국가산림자원조사와 장기생태연구 자료를 활용한 산림경관모형의 모수화 및 적용성 평가
조원희
1
⋅임원택1
⋅김은숙2
⋅임종환2
⋅고동욱3*
1)1
국민대학교 산림자원학과,2
국립산림과학원 기후변화생태연구과,3
국민대학교 산림환경시스템학과 (2020년 3월 16일 접수; 2020년 7월 15일 수정; 2020년 8월 21일 수락)Parameterization and Application of a Forest Landscape Model by Using National Forest Inventory and Long Term
Ecological Research Data
Wonhee Cho
1
, Wontaek Lim1
, Eun-Sook Kim2
, Jong-Hwan Lim2
, Dongwook W. Ko3*
1
Department of Forest Resources, Kookmin University, 77, Jeongneung-ro, Seongbuk-gu, Seoul, 02707, Republic of Korea2
Forest Ecology & Climate Change Division, National Institute of Forest Science, Seoul, Republic of Korea3
Department of Forestry, Environment, and Systems, Kookmin University, 77, Jeongneung-ro, Seongbuk-gu, Seoul, 02707, Republic of Korea(Received March 16, 2020; Revised July 15, 2020; Accepted August 21, 2020)
ABSTRACT
Forest landscape models (FLMs) can be used to investigate the complex interactions of various ecological processes and patterns, which makes them useful tools to evaluate how environmental and anthropogenic variables can influence forest ecosystems. However, due to the large spatio-temporal scales in FLMs studies, parameterization and validation can be extremely challenging when applying to new study areas. To address this issue, we focused on the parameterization and application of a spatially explicit forest landscape model, LANDIS-II, to Mt. Gyebang, South Korea, with the use of the National Forest Inventory (NFI) and long-term ecological research (LTER) site data. In this study, we present the followings for the biomass succession extension of LANDIS-II: 1) species-specific and spatial parameters estimation for the biomass succession extension of LANDIS-II, 2) calibration, and 3) application and validation for Mt. Gyebang. For the biomass succession extension, we selected 14 tree species, and parameterized ecoregion map, initial community map, species growth characteristics. We produced ecoregion map using elevation, aspect, and topographic wetness index based on digital elevation model. Initial community map was produced based on NFI and sub-alpine survey data. Tree species growth parameters, such as aboveground net primary production and maximum aboveground biomass, were estimated from PnET-II model based on species physiological factors and environmental variables. Literature data were used to estimate species physiological factors, such as FolN, SLWmax, HalfSat, growing temperature, and shade tolerance. For calibration and validation purposes, we compared species-specific aboveground biomass of model outputs and NFI and sub-alpine survey data
* Corresponding Author : Dongwook W. Ko ([email protected])
ⓒ Author(s) 2020. CC Attribution 3.0 License.
and calculated coefficient of determination (R
2
) and root mean square error (RMSE). The final model performed very well, with 0. 98 R2
and 8. 9 RMSE. This study can serve as a foundation for the use of FLMs to other applications such as comparing alternative forest management scenarios and natural disturbance effects.Key words: LANDIS-II, Gyebangsan, Species physiological parameterization, Forest landscape model, PnET-II, Forest succession, National forest inventory, Long term ecological research
I. 서 론
우리나라의 산림은 1970년대에 적극적인 녹화 사 업을 통해 임목축적량이 크게 성장하였으며 2014년에 는 물질생산, 기후조절, 수자원 함양 등 총 174조원에 달하는 거대한 공익적 가치를 지니게 되었다(Korea Forest Service, 2018). 그러나 오늘날의 산림은 노령 화와 기후변화 및 다양한 교란 등 과거에 경험하지 못한 변화를 맞이하고 있어 산림경관의 보존을 위한 대책이 필요하다(Ko et al., 2015; Tak et al., 2007).
산림경관은 몇 개의 유역, 수십만 ha 이상을 포함하 는 공간규모의 지형, 기후, 식생 등의 요소로 구성되어 있으며, 비슷한 유형의 산림생태계가 반복적으로 나타 나는 곳을 의미한다(Forman and Godron, 1981; Turner et al., 2001). 산림경관을 구성하는 각 요소들은 시공 간적으로 끊임없이 변화하는 특징을 지니고 있고 산림 경관내 식생은 짧게는 수십년에서 길게는 수세기 동안 여러 시공간적 스케일에서 발생하는 생장과 경쟁 그리 고 쇠퇴를 반복하며 다양한 변화양상을 나타내는 동적 특성을 보인다. 특히 산림천이는 식생의 생장과 경쟁 에 의해 발생하고 산림경관을 변화시키는 주요 원인으 로 꼽히며 식생의 생리적 특성과 지형적 특성, 기후적 특성에 따라 다양한 양상을 보인다(Aber and Melillo, 1991). 따라서, 산림천이에 의한 경관변화 연구는 기 존의 적지적수와 같은 개념의 연구로 동적인 특성을 모두 반영하는데 한계가 있다.
우리나라의 산림경관 변화에 대한 연구는 주로 기 후변화에 의한 침엽수, 아고산대 수종에 대한 서식지 취약성 연구를 수행하였으며 기후와 수종별 생리특성 이 함께 고려된 연구는 미비한 실정이다(Lee et al., 2007; Kong et al., 2012). 따라서, 수종별 생리적 특성 과 함께 시공간적 상호작용을 함께 고려할 수 있는 산림경관 모형을 통한 접근 방식이 필요하다(Baker and Mladenoff, 1999; Ko et al., 2015).
초기 산림경관 모형은 1970년대 JABOWA와 같은
갭모형(Gap model)으로부터 시작되었으며, 교란과 고 사 등으로 생긴 ‘숲틈’에서 수목의 정착, 생장, 고사 등의 생태학적 특성을 반영한 경관변화를 모사할 수 있다(Botkin et al., 1972). 1990년대 이후 산림경관 모형은 전산기술의 발달로 모사 공간이 제한적이던 갭 모형의 한계를 넘어 다양한 공간적 특성을 함께 모사 할 수 있는 모형이 개발되기 시작하였으며, 기후특성 과 식생구조 등의 복잡한 생태계 구조를 고려할 수 있도록 발전되었다(Mladenoff, 2004).
특히 LANDIS-II (LANdscape DISturbance and Succession) 모형은 경관규모에서 시공간적 맥락을 함 께 고려한 확률론적 모형으로 수백 ha에서 수십만 ha 의 공간과 수십년에서 수세기에 이르는 시간적 규모의 산림생태계 변화를 추정할 수 있는 모형이다(Bruijn et al., 2014; Scheller et al., 2007). LANDIS-II는 기존 의 갭모형의 임분동태를 경관규모로 확장시키기 위해 개발되었으며, 산림생태계 변화를 모사하기 위한 수목 의 생장과 경쟁, 교란, 산림관리, 토지이용 변화 등을 다양한 익스텐션(extension)으로 구성하여 연구대상 지의 생태적 생리적 특성에 초점을 맞춘 연구에 적합 한 특징이 있다. 초기 LANDIS-II를 활용한 연구는 산 불, 풍해 등 교란에 의해 생성된 경관규모의 ‘숲틈’에 서의 갱신이 주로 진행되었나, 최근에는 기후변화에 따른 우점수종의 변화와 취약수종의 평가, 산림관리에 따른 경관의 변화, 산림천이에 따른 야생동물 서식지 적합성 변화, 식생의 생장에 따른 생태계서비스 평가 등 다양한 방면으로 영역이 확장되고 있다(Scheller et al., 2007; Haga et al., 2018; United States Department of Agriculture, 2014; Thompson et al., 2016).
따라서, LANDIS-II는 성숙림이 크게 증가한 우리 나라의 산림이 기후변화, 교란, 천이 등과 같은 자연적 영향과 숲가꾸기 등 인위적 영향에 대해 어떤 변화를 겪을지 파악하는데 유용할 것으로 판단된다. 이에 이 연구에서는 산림경관 모형 LANDIS-II를 활용한 우리 나라의 산림경관 생태계 변화 연구를 수행할 수 있는
LANDIS-II 모형에 필요한 입력자료와 수종별 생리특 성 모수를 추정 및 보정하는 방안을 제시하였다. 이와 함께 추정한 입력자료와 모수를 모형에 적용한 후 평 가함으로써 해당 모형의 우리나라 산림에 대한 적용성 을 평가하고자 하였다. 이를 위해 1) 모형의 입력자료 를 제작하고 2) 다양한 문헌조사를 통해 수종별 생활 사와 생리 특성에 대한 모수 추정 방안을 제시하고 3) 국가산림자원조사(National Forest Inventory, NFI) 자료와 장기생태조사자료, 그리고 국립산림과학원에 서 실시한 아고산대 조사자료를 기반으로 모형의 보정 과 검증 방안을 제시하였다.
II. 재료 및 방법
2.1. 연구대상지
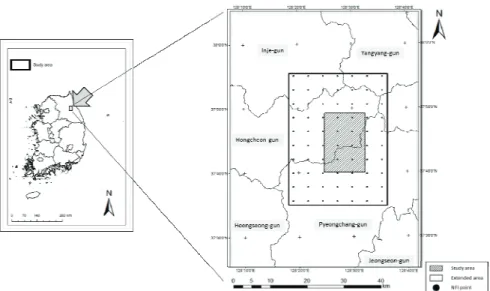
계방산(1,577m)은 강원도 홍천군 내면과 평창군 용 평면 사이에 위치하고 있으며 북위 37°42′15″에서 37°46′15″, 동경 128°26′15″에서 128°30′00″에 걸쳐 있다(Fig. 1). 계방산은 북동쪽으로 오대산(1,563m)과 서쪽에 운두령(1,089m)등 주변의 해발고도 1,000m이 상의 봉우리가 지속되어 접근성이 용이하지 않아 산림 식생 보존이 잘 되어 있고, 해발고도 960m에 있는 1ha 크기의 장기생태조사지와 1,200m 이상에 분포하는 32 개소의 아고산대 조사구에서 실시되는 식생변화에 대 한 지속적인 모니터링 자료를 보유하고 있어 모형의 적용성을 평가하기에 적절할 것으로 판단된다(Cheon
et al., 2014; National Institute of Forest Science, 2015). 이 연구에서는 계방산 중심을 기준으로 11km x 17km 지역을 연구대상지로 설정하여 산림경관 모형 에 적용하였다. 연구대상지의 고도 분포는 568m ~ 1,577m로 1,000m이상의 차이를 보이며 최대경사 45.4°, 평균경사 21.3°로 경사가 급한편이다. 계방산이 위치한 홍천지역의 30년 평년(1981-2010) 연평균기온 은 10.3°C이며, 연평균 강수량은 1,405.4mm이다.
연구대상지의 주요 우점수종은 신갈나무, 박달나무, 소나무, 피나무, 음나무, 고로쇠나무, 난티나무, 물푸 레나무, 다릅나무, 전나무, 층층나무, 잣나무 등이며 (Cheon, 2014), 1,200m이상 아고산지대에는 주목, 전 나무, 사스래나무, 분비나무, 가문비나무 등의 수종이 분포한다(Kim et al., 2014; Korea Forest Research Institution, 2015).
2.2. 모형의 소개
2.2.1. LANDIS-II Biomass-succession 익스텐션 모형
산림경관모형 LANDIS-II는 경관규모에서 수종의 생장, 경쟁, 종자 확산, 정착, 교란 등 산림 경관 변화의 다양한 프로세스를 공간명시적으로 모사하기위해 개 발되었다(He and Mladenoff 1999; Mladenoff 2004;
Scheller et al., 2007). LANDIS-II는 오픈소스형 모형 으로 생태계의 다양한 프로세스를 익스텐션으로 만들
Fig. 1. The location of the study area (grey box). The solid line is extended area for selecting additional NFI points to produce initial-community.
어 연구목적에 맞도록 활용할 수 있는 장점을 지니고 있다. 모형은 기본적으로 산림천이를 모사할 수 있는 천이 익스텐션과 산불, 풍해, 병충해, 산림관리 등을 모사할 수 있는 교란 익스텐션으로 구분되며, 모형의 공간입력자료로는 산림 변화에 영향을 미치는 환경인 자가 동일한 영역을 상정한 생태역(ecoregion) 지도와 수종과 영급 구성의 공간적 분포 지도가 있다.
이 연구의 산림경관변화 모사를 위한 천이모사의 중심에는 LANDIS-II의 Biomass-succession 익스텐 션이 있다. Biomass-succession 익스텐션은 시계열에 따른 각 생태역별 수종과 영급의 생물량의 변화를 추 정하고, 이에 따라 해당연도의 생장, 경쟁, 고사, 정착 등의 산림 생태계 변화를 모사한다. Biomass-succession 익스텐션에서 요구하는 모수는 생장과 밀접한 관련이 있는 수종별, 생태역별 연간최대순생장량(Aboveground Net Primary Productivity, ANPPmax)과 최대생물량 (Maximum biomass, Maxbiomass) 모수와 식생의 적 응과 고사와 밀접한 관련이 있는 생활사 특성인 내음 성(Shade tolerance), 최대수명(longevity) 모수가 있다 (Scheller and Mladenoff, 2004).
Biomass-succession 익스텐션에서 요구하는 모수 는 문헌자료와 경험적 자료에 기반하거나 수종별 생리특 성에 기반한 모형을 통해 추정할 수 있다. 이 연구에서는 ANPPmax와 Maxbiomass는 수종별 생리특성에 기반하 는 PnET-II 모형(Photosynthesis and Evapotranspiration
model)과 LANDIS-II PnET-succession 익스텐션을 통해 추정하였으며, 그 외 수종별 생활사 특성은 문헌 자료에 기반하여 추정하였다.
2.2.2. PnET-II 모형과 PnET-succession 익스텐션
산림생태계내에서 동일한 수종이라도 기후와 지리 적 특성에 따라 생장량은 매우 다양한 양상을 보인다.
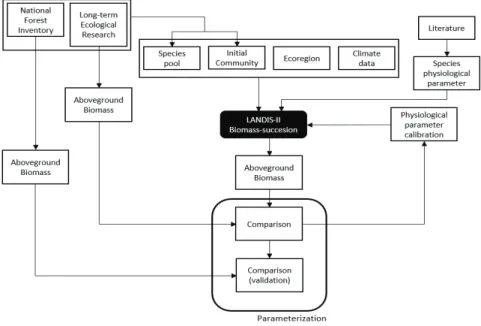
이 연구에서 LANDIS-II Biomass-succession 익스텐 션의 ANPPmax 모수 추정에 사용된 PnET-II모형은 기후와 수종별 생리특성에 따른 연간최대순생장량을 추정하는 모형으로 동일한 수종이라도 기후와 지리적 특성에 따라 차이를 보이도록 설계되었으며, 환경특성 을 반영하는 기후 자료와 광합성과 관련된 수종병 생리 특성 자료에 의해 시계열별 최대 순 생장량인 ANPPmax를 추정한다(Aber et al., 1995). 따라서, PnET-II 모형을 활용해 ANPPmax를 추정하기 위해 이 연구에서는 연 구대상지의 환경특성별 기후자료와 광합성과 관련된 수종별 생리특성 자료를 추정하여 적용하였다(Fig. 2).
Maxbiomass 추정에 사용된 PnET-succession 익스 텐션은 PnET-II 모형을 직접적으로 LANDIS-II 모형 에 적용할 수 있도록 개발되어 PnET-II 모형과 동일한 모수를 필요로한다. 또한 최대 생장 생물량에 제한을 둔 Biomass-succession과 달리 빛과 토양 수분량에 의 한 경쟁을 중심으로 한 천이과정 모사하기 때문에 생
Fig. 2. The study flow for LANDIS-II Biomss-succession parameterization.
장에 제한이 없는 특징이 있다(Bruijn et al., 2014).
그러나 Biomass-succession에 비해 상대적으로 복잡 한 프로세스를 지니고 있어 막대한 연산 시간이 소요 되는 특징이 있다. 따라서, 이 연구에서는 생장에 제한 이 없는 PnET-succession 익스텐션을 활용해 수종별 Maxbiomass를 추정하고 이를 Biomass-succession에 적용하였다.
2.3. 모형 입력자료
2.3.1. 생태역(Ecoregion)
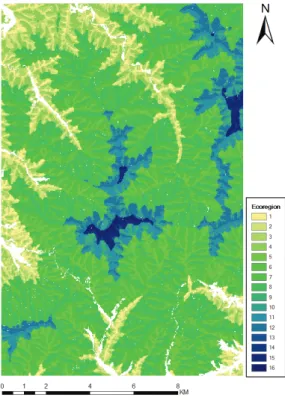
생태역(ecoregion)은 LANDIS-II에서 연구대상지 의 환경특성을 반영하는 공간입력자료이며, 산림의 변 화에 영향을 미치는 기후, 토양, 지형 등의 인자가 비 교적 동질적인 지역을 구분하여 사용한다(He and Mladenoff, 1999; Scheller et al., 2007). 이 연구에서 는 30m 해상도의 수치표고모델(Digital elevation model, DEM)을 활용하여 추정한 고도, 사면, 토양습 윤지수(Topographic Wetness Index, TWI)를 사용하 여 생태역을 분류하였다(Beven and Kirkby, 1979).
2.3.2. 대상 수종 선정
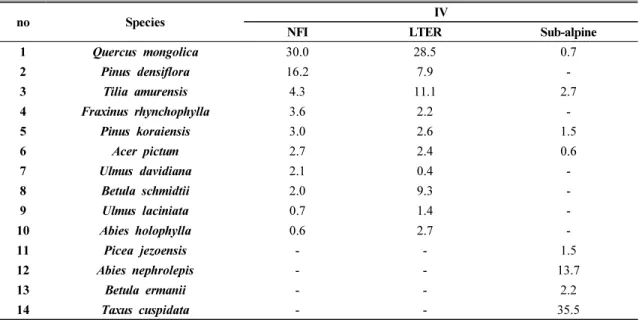
모형에 사용되는 수종은 연구대상지의 주요 우점수 종으로 구성하였다. 우점수종 선정에는 NFI, 계방산 장기생태조사자료, 그리고 국립산림과학원의 계방산 아고산대 조사자료를 활용하였고, 각 표본점의 수종별 흉고단면적으로 계산한 상대피도와 수종별 본수로 계 산한 상대밀도의 평균으로 추정된 중요도가 1.0 이상 인 수종을 선정하였다. NFI를 활용한 수종별 중요도는 계방산을 포함하는 행정구역인 홍천군, 양양군, 인제 군, 평창군, 횡성군을 모두 포함하여 204개의 표본점 으로부터 추정하였다. 장기생태조사지의 중요도는 1997년부터 2012년까지 5년 간격으로 4차례 조사한 자료를 사용하였다(National Institute of Forest Science, 2014). 1,200m이상의 아고산대의 경우 NFI나 장기생 태조사자료가 없으므로 2015년도에 조사된 자료를 이 용하여 중요도를 추정하였다(National Institute of Forest Science, 2015).
2.3.3. 초기 식생형 지도(initial community map) 초기 식생형 지도는 모형에 입력되는 초기 산림 수 종과 영급의 공간 분포 자료이다. 이 연구에서는 NFI (고도 <1200m)와 아고산대 조사자료(고도 ≥1200m)
의 수종과 흉고직경 정보를 활용하여 초기 식생형 지 도를 제작하였다. 연구지에 포함된 NFI 표본점이 적어 식생형의 대표성 확보를 위해 연구대상지에서 범위를 확장시켜 70개의 NFI 표본점을 사용하였으며 수종별 영급은 문헌상 연년생장량에 기반하여 추정하였다 (Lee et al., 2011; Fig. 1). 최종적으로 표본점의 위치 에 따라 해당되는 생태역을 확인하고 이들이 각 생태 역을 대표하는 식생형이라는 가정 하에 해당 식생형을 생태역 전역에 임의적으로 분포시키는 방법으로 초기 식생형 지도를 작성하였다.
2.3.4. 기후자료
기후자료는 모형에서 수종별 ANPPmax에 영향을 주는 요인으로 월 평균 최고 및 최저 기온, PAR, CO
2
농도, 월간 누적 강수량이 요구된다. 연구대상지의 기 후자료 중 기온, 강수량, 일사량은 계방산과 가장 가까 운 인제군 기상관측점(1973-2015년)에서 측정된 자료 로부터 고도와 사면의 영향을 반영하여 Mountain microclimate simulator (MT-CLIM) 모형(United States Department of Agriculture, 1989)을 통해 추정하였다.
PAR은 추정된 일사량을 기반으로 변환계수(Udo et al., 1999)를 적용하여 추정하였고, CO
2
농도는 National Oceanic and Atmospheric Administration’s Earth System Research Laboratory (NOAA-ESRL)에서 제공하는 하와이 마우나 로아 관측소의 대표값을 사용하였다.2.4. 수종별 생활사 및 생리특성 자료를 활용한 LANDIS-II Biomass-succession 익스텐션 모수화 PnET-II 모형과 PnET-succession 익스텐션에 사용 된 수종별 생리특성모수는 문헌과 모형 결과를 종합하 여 추정하였으며, 시간에 따른 수종별 지상부생물량 모형결과를 장기생태조사자료 기반의 지상부생물량 과 비교하여 보정하였다. 모수 추정은 한반도를 포함 한 중국, 일본 등을 연구대상지로 하는 문헌을 활용하 고, 문헌이 없으면 LANDIS-II 모수화 가이드라인 (Gustafson et al., 2015)을 반영하여 초기값을 설정하 였다. 주요 모수로는 엽질소함량비(FolN), 최상위 수 관층의 단위면적당 잎의 질량(SLWmax), 광포화도 (HalfSat), 수종별 상대적 내음성(5단계), 수종별 생장 시작 온도(Tmin)과 생장 최적 온도(Topt), 최대수명 (longevity) 등이 있다.
Scie nt ific na m e Pn ET -II pa ra m et ers FolN (%) SLW max (g/m 2 ) Halfsat (W/ m 2 )T m in ( o C) b) Topt ( o C) b) Shad e tolerance c ) Lon gevity Q. m on golica 2.3 (1.2- 4.6) a ) 80 (26- 104) a ) 388 3.7 21 3 300 P. d ens ifl or a 1.8 (0.8- 1.8) a ) 263 (89-2 63) a ) 600 2.9 21 1 250 T. am ur ens is 2.0 ( - ) d ) 73 (28-8 1) a ) 370 3.0 21 4 325 F. rhync ho ph ylla 1.8 (1.8- 2.8) a ) 80 (59-9 4) a ) 388 0.9 20 3 300 P. ko raie nsis 1.5 (0.8- 1.5) a ) 232 (11 9-23 3) a ) 385 1.0 20 3 325 A. p ict um 1.8 (1.7- 3.0) a ) 65 (30-6 7) a ) 300 2.2 21 5 350 U. davidia na 1.8 ( - ) d ) 84 (33-9 6) a ) 425 3.0 21 3 300 B. sc hm idtii 2.4 ( - ) d ) 90 (27-9 2) a ) 500 1.0 18 2 275 U . laci ni ata 2.0 ( - ) d ) 71 (33-9 6) a ) 370 2.8 19 4 325 A. h olop hylla 1.8 ( - ) d ) 290 ( - ) d ) 425 0.5 16 3 300 P. jezoe nsis 2.0 ( - ) d ) 120 (18 5-21 9) a ) 350 0.6 16 4 325 A. n ep hrole pis 1.8 ( - ) d ) 145 (13 6-14 6) a ) 275 0.5 16 5 350 B. er mani i 2.1 ( - ) d ) 92 (27-9 2) a ) 500 0.1 21 2 275 T. c us pidat a 1.0 (1.1- 2.2) a ) 145 (64-1 45) a ) 250 0.3 17 5 500 a) M itchell, 199 8; Kitao et al ., 200 3; 200 5; Son et al ., 200 1; Ichie et al ., 2002; Ha n et al ., 20 03, Lim et al ., 20 08; Ch o et al ., 20 08; Yan et al ., 20 12; Qi et al ., 20 13; Li u et al ., 20 13; b) Yim , 1977 , c) Shao et al ., 19 94; Jian g et al ., 19 99; X iadon g et al ., 2005; W an g et al ., 201 0; Wang et al. , 2019.
Tab le 1 . The p hysi ol og ica l pa ramete rs of e ach spe cie s
FolN은 수목의 생산성과 양의 상관관계를 지니며, 특히 ANPPmax와 Maxbiomass 증감이 매우 민감하게 반응하는 모수이다. FolN은 문헌상 1.0 - 4.6%까지 다양한 분포를 보였으며, 수종별 생장속도와 생물량 분포에 맞도록 1.0 - 2.4%의 분포를 보이도록 조정하 였다. SLWmax도 FolN과 마찬가지로 수목의 생산성 과 양의 상관관계를 보이는데, 주로 침엽수가 활엽수 에 비해 높은 특징을 보이고 있다(Mitchell and Hinckley, 1993; Reich et al., 1997). SLWmax는 문헌상 최소 65g/m
2
에서 290g/m2
의 분포를 보이며 조사자료의 수 종별 생물량과 비슷한 생장량을 보이도록 미세조정에 활용하였다.Halfsat은 내음성이 높은 수종일수록 낮으며 내음성 이 낮은 수종은 높은 경향을 보인다(Lee, 2014). 따라 서, 문헌자료를 통해 추정한 내음성에 따라 양수는 400 - 600W/m
2
, 중성수는 300 - 500W/m2
, 음수 200 - 400W/m2
를 적용하도록 조정하였다. 수종별 수명 (longevity)은 수종별 쇠퇴(고사)와 관련이 있는 모수 로 이 연구에서는 내음성에 의한 천이양상을 모사할 수 있도록 조정하였다. 생장온도는 Yim (1977)에서 제시한 온량지수(Warmth Index)를 기반으로Tmin은 0°C - 4°C의 분포하도록 조정하고, 최적생장기온인 Topt는 16°C에서 24°C사이로 조정하였다(Table 1).앞서 제시한 생활사 및 생리특성 입력자료를 PnET-II 모형과 PnET-succession 익스텐션에 구동하여 얻은 ANPPmax와 Maxbiomass로 모형을 구동하고 1997, 2002, 2007, 2012년의 모형 결과 중 수종별 지상부생 물량과 Lim (1998)에 의해 추정된 계방산 장기생태조 사자료의 같은 시기 지상부생물량 변화를 비교하였으 며, 만족스러운 결정계수(R
2
)와 최소 평균 제곱근 오차 (Root Mean Square Error; RMSE)가 도출되도록 전체 생리특성 입력자료의 조합을 적절한 범위에서 교차 조 정하며 보정하였다.계방산 장기생태조사는 1ha의 조사지에 걸쳐 10 x 10m 크기의 조사구 100개에서 1997년부터 5년 간격 으로 지속적으로 식생 조사가 이루어지고 있다. 보정 을 위한 모형의 구동은 1997년부터 2012년까지 실시 하였고, 장기생태조사지와 동일한 크기의 공간과 식 생형을 적용하였으며, 조사지가 위치한 해발고도 900 - 960m 남사면에 해당하는 생태역과 기후자료를 활용 하였다. 보정을 실시한 수종은 전체 대상 수종 중 장기 생태조사지에 분포하는 10개 수종으로 선정하였다.
2.5. 모형 검증 및 적용성 평가
보정의 결과로 확보한 모수를 전체연구대상지에 적 용한 결과의 검증은 독립적인 자료(NFI, 아고산대 조 사자료)를 활용하여 해발고도 1,200m를 기준으로 각 각 비교하였다. 1,200m 미만지역은 NFI 자료에서 추 정한 수종별 지상부생물량을 사용하고, 1,200m 이상 지역은 아고산대 조사자료를 활용하여 추정한 전체 지 상부생물량의 비교를 실시하였으며 보정과 동일한 방 법으로 진행하였다. 검증에 사용된 수종은 보정에서 사용된 10개 수종과 아고산대 수종 4개를 포함하여 총 14개 수종에 대해 실시하였다.
III. 결 과
3.1. 모형 입력자료 구축
3.1.1. 생태역 구분
생태역 구분 요소 중 해발고도는 500 - 800m, 800 - 1,200m, 1,200 - 1,400m, 1,400m 초과의 4개로 구분 하였고, 사면은 남사면과 북사면, TWI는 연구대상지 전체의 평균을 기준으로 습윤지역과 건조지역으로 분
Fig. 3. The ecoregions of the study area.
류한 후 종합하여 총 16개의 생태역으로 구분하여 적 용하였다(Fig. 3, Table 2). 고도별 면적 비율은 800m이 상 1,200m미만 지역이 66%이상으로 가장 넓었고, 1,200m이상 아고산대는 12.8%였다. 사면별 면적 비 율은 남사면이 49%, 북사면이 51%로 비슷하였고, 토 양습윤정도는 건조지역이 60%이상으로 습윤지역보다
넓었다.
3.1.2. 대상 수종 선정 및 초기식생형 지도 제작 대상 수종 선정을 위해 실시한 중요도 분석은 NFI 와 장기생태조사자료에서 신갈나무, 소나무, 피나무 등이 높은 중요도를 보였고, 아고산대에서는 주목, 분비
Ecoregion Elevation (m) TWI Aspect area (%) No. community*
1
500 to 800
Wet South 4.8 5
2 North 5.7 3
3 Dry South 4.6 7
4 North 5.5 9
5
800 to 1,200
Wet South 11.2 5
6 North 11.3 9
7 Dry South 22.2 8
8 North 21.9 11
9
1,200 to 1,400
Wet South 1.4 4
10 North 1.5 8
11 Dry South 4.2 -
12 North 4.3 2
13
> 1,400
Wet South 0.1 1
14 North 0.1 2
15 Dry South 0.6 5
16 North 0.6 10
Table 2. Description of each ecoregion in the study area
no Species IV
NFI LTER Sub-alpine
1 Quercus mongolica 30.0 28.5 0.7
2 Pinus densiflora 16.2 7.9 -
3 Tilia amurensis 4.3 11.1 2.7
4 Fraxinus rhynchophylla 3.6 2.2 -
5 Pinus koraiensis 3.0 2.6 1.5
6 Acer pictum 2.7 2.4 0.6
7 Ulmus davidiana 2.1 0.4 -
8 Betula schmidtii 2.0 9.3 -
9 Ulmus laciniata 0.7 1.4 -
10 Abies holophylla 0.6 2.7 -
11 Picea jezoensis - - 1.5
12 Abies nephrolepis - - 13.7
13 Betula ermanii - - 2.2
14 Taxus cuspidata - - 35.5
Table 3. The importance value of tree species used in the study
나무, 사스래나무, 가문비나무 등의 중요도가 높게 나 왔다(Table 3). 따라서, 실시한 중요도분석을 바탕으로 1.0이상의 중요도를 보이는 14개의 수종을 선정하였다.
1,200m 미만 고도의 초기 식생형 선정을 위해 확장 된 연구지에 포함되는 NFI 표본점은 총 70개였으나, 연구지에 부합하는 생태역에 포함되는 57개의 표본점 을 사용하였고 1,200m 이상의 연구지는 32개 표본점 의 아고산지대 조사 자료를 활용하여 89개의 식생형 을 구성하였다(Table 2). 다만 생태역 11번은 해당하 는 표본점이 없어 환경이 유사한 12번 생태역의 초기 식생형을 적용하였다.
3.1.3. 기후 입력자료 구축
MT-CLIM을 통해 추정한 기후자료는 고도와 사면에 따라 8개의 기후대로 구분하였다(Fig. 4). 추정된 기온과 강수량은 사면의 영향을 받지 않았으며 고도에 의한 차 이만 나타났는데, 기온은 고도가 증가함에 따라 감소하 는 경향을 보였고, 강수량은 고도의 증가에 따라 함께 증 가하였다. PAR은 고도에 의한 차이보다 사면에 의한 차 이가 컸으며, 계절적으로는 여름의 사면차는 작았고 봄, 가을, 겨울에는 남사면이 북사면에 비해 높게 나타났다.
3.2. LANDIS-II Biomass-succession 익스텐션 모수화 기후와 수종별 생리특성을 반영한 PnET-II 모형 결 과인 ANPPmax를 수종과 생태역별로 살펴보면 신갈 나무, 난티나무, 피나무 등이 다른 수종에 비해 높게 나타났고, 남사면이 비교적 높았다(Table 4). 그 중 난 티나무가 9번, 11번, 13번, 15번 생태역에서 684g/m
2
로 가장 높은 ANPPmax를 보였고, 다음으로 신갈나무 가 9번, 11번 생태역에서 667g/m2
로 나타났다. 아고산 대 수종의 경우 사스래나무가 595g/m2
로 가장 높았고, 다음으로 분비나무가 420g/m2
로 높았으며, 남사면인 13 번과 15번 생태역의 ANPPmax가 높게 나타났다.PnET-succession 익스텐션의 결과인 수종별 Maxbiomass 는 전나무가 30,393g/m
2
로 가장 높았고, 다음은 사스래 나무로 28,103g/m2
를 보였다. 난티나무(18,506g/m2
), 가문비나무(18,576g/m2
), 고로쇠나무(18,941g/m2
) 등은 20,000g/m2
이하로 추정되었다.3.3. 모형구동 및 검증
3.3.1. 수종별 지상부 생물량 모사 평가
모형 보정을 통해 확정된 모수(ANPPmax, Maxbiomass) 를 적용하여 1997년부터 2012년까지의 수종별 지상 부생물량을 모사하였다. 각 시기별 수종간의 지상부생 물량 비교는 모두 R
2
0.9 이상으로 높은 상관관계를 보였다(Fig. 5). 초기(1997년)의 수종별 지상부생물량 은 소나무, 잣나무를 제외하고 모형의 결과가 조사자 료에 비해 높게 나타났고, 2002년의 결과는 신갈나무, 소나무, 잣나무를 제외한 모형의 결과가 조사자료에 비해 높게 나타났다(Table 5). 2007년은 신갈나무, 소 나무를 제외한 모형의 결과가 조사자료에 비해 높게 나타났으며, 2012년은 신갈나무, 소나무, 박달나무를 제외한 모형의 결과가 조사자료에 비해 높게 나타났 다. 모형에서 각 시계열별 지상부생물량의 증가율은 내음성 4 이상의 피나무, 고로쇠나무, 난티나무, 잣나 무 등과 같은 음수에서 높게 나타났고, 내음성 모수 2 이하의 소나무, 박달나무와 같은 양수에서는 낮게 나타났다(Table 3). 그 결과 각 시기별 수종간의 지상 부생물량 차이로 추정한 RMSE는 초기에 4.1에서 2012년에 7.3으로 다소 증가하는 것으로 나타났다.Fig. 4. The estimated climate by MT-CLIM based on Inje weather station. a) represents the estimated maximum temperature and precipitation in each elevation band. b) represents the monthly mean PAR North and South aspect.
sp ecies ec o 01 eco 02 eco 03 eco 04 eco 05
ec o 06
eco 07 eco 08 eco 09 eco 10
ec o 11
eco 12 eco 13 eco 14 eco 15 eco 16
Ma x bi om as s Q . m on golica 63 9 616 639 61 6 65 9 63 9 659 639 667 63 6 66 7 636 664 645 66 4 645 20 564 P. de nsi flora 37 5 366 375 36 6 38 1 37 3 381 373 376 36 3 37 6 363 375 368 37 5 368 23 319 T. am ur ens is 57 3 556 573 55 6 59 1 57 6 591 576 601 57 5 60 1 575 597 584 59 7 584 22 367 F. rhy ncho ph ylla 56 5 547 565 54 7 58 7 57 1 587 571 599 57 5 59 9 575 600 584 60 0 584 22 688 P. koraie ns 40 3 395 403 39 5 40 8 40 1 408 401 410 39 1 41 0 391 402 395 40 2 395 22 185 A. pict um 55 5 541 555 54 1 57 0 55 7 570 557 577 55 7 57 7 557 576 564 57 6 564 18 941 U . david ia na 58 7 568 587 56 8 60 5 59 0 605 590 616 59 0 61 6 590 614 599 61 4 599 20 709 B. sc hm iditii 51 7 492 517 49 2 57 0 54 7 570 547 606 57 1 60 6 571 618 596 61 8 596 21 483 U. laci niata 63 2 610 632 61 0 66 5 64 5 665 645 684 65 2 68 4 652 684 665 68 4 665 18 506 A. h olo ph ylla 35 4 344 354 34 4 37 8 36 9 378 369 394 36 9 39 4 369 389 380 38 9 380 30 393 P. jezoe nsis 38 0 369 380 36 9 40 2 37 8 402 378 402 38 6 40 2 386 405 395 40 5 395 18 576 A. n ep hr olepis 37 4 362 374 36 2 40 7 37 9 407 379 412 39 5 41 2 395 420 410 42 0 410 21 345 B. erma ni i 52 8 507 528 50 7 56 5 54 7 565 547 589 56 0 58 9 560 595 576 59 5 576 28 103 T. c uspid ata 36 1 354 361 35 4 37 5 36 0 375 360 376 36 6 37 6 366 378 372 37 8 372 19 235
Tab le 4 . Es tim ated A NPP max a nd Ma xb iomass in eac h ec or eg io n. U nit : g/m 2 Sp ecies 1997 20 02 200 7 201 2 Su rv ey Mo de l Su rv ey M ode l Su rv ey Mod el Su rv ey M ode l Q. m on go lica 94.0 99. 8 105.9 99 .7 111.2 99.3 113.6 98.3 P. de nsi flora 13.3 11. 1 15 .9 11 .4 18. 5 11.6 20. 6 11.8 T. a m ure nsis 13.2 18. 6 14 .9 21 .0 16. 8 23.3 18. 4 25.3 F. rhynch ophy lla 3.7 9.0 3.9 10 .2 3.4 11.3 3.0 12.4 P. kora ie nsis 1.0 0.7 1.3 1.2 1.6 1. 6 1.8 2.0 A. pict um 4.1 8.8 4.3 10 .1 4.5 11.3 4.9 12.8 U. da vidia na 0.6 1.0 0.7 1.5 0.8 1. 9 0.8 2.7 B. sc hm iditii 27.6 33. 9 31 .6 35 .0 35. 5 35.7 39. 1 36.4 U. laci niat a 2.5 4.0 3.3 5.5 3.9 6. 7 4.5 8.1 A. h olop hylla 1.3 3.3 1.8 3.6 2.2 4. 0 2.6 4.3 RM SE 4.1 4.3 5.9 7.3
Tab le 5 . Est im at ed abo ve gro und b io m as s in LT ER s ites an d R M SE. Uni t: ton/ ha
3.3.2. 모형 검증
모형보정에 활용되지 않은 NFI 자료와 아고산대조 사자료를 대상으로 실시한 모형의 검증결과 수종별 지
상부생물량의 상관관계는 R
2
0.98로 나타났으며 보정 의 결과와 비슷한 수준을 보였다. RMSE는 8.9로 보정 의 결과와 비슷하였다 (Fig. 6; Table 6). 보정을 거친 Fig. 5. The comparison of species AGB between LTER survey data and LANDIS-II results. The R2
represents 0.992 in 1997, 0.987 in 2002, 0.981 in 2007, and 0.974 in 2012. The dash lines are drawn line in each time step. The red solid line represents 1:1 line.Fig. 6. The comparison of species AGB between survey data and LANDIS-II results. The R
2
represents 0.981. The dash line is drawn line. The red solid line represents 1:1 line.10개 수종의 검증을 실시한 1,200m 미만 지역은 느릅 나무, 물푸레나무, 전나무의 모형에서 추정한 수종별 지상부생물량이 조사자료에 비해 높게 나타났으며, 그 밖에 수종은 조사자료의 결과가 높게 나타났다. 1,200m 이상의 아고산대는 전체 지상부생물량을 비교하였으 며, 모형의 결과가 조사자료에 비해 다소 높게 나타났다.
IV. 고 찰
이 연구에서는 LANDIS-II 모형을 대상으로 산림경 관모형의 적용성 평가를 진행하였다. 이 과정에서 모 형이 요구하는 다양한 입력자료제작, 모형의 모수화, 그리고 모형의 보정 및 검증 방안을 제시하고자 하였다.
4.1. 초기 식생형 지도 기반 자료 대표성 초기식생형지도 제작에 사용된 NFI는 우리나라 전 역을 4km 간격으로 계통추출하여 조사한 자료이며 5 년마다 갱신되어 시공간적으로 보편적인 초기 식생형 을 선정하기에 적합하다. 이는 계방산일대에 뿐만 아 니라 다른 지역에 대한 적용성도 고려한 방법으로 향 후 LANDIS-II의 활용성을 높일 수 있는 선정방법으 로 판단된다. 이와 함께 연구에 사용된 아고산대조사 자료를 통해 NFI 표본점이 적은 1,200m 이상의 독특 한 생태역에 대한 모형의 활용성을 평가할 수 있었다.
그러나 NFI는 표본점 격자가 4km로 연구대상지에 대한 세밀한 식생분포의 차이를 반영하기 어렵다. 뿐 만 아니라 연구대상지 내에 포함된 표본점 수가 매우
적어 이 연구에서도 인근 지역을 포함하여 식생지도를 제작해야 했으므로 연구대상지 대표성 면에서도 한계 가 있다. 이 같은 이유로 다양한 환경조건을 반영하는 각 생태역에 존재하는 식생형을 모두 반영하기 어렵기 때문에, 30미터 해상도로 제작된 생태형과 4km 간격 을 지닌 NFI 표본점 간의 공간적 스케일의 차이에 의 한 간극을 극복하기 위한 개선이 필요하다(United States Department of Agriculture, 2014; Scheller et al., 2007; Haga et al., 2018).
아고산대조사자료는 1,200m 이상 아고산대 전체 식생 현황을 비편향적으로 나타낼 수 있는 체계적인 조사 표본점이 없다는 것이 한계이다. 그 결과로 생태 역 11번은 표본점이 없고 생태역 16번에 전체의 31%
에 해당하는 표본점이 분포한 것을 볼 수 있다(Table 2).
또한 아고산대의 혹독한 기후 조건에 의해 수목의 생 장, 경쟁, 교란 등이 저고도 지역과 비교하여 매우 다 른 양상을 보일 수 있다는 점을 감안한다면(Kim et al., 2012), 모형의 아고산대 적용을 위한 생리생태적 특성에 대한 이해와 아고산대 조사자료의 편향성을 줄 이는 개선이 동시에 필요하다.
4.2. 생리모수 적절성
이 연구에서 추정한 수종별 생리모수는 우리나라, 중국, 일본 등 동아시아 지역의 문헌을 종합하여 모수 화 함으로써 다양한 경관에 적용할 수 있도록 하였다.
그러나 주목은 자료의 한계로 동일 속(genus) 수준에 서 모수화를 시도하였고, 그 외 자료의 부족으로 추정 하지 못한 모수는 보정과정을 통해 추정하여 한계를 극복하였다.
수종별 생리모수를 통해 추정한 ANPPmax는 동일 한 수종이라도 각 생태역별 기후특성에 따라 다양하게 나타나며, 이는 환경특성에 따른 생장량 변화를 모형 과 추정된 모수의 조합으로 적절히 모사할 수 있었음 을 보여준다(Koo et al., 2001; Han et al., 2014; Table 4). 한편 모형에 적용한 기후특성 중 ANPPmax에 주 로 영향을 끼친 요인은 고도에 따른 기온의 차이로 판단된다. 모형에 적용한 기온은 고도와 음의 상관관 계를 보이며, Topt 모수가 낮은 수종일 수록 높은 고도 의 생태역에서 ANPPmax가 증가하는 것으로 나타났 다. 특히 1,200m 이상의 고도에서 아고산대 수종의 ANPPmax가 뚜렷히 증가하는 것으로 나타났고 이를 기반으로 수종별 적정 환경에 따른 천이양상 모사가 가능할 것으로 판단된다.
Species ton/ha
survey model Q. mongolica 113.8 91.9
P. densiflora 22.8 18.3 T. amurensis 38.6 24.2 F. rhynchophylla 7.6 15.7 P. koraiensis 2.6 2.4
A. pictum 15.0 12.0
U. davidiana 5.5 12.7 B. schmidtii 8.0 4.2 U. laciniata 9.3 5.5 A. holophylla 3.3 6.7 sub-alpine 194.3 196.6
RMSE 8.9
Table 6. Estimated initial aboveground biomass in the GBS study area and RMSE. Unit: ton/ha
4.3. 모형 보정 및 검증의 한계
이 연구에서는 보정(장기생태조사자료)과 검증(NFI, 아고산대 조사자료)에 독립적인 자료를 활용함으로써 다방면적인 모형의 적용성을 평가할 수 있었다. 보정 에 활용한 장기생태조사자료와 모형의 수종별 지상부 생물량의 시계열적 추이는 상당히 일치하는 결과를 보 였다. 각 시기별 0.9 이상의 높은 R
2
는 최대 우점수종 인 신갈나무의 영향이 큰 것으로 평가되었다. 한편 모 사기간이 진행되면서 장기생태조사자료의 신갈나무 지상부생물량 증가율이 모형보다 점차 높게 추정되었 는데, 이는 RMSE가 다소 증가하는 원인이 되었다 (Fig. 5; Table 5).모사 초기 모형에서 지상부생물량이 다소 과대평가 하는 경향은 선행연구와 마찬가지로 모형의 특성상 초 기 지상부생물량 추정과정에서 산불, 풍해, 병충해 등 과 같은 외부요인이 반영되지 않기 때문에 발생하는 것으로 보인다(Scheller and Mladenoff, 2004). 또한, 시계열별 지상부생물량의 증가율을 볼 때 내음성에 의 한 식생천이가 강하게 반영되는 것으로 나타났다 (Table 3). 이를 통해 내음성에 의한 식생천이를 모사 할 수 있을 것으로 판단되나, 내음성 모수와 함께 수종 별 쇠퇴(고사)에 영향을 끼치는 수명(longevity) 모수 에 대한 민감도 분석을 통해 모형의 정밀성에 대한 개선이 필요할 것으로 보인다.
NFI와 아고산대 조사자료를 활용한 검증과정의 RMSE는 보정과정에서 나타난 RMSE에 비해 다소 높 게 추정되었는데, 이는 아고산대 수종의 추가와 다양 한 생태역을 고려함과 동시에 보정과 검증에 독립된 자료를 활용한 접근이 가져올 수 있는 최소한 두 가지 한계점으로 보인다(Baker and Mladenoff, 1999;
Garzon-Lopez et al., 2014; Thuiller et al., 2003). 첫 째, 각 자료가 지닌 식생정보의 특성에 의한 차이이다.
LANDIS-II의 식생형 자료는 정해진 크기의 래스터 픽셀 하나에 존재하는 수종과 영급별 생물량으로 정의 된다. 따라서 장기생태조사자료의 정밀성과 국가산림 자원조사자료의 포괄성이라는 특성 차이로 인해 이로 부터 제작된 모형식생자료의 성격이 달라짐에 따라 결 과에 영향을 미칠 수 있다.
둘째, 장기생태조사자료와 국가산림자원조사의 공 간적 대표성 문제이다. 전자의 경우 산림경관스케일에 서 환경조건이 유사한 작은 규모의 고정조사구에서 조 사된 결과이나, 후자는 매우 넓은 지역의 다양한 환경 조건에서 나타나는 식생의 특성을 반영한다는 점에서
공간적 스케일의 불일치가 나타난다. 이 연구에서 사 용된 모형의 특성을 감안하면 전자보다는 다양한 환경 에서의 식생 조사자료를 요구하며, 후자보다는 보다 자세한 환경 조건의 차이를 반영한 식생 조사자료가 필요하다. 따라서 이러한 공간적 특성을 반영할 수 있 도록 다양한 환경 변수에 의한 식생의 차이가 고려된 자료를 활용한다면 산림경관 모형 연구의 개선에 큰 도움이 될 것으로 기대된다.
V. 결 론
오늘날 우리의 산림은 과거 치산녹화 사업시기에 비해 다양한 층위와 다양한 수종구성을 지닌 복잡한 구조의 산림으로 탈바꿈하였고 기후변화, 산불, 풍해, 인위적 교란 등 그동안 경험하지 못한 위험에 노출되 어 있다. 특히, 아고산대 수종의 경우 기후변화에 매우 취약할 것으로 많은 연구를 통해 언급되고 있어 예상 치 못한 산림경관의 변화를 맞이할 것으로 보인다 (Chung et al., 2015; Koo et al., 2016; Lambrecht et al., 2006; Woodward et al., 1995). 따라서 오늘날 복 잡한 산림생태계구조와 기능의 인과관계를 이해하기 위해서 모형을 통한 연구는 필수적이라고 할 수 있다.
이에 이 연구에서는 수종별 생장과 경쟁 그리고 다양 한 교란과 기후의 영향 등을 나타낼 수 있는 산림경관 모형 LANDIS-II를 활용하여 산림경관의 복잡한 구조 의 변화를 모사하기위한 기반을 마련하고자 하였다.
이 연구에서는 LANDIS-II모형을 소개하고 모형에 서 요구하는 환경특성, 식생형, 기후자료 등의 기본적 인 입력자료 제작방법과 다양한 문헌조사를 통한 수종 별 생리특성모수의 추정 및 모형의 전반적인 모수화 과정을 제시하였다. 그리고 이를 실질적으로 모형에 적용하여 모형과 조사자료의 비교를 통해 모형 적용의 가능성을 살펴보았다. 그러나 모형의 보정과 검증과정 에서 조사자료의 한계는 앞으로 개선되어야 할 부분이 다. 특히, 다양한 환경특성과 그에 따른 수종별 생장특 성을 나타내는 조사자료의 확보와 지속적인 모니터링 을 통해 모형의 신뢰도와 안정성을 향상시킬 수 있을 것으로 보인다.
이 연구에서 수행한 적용성 평가는 계방산일대에 분포하는 우점수종을 대상으로 진행하였고 모형에 적 합한 각 수종별 모수를 확보하였다. 이 결과를 기반으 로 계방산일대에 대한 식생천이 모사가 가능할 것으로 보이며, LANDIS-II에서 제공하는 산불, 풍해, 태풍,
병충해 등 다양한 익스텐션을 활용하여 기후변화와 교 란 등의 시나리오기반의 식생천이 모사, 식생천이에 의한 야생동물 서식지 적합성의 연구 등 다방면으로 확장도 기대할 수 있을 것으로 판단된다.
적 요
산림경관모형은 산림생태계의 복잡한 구조와 다양 한 기능의 동적특성을 연구하는데 적합한 모형으로 평 가받는다. 산림경관모형은 경관생태학을 기반으로 제 작되었으며, 그 특성상 넓은 시공간적 규모를 다루기 때문에 새로운 지역에 적용하는데 환경특성, 수종특성 등에 대한 모수화와 검증에 어려움이 있다. 이에 이 연구에서는 산림경관모형 LANDIS-II Biomass succession 익스텐션에 대한 국내 적용성을 평가하기 위해 계방산 일대를 대상으로 1) 공간정보 입력자료 제작 및 수종 특성 모수화, 2) 모형의 보정, 3) 모형의 적용 및 검증 방안을 제시하였다. 모형에 적용한 총 14수종은 국가 산림조사(National Forest Inventory; NFI), 장기생태 조사자료, 아고산대조사자료 기반의 수종별 중요도를 기반으로 선정하였으며, 공간정보 입력자료는 30m 해 상도의 수치표고모형을 기반으로 제작한 생태역 지도 와 NFI와 장기생태조사자료 기반의 초기 식생형 지도 등을 제작하였다. 수종별 생장모수(ANPPmax, Maxbiomass)는 한국, 중국, 일본 등 동아시아 지역의 생리실험 문헌자료를 종합하여 선정한 수종별 생리특 성 모수(FolN, SLWmax, Halfsat, 생장온도, 내음성 등)를 PnET-II 모형에 적용하여 추정하였다. 모형의 보정과 검증은 모형과 조사자료의 수종별 지상부생물 량을 비교하여 산출한 결정계수(R
2
)와 최소 제곱근 오 차(RMSE)를 통해 실시하였으며, 검증결과 0.98의 R2
와 8.9의 RMSE의 준수한 결과를 나타냈다. 따라서, 이 연구를 기반으로 한반도의 산림경관 변화를 모사할 수 있을 것으로 판단되며 산림관리, 산불, 풍해, 병충 해, 기후변화 등 외적요인에 따른 산림경관 변화에 대 한 연구가 수행될 수 있을 것으로 기대된다.감사의 글
본 연구는 과학기술정보통신부(한국연구재단) ‘중 견연구자지원사업(NRF-2017R1A2B4010460)’, 산림 청(한국임업진흥원) ‘산림과학기술연구개발사업(2019 150B10-2023-0301)’, 국립산림과학원 일반연구과제
‘산림생태계서비스 증진을 위한 생태적 산림관리기술 개발’의 지원에 의하여 이루어졌습니다.
REFERENCES
Aber, J. D., and J. M. Melillo, 1991: Terrestrial Ecosystems. Academic Press, Massachusetts, USA, 592pp.
Aber, J. D., S. V . Ollinger, C. A. Federer, P. B. Reich, M. L. Goulden, D. W. Kicklighter, J. M. Melillo, and R. G. Lathrop, 1995: Predicting the effects of climate change on water yield and forest production in the Northeastern United States. Climate Research 5(3), 207-22.
Baker, W. L., and D. Mladenoff, 1999: Spatial Modeling of Forest Landscape Change. Cambridge University Press, USA, 352pp.
Beven, K. J., and M. J. Kirkby, 1979: A physically based, variable contributing area model of basin hydrology. Hydrological Sciences Bulletin 24(1), 43-69.
Botkin, D. B., Janak, J. F., and J. R. Waalis, 2009:
Some ecological consequences of a computer model of forest growth. British Ecological Society 60(3), 849-872.
Bruijn, A. D., E. J. Gustafson, B. R. Sturtevant, J.
R. Foster, B. R. Miranda, N. I. Lichti, and D. F.
Jacobs, 2014: Toward more robust projections of forest landscape dynamics under novel environmental conditions: Embedding PnET within LANDIS-II.
Ecological Modelling 287, 44-57.
Cheon, K. I., 2014: A study on the spatial distribution characteristics and time serial changes of forest vegetation at Mt. Gyebang. Ph.D. Thesis. Kyungpook National University, Daegu, 134pp. (in Korean with English abstract)
Cheon, K. I., J. H. Chun, H. M. Yang, J. H. Lim, and J. H. Shin, 2014: Changes of understory vegetation structure for 10 years in long-term ecological research site at Mt. Gyebang. Journal of Korean Forest Society 103(1), 1-11. (in Korean with English abstract)
Cho, M. S., K. W. Kwon, G. N. Kim, and S. Y.
Woo, 2008: Chlorophyll contents and growth performances of the five deciduous hardwood species growing under different shade treatments.
Korean Journal of Agricultural and Forest Meteorology 10(4), 149-157. (in Korean with English abstract)
Chung, J., H. Kim, S. Lee, K. Lee, M. Kim, and Y.
Chun, 2015: Correlation analysis and growth prediction between climatic elements and radial growth for Pinus koraiensis. Korean Journal of Agricultural and Forest Meteorology 17(2), 85-92. (in Korean with English abstract)
Forman, R. T. T., and M. Godron, 1981: Patches and structural componets for a landscape ecology.
BioScience 31(10), 733-740.
Graumlich, L. J., 1991: Linked References Are Available on JSTOR for This Article : SUBALPINE TREE GROWTH, CLIMATE, AND INCREASING CO
2
: AN ASSESSMENT OF RECENT GROWTH TRENDS. Ecology 72(1), 1-11.Gustafson, E. J., A. M. G. De Bruijn, and B. R.
Miranda, 2015: PnET-Succession v1.0.1 Extension User Guide. LANDIS-II Foundation. Reproduction 0-9.
Haga, C., T. Inoue, W. Hotta, R. Shibata, S.
Hashimoto, H. Kurokawa, T. Machimura, T. Matsui, J. Morimoto, and H. Shibata, 2018: Simulation of natural capital and ecosystem services in a watershed in Northern Japan focusing on the future underuse of nature: By linking forest landscape model and social scenarios. Sustainability Science 14(1), 89- 106.
Han, Q., T. Kawasaki, S. Katahata, Y. Mukai, and Y. Chiba, 2003: Horizontal and vertical variations in photosynthetic capacity in a Pinus densiflora crown in relation to leaf nitrogen allocation and acclimation to irradiance. Tree Physiology 23(12), 851-857.
Han, S., J. An, T. K. Yoon, S. J. Yun, J. Hwang, M. S. Cho, and Y. Son, 2014: Species-specific growth responses of Betula costata, Fraxinus rhynchophylla, and Quercus variabilis seedlings to open-field artificial warming. Korean Journal of Agricultural and Forest Meteorology 16(3), 219- 226. (in Korean with English abstract)
He, H. S., and D. J. Mladenoff, 1999: The effects of seed dispersal on the simulation of long-term forest landscape change. Ecosystems 2(4), 308-319.
Ichie, T., Y. Kitahashi, S. Matsuki, Y. Maruyama, and T. Koike, 2002: The use of a portable non- destructive type nitrogen meter for leaves of woody plants in field studies. Photosynthetica 40(2), 289- 292.
Jiang, H., C. Peng, M. J. Apps, Y. Zhang, P. M.
Woodard, and Z. Wang, 1999: Modelling the net primary productivity of temperate forest ecosystems in China with a GAP model. Ecological Modelling 122(3), 225-238.
Kim, C. H., J. G. Oh, E. O. Kang, C. S. Yu, and J. K. Lim, 2014: Community distribution on mountain forest vegetation of the Gyebangsan area in the Odaesan national park, Korea. Korean Journal of Ecology and Environment 47(3), 135- 145. (in Korean with English abstract)
Kim, C. H., M. G. Jo, J. K. Kim, M. S. Choi, J.
M. Chung, J. H. Kim, and H. S. Moon, 2012:
Vegetation change and growing characteristics of Abies koreana population by altitude in Georim valley of Mt. Jiri. Journal of Agriculture & Life Science 46(1), 63-70. (in Korean with English abstract)
Kitao, M., T. T. Lei, T. Koike, H. Tobita, and Y.
Maruyama, 2003: Higher electron transport rate observed at low intercellular CO
2
concentration in long-term drought-acclimated leaves of Japanese mountain birch (Betula ermanii). Physiologia Plantarum 118(3), 406-413.Kitaoka, S., and T. Koike, 2005: Seasonal and yearly variations in light use and nitrogen use by seedlings of four deciduous broad-leaved tree species invading larch plantations. Tree Physiology 25(4), 467-475.
Ko, D. W., J. H. Sung, Y. G. Lee, and C. R. Park, 2015: The current status and challenges of forest landscape models. Journal of Korean Forest Society 104(1), 1-13. (in Korean with English abstract)
Kong, W., S. Lee, H. Park, and J. A. Yu, 2012:
Ecosystem vulnerability assessment of local government due to climate change. Journal of Climate Change Research 3(1), 51-69. (in Korean with English abstract)
Koo, K. A., J. Kim, W. Kong, H. Jung, and G.
Kim, 2016: Projecting the potential distribution of Abies koreana in Korea under the climate change based on RCP scenarios. Journal of the Korean Society of Envrionmental Restoration Technology 19(6), 19-30. (in Korean with English abstract) Koo, K. A., W. K. Park, and W. Kong, 2001:
Dendrochronological analysis of Abies koreana W.
at Mt. Halla, Korea: Effects of climate change on the growths. The Korean Journal of Ecology 24(5), 281-288. (in Korean with English abstract) Korea Forest Service, 2018: The 6th National Forest
Plan. Korea Forest Service, Daejeon, South Korea, 153pp. (in Korean)
Lambrecht, S. C., M. E. Loik, D. W. Inouye, and J.
Harte, 2006: Reproductive and physiological responses to simulated climate warming for four subalpine