140 This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://
creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
J. Mushrooms 2018 September, 16(3): 140-146 http://dx.doi.org/10.14480/JM.2018.16.3. 140 Print ISSN 1738-0294, Online ISSN 2288-8853
© The Korean Society of Mushroom Science
*Corresponding author E-mail : [email protected]
Tel : +82-55-254-1352, Fax : +82-55-254-1319 Received September 4, 2018
Revised September 13, 2018 Accepted September 20, 2018
Breeding and characterization of a long shelf-life cultivar
‘DanBi 7Ho’ by mono-mono crossing in Pleurotus eryngii
Min Keun Kim
1,*, Soon Ae Sim
1, Ji Hye Park
1, Jae San Ryu
2, Si Lim Choi
1, and Kwang Pyo Hong
11
Division of Environment-friendly Research, Gyeongnam Agricultural Research and Extension Services, Jinju 660-360, Republic of Korea
2
Department of mushroom Science, Korea National College of Agriculture and Fisheries, Jeonju 54874, Korea
ABSTRACT:
Pleurotus eryngii is one of the most commercially important mushrooms cultivated in Korea. However, the shelf-life of the fruiting body is short, limiting its export. A new hybrid strain H17 of P. eryngii was developed to extend the shelf-life by mono-mono crossing between monokaryotic strains derived from DanBi and KNR2774. Although the cultivation period of H17 was slightly longer than that of the reference cultivar Kenneutari No.2, the quality did not change and remained normal after a period of 65.0 days at 4°C. This result was significantly different from that of the reference cultivar Kenneutari No.2. Analysis of the genetic characteristics of the new hybrid strain H17 revealed a different profile from that of the parental and reference cultivars when random amplification of polymorphic DNA (RAPD) primers was used. These results demonstrate that H17 is a new cultivar with improved storability after harvesting.
KEYWORDS: King oyster mushroom, Mono-mono crossing, Pleurotus eryngii, Shelf-life
INTRODUCTION
Pleurotus eryngii, also known as the king oyster mushroom, was originally cultivated in northern Italy and Switzerland in the mid 1970s, where it was known locally as cordoncello (Ohga and Royse, 2004) but is now commonly cultivated in Europe, the Middle East, and North America, as well as in parts of Asia.
Cultivation of Pleurotus eryngii on an industrial scale began in Korea in 1996, and by 2016, production of P.
eryngii was estimated to be 48,588tons (Ministry for Food, Agriculture, Forestry and Fisheries, 2016). The king oyster mushroom accounts for more than 29.9% of the edible mushroom market in Korea. The increasing
popularity of P. eryngii among consumers is due to its flavor, texture, and culinary value. Also, it has pharmaceutical value such as antiproliferative action against leukemia cells, and anti-mitogenic activity toward mouse splenocytes (Ngai and Ng, 2006). The value of P. eryngii as an exportable item is very high and it is expected to become a potential source of agricultural revenue in Korea. However, the shelf-life of the fruiting body is short at 4°C and forms an obstacle to its export. Hence, breeding a strain with long shelf- life is key to solve this problem. Breeding edible mushrooms involves various methods, including protoplast fusion (Chakraborty and Sikdar, 2008), genetically engineered transformation (Chen et al., 2000;
Mikosch et al., 2001), restriction enzyme-mediated
integration methods (Kim et al., 2004; Hirano et al.,
2000). Although these techniques are available, they do
not facilitate the development of a new strain for
commercial cultivation. Mycelial mating of monokaryotic
basidiospores is a typical method for the creation of a
new cultivar. Recently, a variety of Pleurotus ostreatus
that could be cultivated under warm conditions was
developed by interbreeding using mating between
monokaryotic mycelia (Gaitán-Hernández and Salmones,
2008). In the breeding of edible mushrooms, new
cultivar such as Dan Bi and Dan Bi 5 Ho were mainly
developed by the method of mycelia mating. (Kim et al., 2011; Lee et al., 2016). The new cultivar DanBi of P. eryngii is characterized by a small number of primordia formation after scratching. The objective of this study is to breed new strain of P. eryngii with long shelf-lives at low temperature. We report the generation of a new cultivar for commercial cultivation.
MATERIALS AND METHODS
Strain and growth conditions
The strains of P. eryngii used in this study were collected from Gyeong Nam Agriculture Research and Extension Service (GNARES), Korea. The original strains and basidiospores were preserved in the mushroom research laboratory, GNARES. The mycelia were maintained on mushroom complete medium (MCM; 20.0 g Glucose, 2.0 g Yeast, 2.0 g Peptone, 0.5 g MgSO
4,0.46 g KH
2PO
4,1.0 g K
2HPO
4per L) at 25
°C by periodic transfer.
Single sporing and crosses
Monokaryotic mycelia were isolated from basidiospore suspensions at a concentration of 1.0 × 10
6spores per mL. The basidiospore suspensions were diluted with sterilized double-distilled water and spread on MCM in Petri dishes (87 mm Ø). After 5–7 d of incubation in darkness at 25°C, the spores germinated and the presence of monokaryotic mycelium was confirmed by microscopically verifying the absence of mycelial clamp connections. Twenty monokaryotic mycelia were isolated for each strain, DanBi and KNR2774. These 40 single spore cultures were paired in every possible combination. Mating was conducted by placing mycelial blocks (5×5 mm) opposite a monokaryotic mycelium on MCM, 10 mm apart. Dikaryotization was confirmed on observing mycelial clamp connections in the area of contact between the monokaryotic mycelia by using the light microscope, after incubation at 25 °C for 8–10 d.
The mycelial growth rate was estimated as the radial growth length of dikaryotic mycelia on MCM. The new dikaryons subsequently underwent fruiting and cultivation.
Fruiting and cultivation methods
The substrate for growth and fruiting of P. eryngii consisted of sawdust from poplar(56%), wheat bran (22%), rice bran(19%), and dried bean-curd dregs(3%).
The final moisture content of the substrate was 65%.
Moist culture medium with the sawdust mixture was loaded into 850 mL polypropylene bottles. They were sterilized at 121°C for 100 min. After autoclaving, bottles were taken out and cooled at 15°C in the cooling room. Inoculated bottles with the spawn were hauled to an incubation room for 35 d, where the temperature and humidity were maintained at 22–24°C and 65–68%, respectively. When the substrate in the bottles was colonized, the bottles were transferred to a cultivation room for 20 d to obtain fruiting bodies. Fruiting was induced by maintaining a low temperature (about 15 °C) and high humidity (about 90–95%). Most of the fruiting bodies except the most significant were removed using a knife. The mushrooms were harvested at the adult stage, when the stipe and pileus were completely extended.
Hardness and storage on fruiting body
The stipes of the hybrid strains were used for measurements of hardness. The stipe was compressed to the bottom and middle of its original height by using a 5 mm aluminum cylindrical probe to achieve compression value of the mushroom sample. The two fruiting bodies of the new hybrid strains were stored in a polyethylene terephthalate box with a lid (190 mm×150 mm×58 mm) at 4°C. The change in quality including color, morphology, and shelf-life after storage was analyzed at intervals of ten days. All the experiments were carried out in 3 replicates.
Antagonistic test and random amplification of polymorphic DNA (RAPD) analysis
The parental, hybrid and control strains were co- cultured in MCM media at 25°C for 14 d. Antagonism between the hybrid and other strains was evaluated.
Genomic DNA was extracted from mycelia using the method described by Lee et al. (1990). Genomic DNA was amplified by the RAPD technique (Williams et al., 1990) in which 5 arbitrary 10-base oligonucleotide primers (Bioneer Co., Daejeon, KOREA) were used to produce amplified fragments. The RAPD polymerase chain reaction (PCR) was performed using a thermal cycler with an initial denaturation stage of 5 min at 95°C, followed by 45 cycles of denaturation for 1 min at 94°C, annealing for 1 min at 37 °C, extension for 2 min at 72°C, and a final 10 min extension at 72°C. The RAPD products were run on a 1.2% agarose gel in 1×
Trisacetate-EDTA buffer for 45 min at 100 V, with a 1.0
kb DNA plus ladder (Bioneer Co., Daejeon, KOREA) as a size marker, and then stained with an EtBr solution (0.5% µg /mL). The stained gels were visualized and photographed using a UV transilluminator.
Statistical analysis
All data were statistically analyzed using the SAS software version 9.3 for Windows (SAS Institute, Cary, NC). The data used were in triplicate. The accession means for the yield and shelf-life measured were used to compare the performance of the hybrid strains to that of parental strains. Accession differences were determined by the Duncan’s multiple range test (DMRT). Statistical differences were considered at the 5.0% level of significance (p <0.05).
RESULTS AND DISCUSSIONS
Strains produced from mono-mono crosses of DanBi and KNR2774
Interbreeding was conducted by mating the 40 monokaryons isolated from the parental strains DanBi and KNR2774. Each of the 20 monokaryotic mycelia of DanBi was mated with the 20 monokaryons of KNR2774, resulting in a total of 400 crosses. Of these, 118 crosses were observed to result in clamp connections. The mating frequency was 30.0%, which was higher than the expected mating rate for the tetrapolar heterothallic sexual pattern (Eugenio and Anderson, 1968). In tetrapolar basidiomycetes, 25% of random intrastock crosses are expected to be compatible (Carlile and Watkinson 1994). These results showed that the parental strains underwent favorable genetic recombination during the mating of compatible monokaryons. During dikaryon evaluation, the radial growth of the crosses was 17.0–72.0 mm/7d on MCM at 25°C, whereas for the parental strains, it was 55.7 mm/
7d for DanBi and 60.0 mm/7d for KNR2774. The 118 mated dikaryons were cultivated on the solid substrate in 850 mL polypropylene bottles. A mean of 34.7 d was required to cover the surface of the medium for all strains. The formation of primordia by removing the mycelia on the top surface of media took about 9.1 d.
The cultivation period for the mated dikaryons until harvest was about 18.2 d. No significant differences were observed between the parental strains (DanBi and KNR2774) and the mated dikaryons.
Fruiting body production and storage of the new dikaryon strains
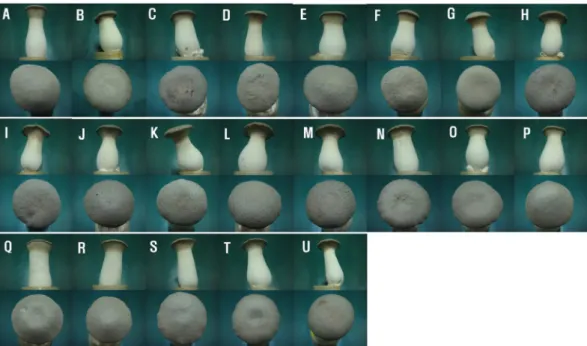
A total of 17 strains were gained by screening for the shelf-life period after harvesting at 4°C (Table 1). The culture time required to harvest the fruiting bodies was 12.7 (H8) to 19.3 (H4) d. The mean production yield of the selected strains was 93.3 g per bottle. The highest yield was recorded for strain H3. The highest of stipe length was recorded for strain H17. Most of the selected strains had a brown color and a convex shape in the pileus (Fig, 1). However, there were significant differences in the shelf-life at 4°C. Most of the new dikaryon strains appeared to wither, turned brown, softened, and gave off a bad odor within 30 d. But, H3, H4, H6, H7, H11, H13, H14, H15, H17, and H18 remained fresh in the stipe and pileus after 30 d. The average shelf-life of the new dikaryon strains was 32.9 d at 4°C. The highest value recorded in this study was 65.0 d for H17. The quality of the fruiting bodies did not change and remained normal in the case of strain H17 after a period of 50 d at 4°C (Fig. 2). This is a significant difference compared to the parental and commercial strains. We named the new hybrid strain H17 “DanBi 7Ho”.
Antagonistic test
Antagonism is a specific example of somatic incompatibility (Huang, 1996). A clear dividing line is formed at the junction between fungal colonies of different genetic backgrounds. To confirm that H17 and the parental strains, including the reference cultivar Kenneutari No.2, have significant differences, an antagonistic test was conducted. The antagonistic test showed that the hybrid strain H17 had different degrees of antagonism resistance to the control strain (Fig. 3).
These results indicated the presence of significant genetic differences between H17 and control strain.
Antagonistic test is a useful and easy method to confirm the development of new hybrid strains.
RAPD analysis
Five arbitrary 10-base oligonucleotide primers were
used to amplify segments of DNA from the new hybrid
strain and the parental strains DanBi and KNR2774. The
reference cultivar Kenneutari No.2 was used a control
for comparison with the new hybrid strain. Among the
tested primers, 2 primers, #8010 (5'-CTGAGACGGA-3')
and #8027 (5'-TGCGCCGCGG-3'), were found to
Table 1. Characteristics of hybrid strains in the bottle cultivation Identification
code Dikaryon
Primordia formation
(days)
Days for harvest
(days)
Stipe length
(mm)
Stipe diameter
(mm)
Pileus diameter
(mm)
Individual weight
(g)
Quality (1-9)
Average of shelf-life (days) DanBi 9.8±0.4a
a17.4±0.3ab 110.8±0.4abcd 43.0±0.2ab 63.9±1.1ab 105.9±3.5abc 6.7±0.1a 42.0±1.4b KNR2774 9.3±0.1a 16.4±0.5abc 92.3±0.9cde 41.8±0.2ab 55.2±1.1b 88.5±0.7abc 5.1±0.2a 34.0±1.4b H1 2X10 8.2±0.3a 15.5±0.2abc 111.6±3.7abc 42.8±4ab 58.5±4.1ab 112.2±6.5ab 6.9±0.5a 28.0±7.1b H2 3X3 9.1±1.1a 18.4±0.1ab 99.5±3.6cde 39.3±4.6ab 58.8±1.6ab 84.9±7.1bc 5.4±0.1a 27.0±8.4b H3 3X4 8.1±0.7a 16.7±0.3ab 101.6±4.2cde 52.3±14.1a 67.1±5.8ab 114.6±13.6a 6.5±0.4a 31.5±4.9b H4 3X10 9.8±0.7a 19.3±0.9a 106.5±10bcde 39.7±4.3ab 57.2±7.6ab 87.8±12.4abc 6.2±1.1a 32.5±20.5b H5 3X12 8.8±1.2a 18.7±0.3ab 103.3±1.9bcde 42.3±5.6ab 58.8±4.5ab 96.0±2.3abc 5.9±0.2a 29.0±8.4b H6 4X19 8.8±1.1a 16.5±0.7abc 100.0±0.5cde 37.2±0.6b 67.5±0.3ab 95.8±0.2abc 6.0±0.4a 35±4.2b H7 5X1 10.3±0.4a 16.5±0.7abc 109.2±1.4abcd 41.3±3.3ab 57.8±0.1ab 101.4±2.1abc 6.7±0.1a 30.0±2.8b H8 5X2 9.2±1.3a 12.6±6.1c 109.6±16.1abcd 34.8±1.6b 62.9±9.0ab 89.8±20.1abc 6.2±0.8a 26.5±7.7b H9 5X7 9.0±1.4a 17.1±1.1ab 91.3±2.4cde 37.0±4.7b 60.1±2.1ab 85.8±8.2bc 5.3±0.5a 24.5±2.1b H10 5X16 9.5±0.7a 16.6±0.4ab 87.4±10.1e 41.9±8.8ab 63.4±0.7ab 97.9±2.7abc 5.1±1.2a 29.5±12.0b H11 5X19 9.0±1.4a 16.5±1.1abc 97.1±3.1cde 38.2±1.2b 70.2±0.6a 90.5±10.1abc 5.5±0.4a 35.0±4.2b H12 8X18 8.7±1.1a 16.7±1.1ab 90.3±10.3de 43.1±8.2ab 57.1±8.7ab 82.8±17.2c 5.1±1.6a 20.0±14.1b H13 11X17 8.1±1.1a 16.5±0.7abc 104.5±4.0bcde 43.1±5.8ab 61.3±1.8ab 93.5±3.5abc 6.1±0.9a 39.5±10.6b H14 15X6 8.3±2.3a 16.0±1.4abc 110.2±5.4abcd 34.4±1.9b 61.1±1.6ab 82.6±6.5c 6.1±0.1a 32.5±0.7b H15 15X15 8.8±1.6a 16.0±1.8abc 103.4±9.8bcde 33.3±2.7b 55.7±2.3b 78.7±16.8c 5.5±1.4a 36.5±17.6b H16 15X18 9.1±1.6a 18.1±2.1ab 101.8±20.0cde 39.5±3.6ab 56.5±15.7b 84.2±28.3bc 5.6±2.1a 27.5±10.6b H17 17X7 8.8±0.2a 15.6±0.1abc 122.7±13.1ab 34.8±6.7b 63.3±7.5ab 103.0±1.8abc 6.4±0.1a 65.0±7.1a H18 17X16 9.0±1.0a 17.5±0.2ab 104.2±10.0bcde 41.5±8.2ab 62.9±1.9ab 86.7±1.1abc 5.6±0.6a 30.0±2.8b Kenneutari No.2 7.5±2.1a 14.9±1.2bc 126.7±6.1a 32.1±2.1b 55.9±0.9b 90.1±18.5abc 6.2±1.1a 23.0±1.4b
a