DOI 10.17480/psk.2017.61.1.30
염증성 호흡기 질환에서 기도점액 과다분비의 병태생리학적 의의 및 조절 방법 탐색
이 현 재#
삼육대학교 교양대학 및 보건관리학과
(Received December 21, 2016; Revised February 10, 2017; Accepted February 13, 2017)
Pathophysiological Significance of Hypersecretion of airway Mucus in Inflammatory Pulmonary Disease and Investigation of Strategies of its Regulation
Hyun Jae Lee#
Department of Health Management and Smith Liberal Arts College, Sahmyook University, 815 Hwarang-ro, Nowon-gu, Seoul 01795, Korea
Abstract — Airway mucus is pivotal for physiological defense against invading particles, chemicals and pathogenic microbes. However, any abnormality of mucins can cause altered airway physiology and impair host defenses leading to severe diseases e.g. asthma and chronic obstructive pulmonary disease. Therefore, in this review, we discussed the recent progress in the field of research on the regulation of the gene expression, production and secretion of airway mucins, in order to make us try to develop the novel efficacious drug candidate for controlling the pathologic hypersecretion of airway mucins.
Keywords airway, mucin, hypersecretion, regulation
기도 점액 과다분비 질환의 병태생리
객담, 점액, 뮤신의 정의
생리적 조건에서, 기도 내부의 점액은 상피세포층을 덮고 있 으며 점액섬모 청소(mucociliary clearance) 기능을 통하여 비산 화학물질, 미세분진 입자, 병원 미생물 등 인체 외부에서 유입되 는 유해인자들을 포집함으로써 기도 내부를 독성 자극으로부터 보호하게 된다.
기도 점액(mucus) 혹은 객담(sputum)은 타액, 혈청 단백질 삼 출물, 박리된 상피세포들과 이들에 함유된 수분, 지질, 당단백질, 단백질, 염류, 효소, 항효소 물질, 산화성 물질, 항산화성 물질, 외
인성 세균, 내인성 항균물질, DNA, 히스타민 등 염증매개물질과 뮤신(점액소, mucin)의 혼합물을 지칭한다.
뮤신(mucin) 이란 serine과 threonine의 아미노산 반복서열 구 조를 가지는 고분자량의 점액성 당단백질(mucous glycoprotein)로 전체 점액 중량의 2% 정도를 차지하는 물질이나, 점액(mucus)에 물리화학적 성질인 점탄성(viscoelasticity)을 부여하는 주요한 생 화학적 구성요소이다.1-4)뮤신은 선상의 펩타이드 서열(apomucin) 에 탄수화물 가지가 부착되어 이루어진 실 모양의 분자로서, 아 포뮤신은 현재까지 동정된 약 20 종의 특정 뮤신 유전자(MUC) 에 의해 그 염기 서열이 결정된다(Fig. 1). 이 가운데 MUC5AC 와 MUC5B 유전자 산물만이 기도 분비 겔 형성 뮤신(gel-forming mucin) 이다. 성숙한 뮤신은 세포막에 부착되어 있거나(막 부착형 뮤신) 혹은 세포막으로부터 분비되고 유리(분비형 뮤신) 된다. 이 들은 핵에서의 유전자 전사, 뮤신의 pre-mRNA 에 대한 전사 후 수식(post-transcriptional modification), mRNA 수송과 조면 소포 체(rER)와의 결합, 번역(translation), co- 또는 post-translational modification 등에 의해 조절된다. 세포막 표면에 존재하는 막 부착 (결합) 뮤신은 섬모와 세포외 기질을 구조적으로 지지하는 역할
#
Corresponding Author Dr. Hyun Jae Lee
Department of Health Management and Smith Liberal Arts Col- lege, Sahmyook University, 815 Hwarang-ro, Nowon-gu, Seoul 01795, Korea
Tel.: 02-3399-1909 Fax.: 02-3399-1876 E-mail: [email protected]
Short Report
종설을 하며 신호전달 경로에도 관여한다. 막 부착 뮤신의 세포외 기 질 단편은 세포막에서 단백분해나 절단에 의해 유리된다. 분비형 뮤신은 분비 과립의 첨부에 분포하게 되며, 조절성 토세포작용 (regulatory exocytosis)에 의해 유리된다.5-6)기도 점액은 섬모와의 적절한 상호작용을 유지하기 위해 점성과 탄성이 이상적인 균형 을 이루고 있고, 점액의 분비선은 잘 발달해 있으며 콜린 작동성 신경과 펩타이드 작동성 신경에 의하여 조절된다.7-8)상피 표면 에 분포하는 점액세포(배상세포)는 비강과 하부 기관 내부의 위 치에 따라 그 수효가 가변적이다.
종합해 보면, 기도 점액은 인간의 기도 배상세포(goblet cell) 및 점막하 점액선(submucosal gland) 등으로부터 생성, 분비되어 기도 내강 표면에 존재하면서 상피세포층을 이루고 있는 또다른 주요 세포인 섬모세포와의 협동적 작용을 통해, 유해한 물질의 제거에 있어서 중요한 역할을 한다. 이처럼, 생리적 상태에서는 뮤신이 생체를 방어하는 역할을 수행하지만 천식이나 만성 폐쇄 성 폐질환(chronic obstructive pulmonary diseases, COPD) 인 만성 기관지염, 기관지 확장증, 폐기종, 낭포성 섬유증 등의 만 성 염증성 호흡기 질환에서 관찰되는 끈끈한 점액의 과다생성 혹 은 과다분비 즉, 뮤신의 양과 질의 이상은 병리적 요소로 작용하 여 점액 축적으로 인한 기관내강 폐쇄, 기도 공기 유입 저해 등 을 통한 질환의 악화를 유발할 수 있다.7, 9-10)
기도 뮤신의 유전자
세포막 부착(결합) 뮤신의 종류 및 그 유전자
세포막 부착(결합) 뮤신과 뮤신양(mucin-like) 물질의 유전자로 는, 13 종의 인체 유래 유전자와 11 종의 마우스 유래 유전자가 Genbank에 등재되어 있다. 이 중에서, MUC1, MUC4, MUC16 등이 폐에서 기본적으로(constitutively) 발현되는 것으로 알려져 있다. MUC1, MUC4, MUC16 등은 세포막 투과 도메인과 당화 도메인 사이에 존재하는 conserved extracellular site에서 아포 단백질이 떨어져 나간다(Fig. 2).
겔 형성 뮤신(gel-forming mucin)의 종류 및 그 유전자 5종의 인간 및 마우스 뮤신 ortholog(다른 생명체에서 발견되는, 같은 기능을 하는 유사 유전자)는 시스테인이 풍부한 von Willebrand 인자 유사 도메인(vWDs) 및 기타 cysteine rich region에 형성된
공유 이황화 결합을 통하여 중합되는 분비형 뮤신으로 구성되어 있다. 이러한 뮤신들을 겔 형성 뮤신이라 지칭한다.
4종의 겔 형성 뮤신(MUC2, MUC5AC, MUC5B, MUC6)은 인 간 염색체 11p15 위치 및 마우스 염색체 7F5에 있는 conserved cluster내의 tandem에 존재하고 있다.11)인간의 경우, 11번 염색 체의 이 부분이 천식에서 기관지 과민반응과 연관 되어 있다.12) 또다른 겔 형성 뮤신인 MUC19도 역시 인간과 마우스 모두에 존재하는 겔 형성 뮤신이지만 11p15 / 7F5 cluster의 일부는 아 닌 것으로 최근 확인되었다.13-14)겔 형성 뮤신은 거대한 아포단 백질 중심부를 가지고 있는 고분자 화합물이다. 이들은 고도로 당화(특히 clustered O-linked tandem 반복 도메인 내에서) 되어 있으며 수분 흡착성이 높고 수천 kDa의 극도로 높은 분자량을 보유한다.
뮤신 유전자들 상호 간에 기능적 반복성을 나타내는 이유는 바 로 이러한 물리적 성질 때문인 것으로 추정되고 있다. 그러나, tandem 반복 도메인 서열, 당화 정도, 위치에서의 이질성(heterogeneity)은 각각의 뮤신이 미세 환경 내에서 각각 특징적인 생리적 기능을 발현하도록 되어 있음을 시사한다. 따라서, 기저 수준에서는 겔 형성 뮤신 유전자의 발현이 조직에 따라 가변적인 것이다. MUC2 는 장관에 발현하는 주요 뮤신이지만, 폐에서도 낮은 수준으로 발현한다. MUC5AC 와 MUC6 는 위에서 발견되는 뮤신이며 이 중에서 MUC5AC는 폐에서도 다량 발현한다. MUC5B와 MUC19 는 타액선, 기관-기관지 점막하 점액선 등에 존재하는 선 뮤신 (glandular mucin) 이다.
가. MUC2
In vivo 에서 MUC2 는 인간 기관-기관지 상피세포에 기저 수 준으로 존재한다. 다수의 연구결과들은 in vitro 기관 상피세포에 서 MUC2 뮤신의 발현을 보여 주었으며, 이러한 연구결과는 MUC2 mRNA 발현 증가 및 MUC2 promoter에 의한 reporter 발현 증가로 이끄는 신호전달 경로의 in vitro 분석 연구를 통해
Fig. 1 − Schematic drawing of a secretory mucin glycoprotein (Rose
and Voynow, 2006).
Fig. 2 − Membrane associated and gel-forming airway mucins
(Evans and Koo, 2009).
확증되었다.15-16)그러나, in vivo에서는 폐쇄성 기도질환에서의 MUC2 발현의 뚜렷한 역할을 일관성 있게 입증할 수 없었다. 단 백질 수준에서, MUC2의 발현은 COPD 환자의 객담 내에 존재 하는 전체 뮤신의 약 2%를 차지한다.17)건강한 정상인과 비교하 여, 천식 환자는 확연히 MUC2 수준의 증가를 보이지만, 이 경 우에도 MUC5AC mRNA 에 비해 MUC2 mRNA 는 1/20 정도 의 낮은 발현 수준을 보인다. 낭포성 섬유증 환자의 경우, 환자가 아닌 경우와 비교했을 때 MUC2 의 증가는 나타나지 않는다.18-19) 다양한 폐 adenocarcinoma 세포주에서 MUC2 가 발현되며, 기도 에서의 MUC2 발현은 특정 종류의 폐암발생과 연관되어 있다. 따 라서, 기도에서의 MUC2 발현은 폐 종양 형성의 표지자로 간주 될 수 있는 것이다.
나. MUC5AC
인간의 폐에서는 기도표면 상피세포에 MUC5AC가 주로 분포 되어 있다.20) MUC5AC 는 건강한 기도의 내강 표면에 분포하 는 배상세포에서도 발현하지만, 천식과 만성 폐쇄성 폐질환에 이 환된 기도에서 가장 다량으로 발현되는 겔 형성 뮤신이다.17)또 한 MUC5AC 는 점막하 점액선의 목 부분을 두르고 있는 상피 세포층과 내강표면에서도 증가되어 있다.21)낭포성 섬유증 환자 의 객담 중에서도 MUC5AC 는 겔 형성 뮤신으로서 가장 우위 를 차지하고 있다.19, 22) 병증이 안정적인 추세를 보이는 환자에 서는 발현이 낮으며 증상이 악화될 경우 약 10배 이상 증가세를 보인다.23) In vitro 의 경우, Pseudomonas aeruginosa, Non- Typeable Haemophilus influenzae, Staphylococcus aureus 와 같 은 pathogen 에 의하여 MUC5AC 발현이 증가될 수 있다.16, 24)마 우스의 경우, Muc5ac 의 발현 위치 및 발현 양식이 인간에서의 그 것과 일치하는 경향을 보인다. 마우스를 모델로 한 천식25) 과 in vivo parainfluenza type I 감염26)에서 muc5ac 발현이 증가한다.
최근에는 겔 형성 뮤신의 유전자 발현에 대한 포괄적 분석을 통 해, 항원자극 마우스에서도 muc5ac 발현이 뚜렷한 증가를 보인 다는 결과가 보고되었다.27)그러므로, 마우스의 폐에서 점액성 세포로의 화생에 있어 muc5ac 발현의 증가는 가장 중심적이고 결정적인 현상인 것이다.
다. MUC5B
MUC5B 는 기관-기관지에 분포하는 선 점액세포 내에서 기본적 으로 발현된다.21-22)인간 기도 표면 상피세포의 겔 형성 뮤신 중 두 번째로 높은 수준으로, MUC5AC 발현 수준의 약 20% 정도로 발현된다. MUC5AC 와는 달리 MUC5B 는 건강한 대조군과 비교 했을 때 흡연자28)와 천식환자21)의 기도에서 미약한 생성량 변화 를 보인다. 인간 폐에서 관찰되는 양상과 흡사하게 마우스 역시 기 저 수준에서 폐에서 Muc5b 가 풍부하게 발현되며,27)항원에 노출 된 마우스라고 해서 Muc5b 발현이 증가하지는 않는다.
라. MUC19
MUC19도 겔 형성 뮤신의 일종으로 11p15 / 7F5 cluster의 일 부는 아닌 뮤신이다.29-30)인간의 중이31)와 안구표면 상피세포 에,32)인간과 마우스 설하선, 기관 점막하 선의 점액세포에 발현 하는 것으로 알려져 있다. 폐에서는, 기저 수준에서 Muc5ac 와 Muc5b 에 비해 1/0−1/100 수준으로 발현된다.27)인간에서 이 유 전자의 ortholog 가 염색체12q12 위치에 존재하지만, 이 유전자 가 폐의 항상성 유지 및 병태생리에 어떻게 관여하고 있는 지에 대해서는 아직 규명되지 않았다.
폐에서의 뮤신 유전자 발현의 조절
MUC5AC 와 MUC5B 유전자의 조절 기전은 인간 및 동물 실 험모델을 이용한 in vitro 나 in vivo 연구를 통해 규명되고 있다.
기관지 brushing, 생검, 조직 검체 등은 in vivo에서 인간 기도상 피 세포에서의 주요 겔 형성 뮤신임을 일관되게 증명하고 있다.
In vitro에서 이들의 조절 기전을 연구하기 위해, 인간 기도 유래 세포주나 일차배양 인체 기도 상피세포가 주로 활용된다. 이들 세 포 시스템의 장점은 인간 뮤신 유전자 조절에 관한 정보를 얻을 수 있다는 점이다. 동물모델 역시 널리 사용되고 있으며 가장 큰 장점은, 유전학적 조작이 가능하며 잘 통제된 조건에서 실험적 으로 질환 표현형을 유도할 수 있다는 점이다(Fig. 3).
기도 점액 과다분비 질환의 병태생리
일반적으로, 점액 과다분비는 계절성 알러지나 상기도 감염과 같은 상황에 동반되는 가벼운 문제로 인식되고 있으나, 천식이 나 만성 폐쇄성 폐질환, 낭포성 섬유증 등의 질환에서는 점액 과 다분비가 질병의 중증도와 사망율을 높이는 요인이 된다. 기도
Fig. 3 − Model of biosynthesis and secretion of mucin glycoproteins
in a goblet or mucous cell (Rose and Voynow, 2006).
의 염증, 뮤신 발현의 변화와 점액성 세포로의 화생(mucous metaplasia), 점액 과다분비, 점액섬모 청소 기능의 손상 등은 모 든 폐쇄성 폐질환에서 공통적으로 관찰되는 병리학적 특징이다.
즉, 뮤신의 생성 변화로 인한 뮤신의 유형이나 양이 변화하고 세 포 내부 분비과립의 부피 비중이 증가하게 되며, 토세포 작용 (exocytosis)의 증대로 세포외 점액 겔의 점성과 두께가 동시에 증가하게 된다. 이로 인하여 기도 폐쇄 및 점액섬모 청소기능의 손상이 유발되고, 그 결과 공기의 기도 출입이 제한되게 되며 미 세입자/병원체의 기관 내부 정체를 야기하게 된다.
천식의 병태생리
천식에서, 환기 차단은 기도 평활근 수축에서의 염증 유발성 변화와 점액 과다분비로 인하여 일어난다. 대부분 환자에서 평 활근 수축은 β2 아드레날린 수용체 작용제의 흡입으로 신속히 완화되나, 병리적 점액의 성상은 즉각적 영향을 받지 않는다. 천식 의 일반적 발병인자는 알러지성, 감염성(특히 바이러스에 의한) 자 극 등이다. 소구경 − 중구경 기도를 막고 있는 광범위한 점액전 (mucus plug) 들은 실제로 기침반사에 의하여 제대로 제거되지 않으며 천식으로 사망한 환자의 부검에서 전형적으로 관찰되는 징후이다.33-36)
뮤신은 이러한 점액전의 가장 핵심적인 고분자 구성요소이
며,37-38) 이런 과다분비가 천식의 치명성을 야기하는 것으로 보인
다. 천식으로 인해 사망한 환자에서 전체 연구사례 98% 에 대한 기 도 절단 관찰 시에 내강의 폐쇄가 약 20−100% 범위에서 확인되 었다.39) 천식에서 과다한 뮤신분비가 환기 폐쇄의 주요 원인임 이 항원노출 마우스 실험을 통해 확인되었으며, 화생(metaplasia) 이 발생한 기도에서 뮤신의 세포외 배출을 억제함으로써 기도의 통기성을 현격히 개선할 수 있음이 보고되었다.40) 경증 혹은 중 등도 수준의 천식환자의 기도 표면 상피세포층에서 배상세포의 수가 증가되어 있으며,21, 41) MUC5AC 와 MUC5B 는 천식 환 자의 기도 내 주요 분비형 뮤신으로 특히 MUC5AC 의 경우 건 강인에 비해 현저히 발현이 증가되어 있다.17) MUC5B 는 인간 의 점막하 점액선의 세포에서 기본적으로 생성되고 분비되며 천 식환자의 경우에도, 점막하 점액선 및 기도 표면 상피세포에서 확인되고 있다.20)마우스의 경우에 Muc5b는 mRNA 수준27)및 단백질 수준에서42)공히 기본적으로 발현된다. 인간과는 달리, 마우스에게는 기관에서 원위(distal) 인 기도에는 점막하 선이 존 재하지 않으며, 마우스 폐에서 기본적인 Muc5b 생성은 기도 표 면에 분포하는 Clara cell에 국한된다. 알러지 동물모델에서 Muc5ac 는 주로 발현이 유도되는 겔 형성 뮤신 유전자이며 Muc5b 발현과 분포는 오히려 안정적 양상을 보인다.25, 13)건강한 상태와 호흡기 질환 이환 시의 뮤신 생성 양상의 차이에 대한 생리학적 및 임상적 의의는 아직 완전히 규명되지 않았으며, 유전자 녹아 웃 마우스 등을 이용한 후속 연구를 통하여 규명될 전망이다.
만성 폐쇄성 폐질환(chronic obstructive pulmonary disease) 의 병태생리
만성 폐쇄성 폐질환(COPD)은 구미 지역에서 지난 20년간 2 배 이상의 사망률 증가를 나타낸, 4위의 주요 사망원인이다.43) 대부분 COPD 환자에게 나타나는 점막하 점액선 비대와 기도 배 상세포로의 화생은 만성 기관지염의 현저한 특징이다. 기침과 객 담 배출은 직경이 큰 기도에서 과다 점액을 제거하는데 도움이 되지만, COPD에서의 점액섬모 청소기능의 손상으로 인해 기침 만으로는 점액 배출이 어려운 직경 2 mm 이하의 소구경의 기도 에서는 제대로 점액을 제거할 수 없게 된다. 천식이나 낭포성 섬 유증과 마찬가지로, 만성 폐쇄성 폐질환에서도 소구경 기도 내 강에 과량의 염증성 점액 삼출물의 축적이 관찰된다.44)
따라서, 폐기능 저하, 호흡기 감염, 병원 입원, 사망 등의 현상 이 만성적 점액 배출과 확실히 연관되어 있음을 알 수 있다.45-46) 수술 후 얻은 검체에 대한 조직병리학적 소견은, COPD 환자의 소구경 기도 내강에서 배상세포 수의 증가와 MUC5AC, MUC5B 생성 및 분비의 증가를 확연히 보여주고 있다.47-48)이러한 소견 은 수술 전 FEV1 값과 역의 상관 관계를 보인다. 즉, FEV1 값 이 높게 나타나는 환자는 그 값이 낮은 환자에 비해 배상세포로 의 화생 정도가 더 낮은데, 이는 기도에 뮤신 생성 세포의 존재가 환기 차단의 증가와 연관되어 있음을 시사하고 있는 것이다.49-50) 종합하면, 이 결과는 소구경 기도에서 생리적 및 임상적으로 측 정할 수 있는 기계적 폐쇄가 점액분비 때문에 야기될 수 있으며, 질환의 발생과 예후에 분명히 영향을 줄 수 있음을 뜻한다.
인간의 COPD 발생 주원인은 흡연이다. 마우스에서, 만성적 담배연기 노출은 점액성 세포로의 화생을 야기한다.51)담배연기 그 자체는 in vitro에서 EGFR cascade 활성화에 의해 뮤신 생 성을 직접적으로 증가시킨다.52) 담배연기에 존재하는 여러 가지 잠재적 독성물질들 중 하나인 acrolein(acrylic aldehyde)은 실험 동물에서 점액성 세포로의 화생과 Muc5ac 생성을 유도한다.53) Acrolein은 또한 인간 기도상피 세포주에서 MUC5AC 생성을 촉 진하며, COPD 환자의 호기 농축물과 유도배출 객담 내에 상당한 수준으로 증가되어 있다.54-55)점액은 때로 비정형성 H. influenzae 와 같은 세균성 병원체의 부착과 성장에 적합한 매개체로 작용할 수 있음을 시사하는 보고도 있다.56)그람 양성 및 음성균의 생성 물은 MUC5AC 및 MUC2 유전자 발현과 in vtro 에서 인간 호 흡기 상피세포주에서의 뮤신 분비를 증진시키며,24)비슷한 경향 이 일부 동물모델의 in vivo 실험에서도 관찰되었다.57)바이러스 감염 또한 인간의 COPD 악화에 밀접한 연관성이 있다.58-61)흡 연하는 COPD 환자의 수술 후 검체와 비흡연자의 검체를 비교 해 보면 말초 기도상피세포층에서 배상세포의 수가 현저히 증가 되어 있음을 알 수 있다. 이런 현상은 또한 macrophage와 CD8 positive T-lymphocyte 수의 증가와 함께 나타나며,62)이들은 모 두 바이러스 감염을 의미하는 요소이기도 하다.63) In vivo 에서
세균 혹은 바이러스에 의한 염증 초기단계에 IL-6 생성이 증가된 다. 이로 인해, COPD 환자와,64-65) 실험적으로 사람과66-68) 마우
스에서66-67)유도된 바이러스성 호흡기 감염에서 IL-6 수준이 증
가됨을 알 수 있었다.
기도 염증에서의 뮤신 과다 생성
뮤신 발현은 다양한 종류의 사이토카인 발현 과정을 야기하는 광범위한 염증성 자극에 반응함으로써 유도된다. 현재까지 이런 과정을 가장 특징적으로 보여주는 예는 알러지성 기도 염증과 관 련된 점액 생성 작용이다. 마우스가 항원에 감작된 후 항원 에어 로졸에 단회 노출되었을 때 급성 호 중성구 반응(노출 후 2−6시 간)에 이어 지연성 Type II T-helper(Th2) 임파구 및 호산성구 반응(노출 후 1−7일) 이 연속적으로 일어난다.69-70) mRNA 와 단 백질 수준에서 증가된 뮤신 생성이 지연성 반응 중에 나타나지 만 급성 반응 시에는 나타나지 않는다.69-71, 27)
그러나, 알러지성 기도 질환에서 호산성구의 역할에 대하여 아 직 논란의 여지가 있는 것은, 호산성구를 제거하면 알러지 유발원 에 의한 기도 과민성도 차단된다는 일부 연구결과가 있기는 하지 만 다른 연구에서는 같은 결과가 나오지 않았기 때문이다.72-74)유 전적으로 호산성구를 제거하면 점액성 세포로의 화생이 −50%
수준까지 감소되며,73)항원에 노출된 호산성구 결핍 마우스에 대 하여 호산성구를 재보충(reconstitution) 해 주었을 때 점액성 세 포로의 화생 수준이 정상 동물의 수준으로 회복된다.73)
중요한 것은, 이런 효과가 Th2 세포 의존적이며 항원감작 되 지 않은 마우스에게 호산성구를 투여하였을 때에는 나타나지 않 는다는 점이다.73)그러므로, 호산성구가 점액성 세포로의 화생을 촉발할 수는 있지만 혼자만의 작용으로는 충분치 않다는 것이다.
알러지성 염증은 점액성 세포로의 화생을 연구함에 있어 특성이 가장 잘 규명된 실험모델이기는 하나, 그 외의 다른 동물모델 또 한 계속 시험되어 왔다. 여기에는 감염성, 화학 물질, 환경적 접 촉 뿐 아니라 사이토카인 투여 및 과다발현 모델 등이 있다. 바 이러스 감염은 유전적으로 민감한 마우스 종에서 점액성 세포로 의 화생을 유도하게 된다. 흥미로운 것은, 점액성 세포들이 염증 의 해소 및 바이러스의 소실 단계 후 나타나게 되며 동물의 여명 기간 동안 지속된다는 점이다.26) S. aureus, Burkholderia cepaiciae 등에 의한 호흡기 감염증이 또한 점액성 세포로의 화생을 촉발함 이 보고되었다.75, 16, 24)선천성 면역 사이토카인의 활성화가 이 과정에 어떤 역할을 할 것으로 추정되며 주로 in vitro 실험을 이 용하여 연구되고 있다. 살아있는 Aspergillus fumigatus 의 감염 이 마우스에서 점액성 세포로의 화생을 유도하는지 알려지지 않 았지만, 다수의 동물 모델은 A. fumigatus 점적 흡입투여가, 감작 된 마우스에서 점액성 세포로의 화생을 유도하였음을 보여주고 있 다.76-77) A. fumigatus 추출물과 정제한 A. fumigatus protease를 사 용한 경우에서도 역시 같은 작용이 나타난 바, 뮤신 생성을 도와
주는 A. fumigatus 의 점적 흡입투여 효과는 아마도 Th2-유도 염증에 의해 매개되는 것으로 보인다.
점액성 세포로의 화생(mucous metaplasia)을 유도하는 신 호전달 경로
특정 선천 및 적응 면역반응은 in vitro 와 in vivo 에서 점액성 세포로의 화생을 야기하게 된다. 최근의 연구에 의하면 이런 반응 들은 상호 배타적이기 보다는 오히려 선천 면역 신호가 전반적으 로 적응 면역 신호의 발달, 활성화, 뮤신 생성 기능을 증폭시키는 것으로 추정되고 있다.
가. 염증성 사이토카인 신호전달 경로
보체와 유사하게, proinflammatory cytokines(IL-1 and TNF) 와 inflammatory cytokines(IL-6 and interferons(IFNs)) 는 감 염성 자극이나 손상에 이어 신속하게 유도된다. 이러한 사이토 카인의 생성은 선천 면역 반응 자극을 강화시킨다. IL-1 과 TNF 는, chemokines(IL-8)과 세포부착 분자들(ICAM-1)의 유도를 경 유한 호중성구 및 다른 백혈구들의 동원과 IL-6의 유도를 포함 하는, acute phase response에서의 매개체들이다. IL-6는 B, T- 임파구를 활성화하며, 그로 인해 적응 면역 반응을 촉발하게 된 다. 인터페론은 바이러스에 의해 유도되며, 역시 T-임파구 분화 에 기여한다.
나. 성장인자 신호전달 경로
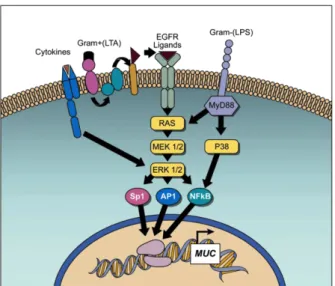
최근 연구에 의하여, 세포 및 조직의 발생, 성장, 자극, 복구 등의 과정을 조절하는 신호들이 점액성 세포로의 화생 과정에 관 여한다는 사실을 알 게 되었다. 이들은 염증성 신호전달 경로와 평행하게 혹은 순차적으로 작용하거나 몇몇 경우에서는 염증과 는 독립적으로 작용하기도 한다. 여기에 해당하는 신호전달 경로 로는, 상피세포 성장인자 수용체(EGFR), transforming growth factor(TGF)-β, Wnt/β-catenin, HIF-1 신호전달 경로 등이 있다 (Fig. 4).
이들 중 다수는, 염증성 자극이나 세포 손상에 반응하여 활성
화되며,78, 54)현재 이런 경로들 간의 중요한 상호작용을 규명 중
에 있다. 그 중에서 EGFR 신호전달 경로를 중심으로 살펴보면 다음과 같다. 점액성 세포로의 화생의 발달 과정에서 EGFR 의 역할은, 항원노출 흰쥐에서 EGFR tyrosine kinase 의 활성 억 제가 점액성 세포로의 화생을 차단할 수 있음을 보여준 Takeyama 등79) 에 의해 제시되었다.
흥미로운 점은, 항원에 노출되지 않은(naive) 동물에서는 EGFR ligand 인 TGF-α의 직접적 투여에 의해서 점액성 세포로의 화생 이 야기되지 않았다는 점이다. 동물실험 및 잘 분화된 기도 상피 세포에서, EGFR은 기저부 세포막(basolateral plasma membrane) 에 국한되는 양상을 보인다. 그러나, 치밀 결합(tight junction)에 대
한 기계적 손상이 일어나면 EGFR이 첨부 세포막(apical plasma membrane)으로 전위하고 그 위치에서 리간드의 자극을 받을 수 있게 된다.80) Sendai virus, cigarette smoke, chemical toxicants, proteases와 같은 매우 다양한 종류의 신호들에 반응하여 유발되 는 in vitro 점액성 세포로의 화생 유도에 있어서도 EGFR 활성 화가 요구된다.
천식,81-83)낭포성 섬유증,84) COPD85-86)환자의 기도상피에서
EGFR 의 발현이 증가된다. EGFR 활성화는 항원, 바이러스, 호 중성구, 흡연 등에 반응하여 나타나는 인간 기도상피 세포에서의 MUC5AC 발현 항진과, 동물모델에서의 Muc5ac 유도와 점액성 세 포로의 화생에 있어 중요하게 작용함이 보고되어 있다.79, 87, 89, 52, 90)
비록 EGFR 자극에 의하여 활성화되는 상위의 신호전달 경로는 이 미 잘 알려져 있지만, EGFR 신호전달 하위 경로에서 MUC5AC/
Muc5ac 유전자 발현을 유도하는 특정 전사인자들에 대해서는 Muc5ac promoter 의 EGF- 매개성 HIF-1 활성화 현상을 발견 한 최근의 연구를 통하여 정보를 얻게 되었다.
기도 점액 과다분비 질환의 조절 방법론
기도 점액 과다생성 조절
점액세포의 분화, 뮤신 유전자 발현, 뮤신 당단백 합성, 뮤신 세포외 배출(exocytosis) 등 각 단계의 조절 기전에 대해 더 잘 이해하게 됨으로써 향후에는 뮤신 과다생성과 분비를 직접적으 로 조절할 수 있는 방법을 개발하는 방향으로 연구가 진행될 것 으로 추정된다.
천식에서 glucocorticoid 흡입과 같은 항염증 약물의 사용에 의
한 뮤신생성 차단은 가장 일반적이고 효율성이 높은 치료방법으 로 이용된다. 그러나, 병인(etiology)에 있어서 낭포성 섬유증(CF) 이나 COPD의 경우에는 천식의 그것과는 확연히 다른 면이 있기 에 이같은 시도가 그다지 효과를 얻지 못할 수 있다. 천식에서는 알러지성 염증으로 인해 거의 대부분 점액성 세포로의 화생 (mucous metaplasia)이 유발되는데, 이는 당질 코티코이드 (glucocorticoid) 투여로 조절될 수 있다. 그러나, CF와 COPD에 서 관찰되는 염증 반응은 당질 코티코이드 투여로 뚜렷하게 개 선되지 않는다.
점액성 세포로의 화생(mucous metaplasia)에 대한 더 선택적 치료방법을 찾기 위해서 이와 관련이 있는 뮤신과, 뮤신생성을 유도하는 신호전달 경로, 뮤신 유전자 발현을 활성화하는 전사 인자들, 뮤신 생성을 조절하는 생합성 기전, 뮤신 유리(release) 를 조절하는 세포외 방출 기전 등에 대해 관심이 집중되어야 할 것이다.
한 예로, 뮤신 유전자 발현을 활성화하는 전사인자들의 조절 을 통한 뮤신 과다생성 억제 전략에 관하여 요점 위주로 살펴보 면 다음과 같다. 유전자 발현 조절 수준에서 작용하는 특정 전사 인자들이 결합하는 것으로 추정되는 부위가 MUC2(GenBank accession numbers U6716791)와 U6806192)), MUC5AC (GenBank accession number AF01683493)), MUC5B (GenBank accession numbers AJ01158294) 와 AJ01245395)) 등에서 각각 확인되었다.
MUC2 promoter는 5
'
-flanking region에 TATA box 와 activator protein(AP)-1, AP-2, CREB, C/EBP transcription factor 들의 다양한 부착 부위 및 하나의 E-box 를 포함한다.92) MUC5AC promoter도 functional CRE site 인 TATA box96-97) 와, Sp1, AP-2, glucocorticoid 수용체(GR), ETV4, NF-κB 전사인자 등의 다양한 부착 부위를 포함한다.93) 따라서, 이들 전사인자들의 활 성을 조절함으로써 MUC5AC 뮤신의 생성을 조절할 수 있을 가 능성에 대한 검증이 요구된다 하겠다.기도 점액 과다분비 조절
겔 형성 뮤신은 일단 합성이 된 후, 조절성 토세포 작용 (regulated exocytosis) 으로 인한 분비(유리) 자극을 받기까지 일 명 분비과립으로도 불리는 세포내 저장낭에 보관된다. 분비 산 물의 합성률에 의해 주로 조절되는 기본적 토세포 작용 (constitutive exocytosis) 과는 달리, 조절성 토세포 작용은 세포 외 리간드에 반응하여 분비가 활성화된다. 상당수의 리간드가 뮤 신 분비를 자극하는 것으로 알려져 있다. 여기에는 염증 매개물 질, 신경 전달물질, 뉴클레오티드, 뉴클레오시드 등이 있다.98)이 들 가운데, 뉴클레오티드인 ATP 와 UTP 는 뮤신 유리를 강력 히 유도할 수 있고 생리적으로도 가장 개연성이 높다.99-100, 71)
ATP 는 표면 점액세포에 대한 기계적, 자극성, 염증성 자극에 반 응하여 유리된다.101-102)
Fig. 4 − Schematic of signal transduction pathways that regulate
mucin gene transcription (Rose and Voynow, 2006).
마우스에서, 흡입된 methacholine 에 의하여 표면 점액세포의 탈과립이 거의 완전하게 일어난다. 무스카린 수용체 mRNA 가 표면 상피세포에 국한되어 있는데, 이는 상피세포에 대한 직접 적 콜린성 작용이 뮤신분비를 자극할 수 있음을 시사하고 있는 것이다. 그러나, 이런 현상이 단백질 및 기능적 수준에서 확증되 지는 않았다. In vivo 에서 뮤신분비를 유도하는 콜린성 자극과 관련하여 가능한 하나의 가설은, 콜린성 자극에 의한 기도 평활 근 수축에 후속되는 기계적 변형에 반응하여 상피세포가 ATP를 유리할 수 있다는 가설이다. P2Y2 퓨린성 수용체는 기도 상피세 포 표면 첨부에 존재하며, 이를 자극하게 되면 칼슘 유리, diacylglycerol 형성, G protein 활성화를 경유한 조절성 토세포 작용의 활성화가 야기된다. 조절성 토세포 작용은, 표면 첨부로 SG 수송, 첨부 피층 액틴 장벽의 해리, 세포막 내부 소엽과 연관 된 tethering 및 docking 단백질과 SG 와의 결합, 세포막과 SG 와의 칼슘 의존성 융합으로 이어지는 신호전달 경로의 활성화를 야기하는 세포내 혹은 세포외 자극이 포함된 과정이다. 이 과정 의 각 단계는 약 12개 이상 단백질의 협동작용을 필요로 한다.
기도 점액 과다분비 질환의 치료제 개발 전략
기존 사용 약물의 약리
지금까지, 기도 점액 과다분비 질환의 치료 전략으로는 corticosteroid 와 같은 항 염증제를 사용하여 점액성 세포로의 화 생(mucous metaplasia)을 감소시키거나, 식염수 분무-흡입, DNA severing enzyme 혹은 thiol 함유 약물을 이용한 점액의 점성 감 소, beta-agonist 를 통한 섬모운동 자극의 유발 등을 들 수 있으나 불행하게도 이들 요법은 효율의 제한성을 극복하기 어려운 바, 새 롭고도 근본적인 치료 전략이 절실히 요구되고 있다.
현재, 호흡기 질환의 임상에서 기도 점액분비 이상의 치료를 위해서 1) 점액성 분비물의 점도를 낮추어 분비물의 배출을 용 이하게 하는 방법, 2) 분비물의 배출을 더욱 자극하여 분비물을 용이하게 다량 배출시키려는 방법 등이 사용되어 왔다.103)첫 번 째 방법인 점액성 분비물의 점도를 낮추기 위해서는 점액 용해제 (mucolytics)를 이용해 왔다. 점액 용해제에 속하는 약물로는 ambroxol, bromhexine, acetylcysteine, S-carboxymethylcysteine 등의 약물이 있는데, 이 약물군의 투여로 점액의 점도 저하가 과 다할 경우, 오히려 점액이 원위 기관지로 흘러 들어가, 기관지 폐 쇄가 더욱 악화될 수 있다.103)따라서, 현재 호흡기 질환의 임상 에서 원칙적으로 권장되는 방법은 환자에게 충분한 양의 수분 보 급을 통해 기침을 하여 객담이 잘 배출될 정도로 점도를 조절하 는 방법인데 약물 투여만으로는 어렵다고 보고 있다.
두 번째 방법인 분비물 배출을 더욱 자극하는 방법은 소위 거 담제(expectorants) 를 투여하는 방법인데, 기도에 대한 직접적 자극 혹은 미주신경-위 반사(vagal gastric reflex) 를 경유하여 점
액선의 분비를 증가시킴으로써 분비물의 배출을 자극하는 방법 을 의미한다. 거담제에 속하는 약물로는 길경, 용담, 세네가, 원 지 등 saponin 함유 생약류, NH4Cl, KI, NH4I, guaiacol 등이 있 는데, 작용 기전이 규명되어 있지 않으며 투여함으로써 얻어지 는 치료적 이익이 부작용에 비해 크지 않다는 평가가 있다.103)
기도 점액 과다분비 질환의 치료제(조절제) 개발 전략 과도하게 생성되고 분비되는 점액을 기도에서 제거하거나 조 절하는 방법은 이론적으로 크게 두 가지 방법이 있을 수 있다.
첫째는, 점액의 점도를 낮추는 약물을 투여하여 흡인하는 물리 적 방법이 될 수 있겠고, 둘째는, 점액의 과다생성 및 분비를 억 제하는 약물을 투여하는 방법이 될 것이다.
그러나, 점액의 농도를 묽어지게 하여 흡입하는 방법은 임상 적으로 기도 내벽의 자극에 따른 과민반응으로 도리어 반사적 점 액 과다분비를 야기할 수 있으므로, 약리학적 방법으로 뮤신의 분비 혹은 생성을 조절(억제) 하는 것이야 말로 기도점액 과다 분비의 조절에 대한 매우 중요한 접근방법이라 할 수 있을 것이다.
그러므로, 아직까지 구체적으로 규명되거나 발견되지는 않았 지만, 언급한 두 번째의 방법인 점액 과다생성과 분비(유리)를 억 제하기 위한 새로운 작용기전의 약물 개발은, 향후 호흡기 질환 자에서 기도점액 과다분비 조절을 가능하게 하는 중요한 접근 방 법이 될 수 있을 것이다.
References
1) Newhouse, M. T. and Biennenstock, J. : Respiratory tract defense mechanism, In, textbook of pulmonary disease (Baum, G.L. and Wolinsky, E.(eds)), 3rd ed., Little Brown and Company (1983).
2) Kim, K. C. and Brody, J. S. : Gel contraction causes mucin release in primary rat tracheal epithelial cells growing on a collagen gel.
J. Cell Biol. 105, 158 (1987).
3) Culpitt, S. V., Rogers, D. F., Traves, S. L., Barnes, P. J. and Donnelly, L. E. : Sputum matrix metalloproteases: comparison between chronic obstructive pulmonary disease and asthma.
Respir. Med. 99(6), 703 (2005).
4) Voynow, J. A. and Rubin, B. K. : Mucins, mucus, and sputum.
Chest 135(2), 505 (2009).
5) Wine, J. J. and Joo, N. S. : Submucosal glands and airway defense.
Proc. Am. Thorac. Soc. 1, 47 (2004).
6) Evans, C. M. and Koo, J. S. : Airway mucus: the good, the bad, the sticky. Pharmacol. Ther. Mar 121(3), 332 (2009).
7) Rogers, D. F. and Barnes, P. J. : Treatment of airway mucus hypersecretion. Ann. Med. 38(2), 116 (2006).
8) Wine, J. J. : Parasympathetic control of airway submucosal glands: central reflexes and the airway intrinsic nervous system. Auton. Neurosci. 133, 35 (2007).
9) Del Donno, M., Bittesnich, D., Chetta, A., Olivieri, D. and Lopez- Vidriero, M. T. : The effect of inflammation on mucociliaryclearance in asthma: an overview Chest 118, 1142 (2000).
10) Rogers, D. F. : Airway mucus hypersecretion in asthma: an undervalued pathology? Curr. Opin. Pharmacol. Jun; 4(3), 241 (2004).
11) Escande, F., Porchet, N., Bernigaud, A., Petitprez, D., Aubert, JP. and Buisine, M. P. : The mouse secreted gel-forming mucin gene cluster. Biochim. Biophys. Acta. 1676, 240 (2004).
12) The Collaborative Study on the Genetics of Asthma. A genome- wide search for asthma susceptibility loci in ethnically diverse populations. Nat. Genet. 15, 389 (1997).
13) Chen, Y., Zhao, Y. H., Di, Y. P. and Wu, R. : Characterization of human mucin 5B gene expression in airway epithelium and the genomic clone of the amino-terminal and 5'-flanking region. Am. J.
Respir. Cell. Mol. Biol. 25, 542 (2001)
14) Culp, D. J., Latchney, L. R., Fallon, M. A., Denny, P. A., Denny, P. C., Couwenhoven, R. I. and Chuang, S. : The gene encoding mouse Muc19: cDNA, genomic organization and relationship to Smgc.
Physiol. Genomics. 19, 303 (2004).
15) Jono, H., Shuto, T., Xu, H., Kai, H., Lim, D. J., Gum, J. R. Jr., Kim, Y. S., Yamaoka, S., Feng, X. H. and Li, J. D. : Transforming growth factor-beta -Smad signaling pathway cooperates with NF-kappa B to mediate nontypeable Haemophilus influenzae-induced MUC2 mucin transcription. J. Biol. Chem. 277, 45547 (2002).
16) Li, J. D., Dohrman, A. F., Gallup, M., Miyata, S., Gum, J. R., Kim, Y. S., Nadel, J. A., Prince, A., and Basbaum, C. B. : Transcriptional activation of mucin by Pseudomonas aeruginosa lipopolysaccharide in the pathogenesis of cystic fibrosis lung disease. Proc. Natl. Acad.
Sci. U S A. 94(3), 967 (1997).
17) Kirkham, S., Sheehan, J. K., Knight, D., Richardson, P. S., &
Thornton, D. J. : Heterogeneity of airways mucus: variations in the amounts and glycoforms of the major oligomeric mucins MUC5AC and MUC5B. Biochem. J. 361, 537 (2002).
18) Hovenberg, H. W., Davies, J. R., Herrmann, A., Linden, C. J. &
Carlstedt, I. : MUC5AC, but not MUC2, is a prominent mucin in respiratory secretions. Glycoconj. J. 13, 839 (1996).
19) Davies, J. R., Svitacheva, N., Lannefors, L., Kornfalt, R. &
Carlstedt, I. : Identification of MUC5B, MUC5AC and small amounts of MUC2 mucins in cystic fibrosis airway secretions.
Biochem. J. 344(Pt 2), 321 (1999).
20) Groneberg, D. A., Eynott, P. R., Lim, S., Oates, T., Wu, R., Carlstedt, I., Roberts, P., McCann B., Nicholson, A. G, Harrison, B.
D. and Chung, K. F. : Expression of respiratory mucins in fatal status asthmaticus and mild asthma. Histopathology 40, 367 (2002).
21) Ordonez, C. L., Khashayar, R., Wong, H. H., Ferrando, R., Wu, R., Hyde, D. M., Hotchkiss, J. A, Zhang, Y., Novikov, A., Dolganov, G.
and Fahy, J. V. : Mild and moderate asthma is associated with airway goblet cell hyperplasia and abnormalities in mucin gene
expression. Am. J. Respir. Crit. Care Med. 163, 517 (2001).
22) Groneberg, D. A., Eynott, P. R., Oates, T., Lim, S., Wu, R., Carlstedt, I., Nicholson, A. G. and Chung, K. F. : Expression of MUC5AC and MUC5B mucins in normal and cystic fibrosis lung. Respir. Med. 96, 81 (2002).
23) Henke, M. O., John, G., Germann, M., Lindemann, H. and Rubin, B. K. : MUC5AC and MUC5B mucins increase in cystic fibrosis airway secretions during pulmonary exacerbation. Am.
J. Respir. Crit. Care. Med. 175, 816 (2007).
24) Dohrman, A., Miyata, S., Gallup, M., Li, J. D., Chapelin, C., Coste, A., Escudier, E., Nadel, J. and Basbaum, C. : Mucin gene (MUC 2 and MUC 5AC) upregulation by Gram-positive and Gram-negative bacteria. Biochim. Biophys. Acta. 1406(3), 251 (1998).
25) Zudhi Alimam, M., Piazza, F. M., Selby, D. M., Letwin, N., Huang, L. and Rose, M. C. : Muc-5/5ac mucin messenger RNA and protein expression is a marker of goblet cell metaplasia in murine airways. Am. J. Respir. Cell. Mol. Biol. 22, 253 (2000).
26) Walter, M. J., Morton, J. D., Kajiwara, N., Agapov, E. and Holtzman, M. J. : Viral induction of a chronic asthma phenotype and genetic segregation from the acute response. J. Clin. Invest.
110, 165 (2002).
27) Young, H. W., Williams, O. W., Chandra, D., Bellinghausen, L. K., Perez, G., Suarez, A. Tuvim MJ, Roy MG, Alexander, S. N., Moghaddam, S. J., Adachi, R., Blackburn, M. R., Dickey, B. F. and Evans, C. M. : Central role of Muc5ac expression in mucous metaplasia and its regulation by conserved 5' elements. Am. J.
Respir. Cell. Mol. Biol. 37, 273 (2007).
28) Innes, A. L., Woodruff, P. G., Ferrando, R. E., Donnelly, S., Dolganov, G. M., Lazarus, S. C. and Fahy, J. V. : Epithelial mucin stores are increased in the large airways of smokers with airflow obstruction. Chest 130, 1102 (2006).
29) Fallon, M. A., Latchney, L. R., Hand, A. R., Johar, A., Denny P. A., Georgel P. T., Denny, P. C. and Culp, D. J. : The sld mutation is specific for sublingual salivary mucous cells and disrupts apomucin gene expression. Physiol. Genomics 14, 95 (2003).
30) Chen, Y., Zhao, Y. H., Kalaslavadi, T. B., Hamati, E., Nehrke, K., Le, A. D., Ann, D. K. and Wu, R. : Genome-wide search and identification of a novel gel-forming mucin MUC19/Muc19 in glandular tissues. Am. J. Respir. Cell. Mol. Biol. 30, 155 (2004).
31) Kerschner, J. E. : Mucin gene expression in human middle ear epithelium. Laryngoscope 117, 1666 (2007).
32) Yu, D. F., Chen, Y., Han, J. M., Zhang, H., Chen, X. P., Zou, W.
J., Liang, L. Y., Xu, C. C. and Liu, Z. G. : MUC19 expression in human ocular surface and lacrimal gland and its alteration in Sjogren syndrome patients. Exp. Eye Res. 86, 403 (2008).
33) Huber, H. L. and Koessler, K. K. : The pathology of bronchial asthma. Arch. Intern. Med. 30, 689 (1992).
34) Dunnill, M. S. : The pathology of asthma, with special reference to changes in the bronchial mucosa. J. Clin. Pathol. 13, 27 (1960).
35) Lethbridge-Cejku, M., Schiller, J. S. and Bernadel, L. : Summary Health Statistics for U.S Adults: National Health Interview Survey, 2002. National Center for Health Statistics (Rep. No. Vital Health Stat. 10 (222) (2004).
36) Dey, A. N. and Bloom, B. : Summary Health Statistics for U.S.
Children: National Health Interview Survey, 2003. National Center for Health Statistics (Rep. No. Vital Health Stat 10 (223)) (2005).
37) Sheehan, J. K., Richardson, P. S., Fung, D. C., Howard, M. and Thornton, D. J. : Analysis of respiratory mucus glycoproteins in asthma: a detailed study from a patient who died in status asthmaticus. Am. J. Respir. Cell. Mol. Biol. 13, 748 (1995).
38) Sheehan, J. K., Howard, M., Richardson, P. S., Longwill, T. and Thornton, D. J. : Physical characterization of a low-charge glycoform of the MUC5B mucin comprising the gel-phase of an asthmatic respiratory mucous plug. Biochem. J. 338 (Pt 2), 507 (1999).
39) Kuyper, L. M., Pare, P. D., Hogg, J. C., Lambert, R. K., Ionescu, D., Woods, R. and Bai, TR. : Characterization of airway plugging in fatal asthma. Am. J. Med. 115, 6 (2003).
40) Agrawal, A., Rengarajan, S., Adler, K. B., Ram, A., Ghosh, B., Fahim, M. and Dickey BF. : Inhibition of mucin secretion with MARCKS-related peptide improves airway obstruction in a mouse model of asthma. J. Appl. Physiol. 102, 399 (2007).
41) Fahy, J. V. : Goblet cell and mucin gene abnormalities in asthma. Chest(J). 122, 320S (2002).
42) Zhu, Y., Ehre, C., Abdullah, L. H., Sheehan, J. K., Roy, M., Evans, C. M., Dickey, B. F. and Davis, C. W. : Munc13-2-/- baseline secretion defect reveals source of oligomeric mucins in mouse airways. J. Physiol. 586, 1977 (2008).
43) Mannino, D. M., Homa, D. M., Akinbami, L. J., Ford, E. S., and Redd, S. C. : Chronic obstructive pulmonary disease surveillance—United States, 1971–2000. Respir. Care. 47, 1184 (2002).
44) Hogg, J. C., Chu, F., Utokaparch, S., Wood, R., Elliott, W. M., Buzatu, L., Cherniack, R. M., Rogers, R. M., Sciurba, F. C., Coxson, H. and Paré, P. D. : The nature of small-airway obstruction in chronic obstructive pulmonary disease. N. Engl.
J. Med. 350, 2645 (2004).
45) Prescott, E., Lange, P. and Vestbo, J. : Chronic mucus hypersecretion in COPD and death from pulmonary infection.
Eur. Respir. J. 8, 1333 (1995).
46) Vestbo, J. : Epidemiological Studies in Mucus Hypersecretion (pp. 3−12). 248 ed. (2002).
47) Lumsden, A. B., McLean, A. and Lamb, D. : Goblet and Clara cells of human distal airways: evidence for smoking induced changes in their numbers. Thorax 39, 844 (1984).
48) Caramori, G., D Gregorio,, C., Carlstedt, I., Casolari, P.,
Guzzinati, I., Adcock, I M., Barnes, P. J., Ciaccia, A., Cavallesco, G, Chung, K. F. and Papi, A. : Mucin expression in peripheral airways of patients with chronic obstructive pulmonary disease. Histopathology 45, 477 (2004).
49) Wright, J. L., Lawson, L. M., Pare, P. D., Kennedy, S., Wiggs, B. and Hogg, J. C. : The detection of small airways disease.
Am. Rev. Respir. Dis. 129, 989 (1984).
50) Kim, V., Criner, G. J., Abdallah, H. Y., Gaughan, J. P., Furukawa, S. and Solomides, C. C. : Small airway morphometry and improvement in pulmonary function after lung volume reduction surgery. Am. J. Respir. Crit. Care Med. 171, 40 (2005).
51) Cavarra, E., Martorana, P. A., Cortese, S., Gambelli, F., Di Simplicio, P. and Lungarella, G. : Neutrophils in beige mice secrete normal amounts of cathepsin G and a 46 kDa latent form of elastase that can be activated extracellularly by proteolytic activity. Biological Chemistry Hoppe-Seyler 378, 417 (1997).
52) Shao, M. X., Nakanaga, T. and Nadel, J. A. : Cigarette smoke induces MUC5AC mucin overproduction via tumor necrosis factor-alpha-converting enzyme in human airway epithelial (NCI-H292) cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 287, L420 (2004).
53) Borchers, M. T., Wesselkamper, S., Wert, S. E., Shapiro, S. D.
and Leikauf, G. D. : Monocyte inflammation augments acrolein-induced Muc5ac expression in mouse lung. Am. J.
Physiol. 277, L489 (1999).
54) Deshmukh, H. S., Case, L. M., Wesselkamper, S. C., Borchers, M. T., Martin, L. D., Shertzer, H. G., Nadel, J. A. and Leikauf, G. D. : Metalloproteinases mediate mucin 5AC expression by epidermal growth factor receptor activation. Am. J. Respir. Crit.
Care Med. 171, 305 (2005).
55) Deshmukh, H. S., Shaver, C., Case, L. M., Dietsch, M., Wesselkamper, S. C., Hardie, W. D., Korfhagen, T. R., Corradi, M., Nadel, J. A., Borchers, M.T. and Leikauf, G. D. : Acrolein- activated matrix metalloproteinase 9 contributes to persistent mucin production. Am. J. Respir. Cell. Mol. Biol. 38, 446 (2008).
56) Wilson, R. : Bacterial interactions with respiratory epithelium.
Eur. Respir. Rev. 12, 347 (2005).
57) Cash, H. A., Woods, D. E., McCullough, B., Johanson, W. G. Jr.
and Bass, J. A. : A rat model of chronic respiratory infection with Pseudomonas aeruginosa. Am. Rev. Respir. Dis. 119, 453 (1979).
58) Greenberg, S. B., Allen, M., Wilson, J. and Atmar, R. L. : Respiratory viral infections in adults with and without chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med.
162, 167 (2000).
59) Wedzicha, J. A. and Donaldson, G. C. : Exacerbations of chronic obstructive pulmonary disease. Respir. Care 48, 1204 (2003).
60) Beckham, J. D., Cadena, A., Lin, J., Piedra, P. A., Glezen, W. P., Greenberg, S, B. and Atmar, R. L. : Respiratory viral infections in patients with chronic, obstructive pulmonary disease. J.
Infect. 50, 322 (2005).
61) Falsey, A. R., Hennessey, P. A., Formica, M. A., Cox, C. and Walsh, E. E. : Respiratory syncytial virus infection in elderly and high-risk adults. N. Engl. J. Med. 352, 1749 (2005).
62) Saetta, M., Turato, G., Baraldo, S., Zanin A., Braccioni, F., Mapp, C. E., Maestrelli, P., Cavallesco, G., Papi, A. and Fabbri, L. M. : Goblet cell hyperplasia and epithelial inflammation in peripheral airways of smokers with both symptoms of chronic bronchitis and chronic airflow limitation. Am. J. Respir. Crit.
Care Med. 161, 1016 (2000).
63) Maestrelli, P., Saetta, M., Mapp, C. E. and Fabbr,i L. M. : Remodeling in response to infection and injury. Airway inflammation and hypersecretion of mucus in smoking subjects with chronic obstructive pulmonary disease. Am. J. Respir. Crit.
Care Med. 164, S76 (2001).
64) Wedzicha, J. A., Seemungal, T. A., MacCallum, P. K., Paul, E.
A., Donaldson, G. C., Bhowmik, A., Jeffries, D. J. and Meade, T. W. : Acute exacerbations of chronic obstructive pulmonary disease are accompanied by elevations of plasma fibrinogen and serum IL-6 levels. Thromb. Haemost. 84, 210 (2000).
65) Bucchioni, E., Kharitonov, S. A., Allegra, L., and Barnes, P. J.
: High levels of interleukin-6 in the exhaled breath condensate of patients with COPD. Respir. Med. 97, 1299 (2003).
66) Conn, C. A., McClellan, J. L., Maassab, H. F., Smitka, C. W., Majde, J. A. and Kluger, M. J. : Cytokines and the acute phase response to influenza virus in mice. Am. J. Physiol. 268, R78 (1995).
67) Mo, X. Y., Sarawar, S. R. and Doherty, P. C. : Induction of cytokines in mice with parainfluenza pneumonia. J. Virol. 69, 1288 (1995).
68) Zhu, Z., Tang, W., Ray, A., Wu, Y., Einarsson, O., Landry, M. L., Gwaltney, J. Jr. and Elias, J. A. : Rhinovirus stimulation of interleukin-6 in vivo and in vitro. Evidence for nuclear factor kappa B-dependent transcriptional activation. J. Clin. Invest.
97, 421 (1996).
69) Trifilieff, A., Ahmed, E. and Bertrand, C. : Time course of inflammatory and remodeling events in a murine model of asthma: effect of steroid treatment. Am. J. Physiol. Lung Cell.
Mol. Physiol. 279, L1120 (2000).
70) Tomkinson, A., Cieslewicz, G., Duez, C., Larson, K. A., Lee, J.
J., and Gelfand, E. W. : Temporal association between airway hyperresponsiveness and airway eosinophilia in ovalbumin- sensitized mice. Am. J. Respir. Crit. Care Med. 163, 721 (2001).
71) Evans, C. M., Williams, O. W., Tuvim, M. J., Nigam, R., Mixides, G. P., Blackburn, M. R., DeMayo, F. J, Burns, A. R., Smith, C., Reynolds, S. D., Stripp, B. R. and Dickey, B.F. : Mucin is
produced by Clara cells in the proximal airways of antigenchallenged mice. Am. J. Respir. Cell. Mol. Biol. 31, 382 (2004).
72) Corry, D. B., Folkesson, H. G., Warnock, M. L., Erle, D. J., Matthay, M. A., Wiener-Kronish, J. P. and Locksley, R. M. : Interleukin 4, but not interleukin 5 or eosinophils, is required in a murine model of acute airway hyperreactivity [see comments] [published erratum appears in J Exp Med 1997 May 5 185(9): 1715. J. Exp. Med. 183, 109 (1996).
73) Lee, J. J., Dimina, D., Macias, M. P., Ochkur, S. I., McGarry, M.
P., O'Neill, K. R. Protheroe C, Pero R, Nguyen T, Cormier SA, Lenkiewicz E, Colbert D, Rinaldi L, Ackerman SJ, Irvin CG.
and Lee, N. A. : Defining a link with asthma in mice congenitally deficient in eosinophils. Science 305, 1773 (2004).
74) Fulkerson, P. C., Fischetti, C. A., McBride, M. L., Hassman, L.
M., Hogan, S. P. and Rothenberg, M. E. : A central regulatory role for eosinophils and the eotaxin/CCR3 axis in chronic experimental allergic airway inflammation. Proc. Natl. Acad.
Sci. U S A. 103, 16418 (2006).
75) Davidson, D. J., Dorin, J. R., McLachlan, G., Ranaldi, V., Lamb, D., Doherty, C., Govan, J. and Porteous, D. J. : Lung disease in the cystic fibrosis mouse exposed to bacterial pathogens. Nat.
Genet. 9, 351 (1995).
76) Grunig, G., Corry, D. B., Leach, M. W., Seymour, B. W., Kurup, V. P. and Rennick, D. M. : Interleukin-10 is a natural suppressor of cytokine production and inflammation in a murine model of allergic bronchopulmonary aspergillosis. J.
Exp. Med. 185, 1089 (1997).
77) Hogaboam, C. M., Blease, K., Mehrad, B., Steinhauser, M. L., Standiford, T. J., Kunkel, S. L. and Lukacs, N. W. : Chronic airway hyperreactivity, goblet cell hyperplasia, and peribronchial fibrosis during allergic airway disease induced by Aspergillus fumigatus.
Am. J. Pathol. 156, 723 (2000).
78) Wen, F. Q., Kohyama, T., Liu, X., Zhu, Y. K., Wang, H., Kim, H.
J., Kobayashi, T., Abe, S., Spurzem, J. R. and Rennard, S. I. : Interleukin-4-and interleukin-13-enhanced transforming growth factor-beta2 production in cultured human bronchial epithelial cells is attenuated by interferon-gamma. Am. J. Respir. Cell. Mol.
Biol. 26, 484 (2002).
79) Takeyama, K, Dabbagh, K, Lee, H. M, Agustí, C, Lausier, J. A., Ueki, I. F., Grattan, K. M. and Nadel, J. A.. : Epidermal growth factor system regulates mucin production in airways. Proc.
Natl. Acad. Sci. USA. 96, 3081 (1999).
80) Vermeer, P. D., Einwalter, L. A., Moninger, T. O., Rokhlina, T., Kern J. A., Zabner, J. and Welsh, M. J. : Segregation of receptor and ligand regulates activation of epithelial growth factor receptor. Nature 422, 322 (2003).
81) Amishima, M., Munakata, M., Nasuhara, Y., Sato, A., Takahashi, T., Homma, Y. and Kawakami, Y. : Expression of epidermal growth factor and epidermal growth factor receptor