Thus, Dodam alone seems to be did not cause any cytotoxic effect on the cell viability.
A B
Fig. 1. Effects of Dodam on the Rotenone-induced cytotoxicity in Neuro-2A cells. Cell cytotoxicity was assessed by MTT assay as described in Materials and Methods. A) Cells were treated with variable concentrations of rotenone for 24 h and an MTT assay was carried out. B) Cells were pre-treated with variable concentrations of Dadam for 3 h and then incubated with 0.5 mM rotenone for 24 h. Data are means ± SD of three independent experiments. *p<0.05 compared with cells treated with control, **p<0.005 compared with cells treated with rotenone only.
2. Effects of Dodam on Rotenone-induced apoptosis in Neuro-2A cells
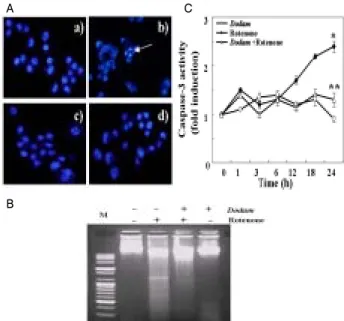
To characterize whether the cell death by rotenone was caused by apoptosis, typical assay was performed. First, morphological change of apoptosis was assessed using DAPI staining. As shown in Fig. 2A-a, the nuclei of control cells had a regular and ovum shape. However, apoptotic nuclei characterized by nuclear condensation and fragmentation, appeared after exposure to 0.5 mM rotenone for 24 h(Fig.
2A-b). Dodam (0.5 mg/ml) pretreatment blocked the rotenone-induced nuclear damage, while the nuclear morphology of cells exposed to Dodamalone was similar to that of untreated cells. Second, we observed DNA fragmentation using agarose electrophoresis. Cells treated with rotenone showed a ladder pattern of discontinuous DNA fragments(Fig. 2B-b). However, Dodam prevented DNA fragmentation by rotenone(Fig. 2B-d).
A C
B
Fig. 2. Effects of Dodam on Rotenone-induced apoptosis in Neuro-2A cells. The cells were pre-treated with 0.5 mg/ml Dodam for 3 h then incubated in the absence or in the presence of 0.5 mM rotenone for 24 h. A) Effects of Dodam in rotenone-induced chromatin condensation. Nucleus morphologic changes detected by DAPI stain: a) Untreated; b) Rotenone only; c) Dodam and rotenone; d) Dodam only. B) Detection of DNA ladder formation. C) Dodam inhibited rotenone-induced increase in caspase-3 activity. Caspase activity was assayed by a fluoremetric methodusing Ac-DEVD-AMC as substrate. Data are expressed as mean ± SD. *p<0.05 compared with control; **p <0.005 compared with rotenone only.
We also evaluated the activation of capsase-3 in cellular extract of Neuro-2A cells using fluorometric assay as a hallmark of apoptosis. The effect of Dodam on rotenone-induced caspase-3 activation was shown Fig. 2C.
Following 24 h treatment with rotenone, we detected a caspase-3 activity increase to 2.5 folds of the control level.
Pretreatment with Dodam significantly attenuated rotenone-induced caspase-3 activation. Dodam alone did not
show a significant effect on the caspase-3 activity in Neuro-2A cells, which was consistent with its lack of apoptotic response.
3. Treatment with Dodam prevented mitochondrial dysfunction in rotenone-treated Neuro-2A cells
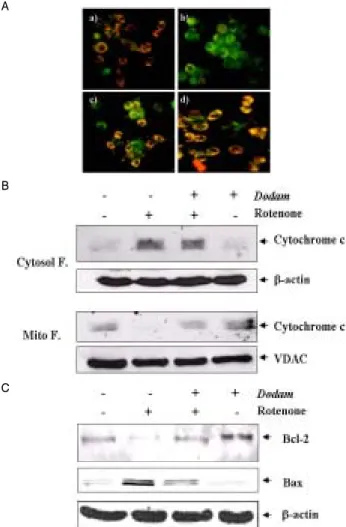
The interruption of electron flow at complex I can lead to reactive oxygen species (ROS) production due to redox cycling of reduced electron carriers upstream of the inhibition sitein mitochondria. If rotenone was acting at complex I to inhibit bioenergetics in Neuro-2A cells, we predict that this action would eventually lead to mitochondrial membrane permeability transition (MMPT), since MMPT can be initiated by ROS21). We first examined the effect of rotenone–induced cell death is mediated via mitochondrial function. A mitochondrial activity marker JC-1 forms monomers of green fluorescence at a low membrane potential, which becomes J-aggregates of orange fluorescence at a higher membrane potential. Fig. 3A revealed that Dodam prevented the rotenone-induced MMPT, while treatment of rotenone resulted in a disruption of MMPT(Fig. 3A-b).
The mitochondrial membrane transition pore opening is associated with collapse of the membrane voltage10), resulting in the release of cytochrome-c into cytosol. Using Western blotting, second, we investigated the possible effects of Dodam on the rotenone-induced cytochrome-c release from mitochondria. As shown in Fig. 3B, rotenone could induce cytochrome-c release significantly. While cytochrome-c protein levels in the cytosolic fraction of the cells treated with Dodam or Dodam plus rotenone for 24 h showed a slightly increase, the protein level was markedly increased in cells treated with rotenone only(Fig. 3B).
The Bcl-2 family consists of both apoptotic and anti apoptotic proteins and the balance between these proteins is critical to turning on and off the cellular apoptotic machinery22). Bcl-2 family members are intimately involved in cell death processes caused by rotenone18). In this study, we investigated whether Dodam has any effect on the expression of Bax and Bcl-2 in rotenone-treated cells using Western blot analysis. As shown in Fig. 3C, Bax protein expression was significantly increased in rotenone-treated cells compared with control cells. However, pretreatment with Dodam (0.5 mg/ml) could decrease the level of Bax expression almost to the normal value. These results suggested a notion that Dodam pretreatment shifted the balance between positive and negative regulators of apoptosis towards cell survival.
In additionally, Complex I and complex III activities were measured in all preparations of mitochondria. As shown in Table 1, mitochondria from rotenone treated Neuro-2A cells exhibited
a marked decrease in the activity of both enzyme complexes, including complex I and complex III, compared with control and Dodam only treated groups. However, Dodam pretreatment had significant protective effects and actually attenuated the decline in the complex I and complex III activities.
A
B
C
Fig. 3. Protective effect of Dodam on mitochondrial dysfunction by Rotenone. A) Effect of Dodam on Rotenone-induced MMP alteration (ΔΨm). Cells treated with indicated condition of above for 15 h were stained with JC-1. a) Untreated; b) Rotenone only; c) Dodam and Rotenone; d) Dodam only. B) Dodam prevents mitochondrial cytochrome c release induced by Rotenone. Cytochrome c release was detected by Western blot analysis. C) Western blot analysis of expression of Bcl-2 family in Neuro-2A cells treated with Rotenone. Cytosol F., cytoplasmic fraction; Mito. F., mitochondrial fraction VDAC, voltage-dependent anion-selective channel protein.
Table 1. Activity of mitochondrial respiratory chain complexes in Neuro-2A cells
Control Rotenone Dodam Dodam+Rotenone Complex Ⅰ 100 60.4 ± 3.98 98.1 ± 2.98 85.0 ± 6.48 Complex Ⅰ+Ⅲ 100 68.2 ± 6.34 99.1 ± 4.32 82.4 ± 2.71 Complex Ⅲ 100 76.7 ± 2.87 94.4 ± 6.02 80.5 ± 6.23 The results are the means ± S.E.M. of duplicate determinations in three different experiments. * P < 0.05, significantly different as compared to control.
4. Treatment with Dodam suppressed the generation of ROS by rotenone in Neuro-2A cells
Rotenone interacts with mitochondrial complex I,
blocking ATP production and promoting oxygen free radical formation23). To determine whether Dodam could prevent the rotenone-induced ROS generation and the resulting oxidative stress, levels of ROS production in cells was determined using the fluorescence probes, DCFH-DA and hydroethidium (HE).
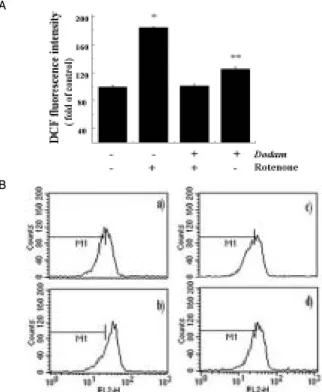
Cells exposed to rotenone for 15 h displayed the increased intensity of DCF-labeled cells compared to control cells. The addition of rotenoneto Neuro-2A cells caused a 2-folds increase in the DCF fluorescence. However, cells pretreated with Dodam showed a reduced intensity of DCF-labeled cells when compared to rotenone-treated cells(Fig. 4A). Fig. 4B demonstrated a typical flow cytometric histogram of hydroethium. The red fluorescence detected in rotenone-treated cells indicated that rotenone indeed caused the generation of superoxide (O2-).
Cells pretreated with Dodam markedly suppressed the intracellular superoxide level induced by rotenone. The results suggested the involvement of ROS in rotenone-induced cytotoxicity and anti-oxidative activity of superoxide.
A
B
Fig. 4. Inhibitory effect of Dodam on ROS production by Rotenone.
The cells were pre-treated with 0.5 mg/ml Dodam for 3 h then incubated in the absence or in the presence of 0.5 mM rotenone for 21 h. The cells were stained with DCFH-DA or hydroethidum (HE) for 30 min. A) The fluorescence intensity of DCF was measured using fluorescent microplate reader. Data are expressed as percentages of DCF fluorescence with respect to control values are mean ± SD. *p<0.05 compared with control; **p <0.005 compared with rotenone only. B) The fluorescence intensity of hydroethidum (HE) was measured using flow cytometry: a) Untreated; b) Rotenone only; c) Dodam and rotenone;
d) Dodam only.
5. Treatment of Dodam inhibited the lipid peroxidation by rotenone in Neuro-2A cells
To determine whether the observed increase in the rate
of lipid peroxidation in the Neuro-2A cells treated with rotenone, we assessed lipid peroxidation by using TBARS assay. Although cells treated with rotenone for 6 h showed no significantly differences in lipid peroxidation according to increasing concentrations of rotenone, when cells were treated with rotenone for 24 h, the rate of lipid peroxidation significantly increased in a dose and time dependent fashion (Fig. 5A). As shown in Fig. 5B, treatment with rotenone for 24 h resulted in showed almost two folds of TBARS formations with respect to control or Dodam only treated cells.
A B
Fig. 5. Effect of Dodam on Rotenone-induced lipid peroxidation. Cells were treated with the indicated conditions of rotenone as described in Material and Methods.
A) Lipid peroxidation was measured in rotenone-treated cells in a time and dose-dependent manner. B) Effectof Dodam in rotenone-induced lipid peroxidation. Data are expressed as mean ± SD of three independent experiments. *p<0.05 compared with control; **p <0.005 compared with rotenone only.
Discussion
The aim of the present investigation was to evaluate the protective effect and to determine the possible molecular mechanisms of Dodam in rotenone-treated Neuro-2A cells. In this study, we demonstrated that Dodam protects Neuro-2A cells against rotenone-induced cytotoxicity through inhibition of mitochondrial dysfunction and activation of caspase cascades.
Mitochondria are considered as a critical mediator of cell death in various neurodegenerative diseases24). Especially, mitochondrial dysfunction caused by reduced activity of either complex I or complex III is known to be associated with traumatic brain injury and a number of neurodegenerative diseases25-28). These electron transport chain dysfunctions can be mimicked using specific blockers, such as rotenone28,29). Complex I can be specifically blocked by rotenone and complex III is blocked by antimycin. Treatment with these blocking agents prevents transfer of electrons down the chain but at the same time accentuates their ability to transfer electrons to oxygen and generate superoxide30-32).
Rotenone interacts with mitochondrial complex I, blocking ATP production and promoting the formation of
oxygen free radicals23). The mitochondrial derived ROS are vital not only because mitochondrial respiratory chain components are present in almost all eukaryotic cells, but also because the ROS produced in mitochondria can readily influence mitochondrial function without having to cope with long diffusion times from the cytosol. Consistently with these reports, several lines of evidence suggest that the mitochondrial membrane potential may be an effector of both the release of cytochrome c and the induction of Bcl-2/Bax14). The loss of ΔΨm is related to the release of apoptogenic molecules, including cytochrome-c and activation of the pro-apoptotic proteins near from the outer mitochondrial membrane. Our present results shown that rotenone induced mitochondria dysfunction, release of cytochrome-c, down-regulation of Bcl-2, and up-regulation of Bax protein in Neuro-2A cells. It also demonstrated that rotenone increased the production of ROS and lipid peroxidation. This is in agreement with a recent report that rotenone induces apoptosis via enhancing the amount of mitochondrial ROS33). Regulation of intracellular ROS and modification of apoptotic cascades may control apoptotic events and provide new strategies for prevention and treatment of neurodegenerative disease.
Degeneration of the nigrostriatal dopaminergic neurons suggest that dopaminergic neurons were more vulnerable to mitochondrial dysfunction induced by rotenone than other neurons29). However, based on the studies about the specificity of rotenone effects has been challenged. Recently, it presumably acts on all neurons but not only on dopaminergic neurons11). It is now believed that damage to non-dopaminergic neurons, involving oxidative stress and/or mitochondrial impairment, culminated in activation of an apoptotic cascade1). These biochemical characteristics suggest a possibility that the mitochondrial respiration chain in neuronal cells may be vulnerable to therapeutic intervention and may serve as a potential target for neurodegenerative disease therapy. Therefore, selective vulnerability to rotenone in neuro-2A cells indicates the presence of intrinsic vulnerability factors unrelated to the dopamine system.
This study showed that Dodam markedly protects neural cells from rotenone. The production of ROS by rotenone ultimately resulted in lipid peroxidation, which is believed to be responsible, in part, for subsequent cellular damage. Our findings clearly demonstrate that rotenone treatment induces oxidative damages, including lipid peroxidation, in Neuro-2A cells and that the cellular redox status are critically important in determination of susceptibility of these cells against inhibition of mitochondrial complex I. However, Dodam markedly suppressed rotenone-induced ROS generation, which
may be one of the underlying mechanisms of neuro-protective effects of Dodam. In conclusion, the present study demonstrates that Dodam herbal formula may have clinical advantages in treatment of neurodegenerative disorders caused by perturbation of redox homeostasis with oxidative stresses.
Acknowledgement
This research was supported by the Program for the Training of Graduate Students in Regional Innovation which was conducted by the Ministry of Commerce Industry and Energy of the Korean Government.
Reference
1. Mattson, M.P., Duan, W., Perdersen, W.A., Culmsee, C.
Neurodegenerative disorders and ischemic brain diseases.
Apoptosis 6: 69-81, 2001.
2. Sun, A.Y., Chen, Y.M. Oxidative stress and neurodegenerative disorders. J Biomed. Sci. 5: 401-414, 1998.
3. Chan, P.H. Reactive oxygen radicals in signaling and damage in the ischemic brain. J Cereb Blood Flow Metab.
21: 2-14, 2001.
4. Love, S. Oxidative stress in brain ischemia. Brain Pathol. 9:
119-131, 1999.
5. Halliwell, B. Role of free radicals in neurodegenerative diseases: therapeutic implications for antioxidant treatment.
Drugs Aging 18: 685-716, 2001.
6. Di Monte, S.M., Sohn, Y.K., Wands, J.R. Correlation of p53- and Fas (CD95)-mediated apoptosis in Alzheimer's disease.
J Neurol Sci. 152(1):73-83, 1997.
7. Jenner, P. Oxidative stress in Parkinson’s disease. Ann Neurol. 53(Suppl. 3):S26-S38, 2003.
8. Tatton, W.G., Chalmers-Redman, R., Brown, D., Tatton, N.
Apoptosis in Parkinson's disease: signals for neuronal degradation. Ann Neurol Suppl. 3: 61-70, 2003.
9. Napieralski, J.A., Raghupathi, R., McIntosh, T.K. The tumor-suppressor gene, p53, is induced in injured brain regions following experimental traumatic brain injury. Mol Brain Res. 71: 78-86, 1999.
10. Seaton, T.A., Cooper, K.M., Schapira, A.H. Free radical scavengers protect dopaminergic cell lines from apoptosis induced by complex I inhibitors. Brain Res. 28(777):110- 118, 1997.
11. Talpade, D.J., Greene, J.G., Higgins, D.S., Greenamyre, J.T.
In vivo labeling of mitochondrial complex I (NADH:
ubiquinone oxidoreductase) in rat brain using [(3)H]dihydrorotenone. J Neurochem. 75(6):2611-2621, 2000.
12. Ian, A.T., Kim, Y.L., Albert, S.J., Wallace, D.C. Assessment of mitochondrial oxidative phosphorylation in patient muscle biopsies, lymphoblast, and transmitochondrial cell lines. Methods Enzymol. 264: 484-509, 1996.
13. Jiménez, M.F.M., Reus, M.I.S., Andres, D., Cascales, M., Benedi, J. Effect of fraxetin on antioxidant defense and stress proteins in human neuroblastoma cell model of rotenone neurotoxicity. Comparative study with myricetin and N-acetylcysteine. Toxicol & Appl Pharm. 209: 214-225, 2005.
14. Armstrong, J.S., Yang, H., Duan, W., Whiteman, M.
Cytochrome c (1) regulates the mitochondrial permeability transition by two distinct pathways. J Biol Chem. 279:
50420-50428, 2001.
15. Isenberg, J.S., Klaunig, J.E. Role of the Mitochondrial Membrane Permeability Transition (MMPT) in Rotenone-Induced Apoptosis in Liver Cells. Toxicol. Sci. pp 340-351, 2000.
16. Gschwind, M., Huber, G. Apoptotic cell death induced by beta-amyloid 1-42 peptide is cell type dependent. J Neurochem. 65(1):292-300, 1995.
17. Lannuzel, A., Michel, P.P., Ho¨glinger, G.U. The mitochondrial complex I inhibitor annonacin is toxic to mesencephalic dopaminergic neurons by impairment of energy metabolism. Neuroscience 121: 287-296, 2003.
18. O’Malley, K.L., Liu, J., Lotharius, J., Holtz, W. Targeted expression of BcL2 attenuates MPP+ but not 6-OHDA induced cell death in dopaminergic neurons. Neurobiol Dis. 14(1):43-51, 2003.
19. Ohkawa, H., Ohishi, N., Yagi, K. Assay of lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 95: 351-358, 1979.
20. Ford, M.S., Maggirwar, S.B., Rybak, L.P., Whitworth, C., Ramkumar, V. Expression and function of adenosine receptors in the chinchilla cochlea. Hear Res. 105: 130-140, 1997.
21. Zhang, J.G., Nicholls-Grzemski, F.A., Tirmenstein, M.A., Fariss, M.W. Vitamin E succinate protects hepatocytes against the toxic effect of reactive oxygen species generated
at mitochondrial complex I and II by alkylating agents.
Chemico-Biol Interactions 138: 67-284, 2001.
22. Cory, S., Adams, J.M. The Bcl2 family: regulators of the cellular life-or-death switch. Nat Rev Cancer 2(9):647-56, 2002.
23. Adams, J.D., Klaidman, L.K., Leung, A.C. MPP+ and MPDP+
induced oxygen radical formation with mitochondrial enzymes. Free Radic Biol Med. 15: 181-186, 2001.
24. Ke, C., Xiaoling, L., Keyi, X., Ven, M. Role of oxidative stress in neurodegenertion: recent developments in assay methods for oxidative stress and naturaceutical antioxidants. Progress in Neuro-Psych. & Biological Psychiaty. 28: 771-799, 2004.
25. Lenaz, G. Role of mitochondria in oxidative stress and aging. Biochim Biophys Acta 1366: 53-67, 1998.
26. Cassarino, D.S., Bennett, J.P. An evaluation of the role of mitochondria in neurodegenerative diseases: mitochondrial mutations and oxidative pathology, protective nuclear responses, and cell death in neurodegeneration. Brain Res.
Rev. 29: 1-2, 1999.
27. Beal, M.F. Energetics in the pathogenesis of neurodegenerative diseases. Trends Neurosci. 23: 298-304, 2000.
28. Orth, M., Shapira, A.H. Mitochondria and degenerative disorders. Am J Med Genet. 106: 27-36, 2001.
29. Betarbet, R., Sherer, T.B., Sherer, T.B., Di Monte, D.A., Greenamyre, J.T. Mechanistic approaches to Parkinson's disease pathogenesis. Brain Pathol. 12(4):499-510, 2002.
30. Chance, B., Sies, H., Boveris, A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 59: 527-605, 1979.
31. Turrens, J.F. Superoxide production by the mitochondrial respiratory chain. Biosci Rep. 17: 3-8, 1997.
32. Nicholls, D.G., Budd, S.L. Mitochondria and neuronal survival. Physiol Rev. 80: 315-360, 2000.
33. Li, N., Ragheb, K.E., Lawler, G., Sturgis, J., Rajwa, B., Melendez, J.A., Robinson, J.P. Mitochondrial complex I inhibitor rotenone induces apoptosis through enhancing mitochondrial reactive oxygen species production. J Biol.
Chem. 278: 8516-8525, 2003.