Antioxidant Activity of Manno-oligosaccharides Derived from the Hydrolysis of Polymannan by Extracellular Carbohydrase of Bacillus N3
9
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
수치
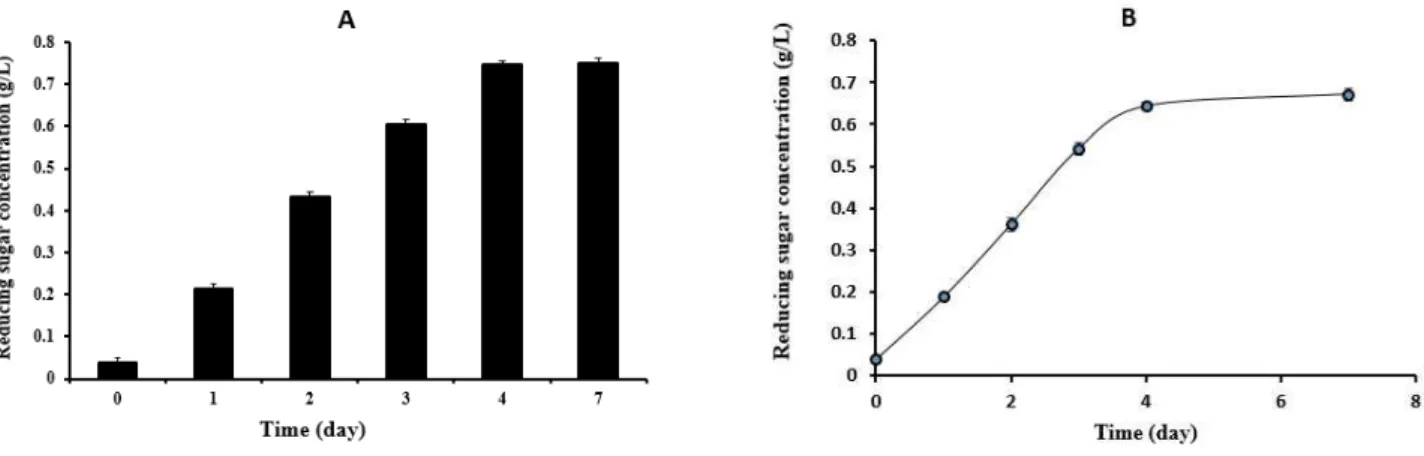
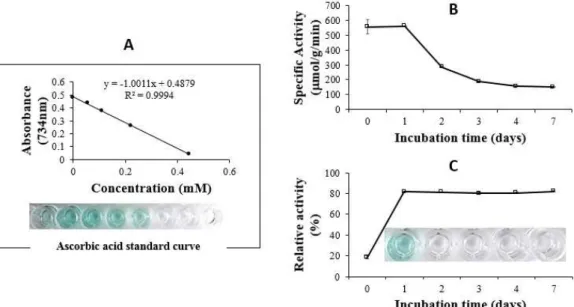

관련 문서
To measure antioxidant effects of the yuzu seed oil, DPPH radical scavenging and lipid peroxidation were investigated... DPPH radical scavenging
Hesperetin belongs to the class of flavonoids, called flavanones, which are abundant in Citrus fruits. Hesperetin is derived from the hydrolysis of its
Fresh samples were immediately tested for antioxidant activity, and antioxidant activity was compared according to gestational age and measured the difference
It considers the energy use of the different components that are involved in the distribution and viewing of video content: data centres and content delivery networks
After first field tests, we expect electric passenger drones or eVTOL aircraft (short for electric vertical take-off and landing) to start providing commercial mobility
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
The initial rate of the hydrolysis reaction is much larger than the sum of the rates of the condensation reactions so that [SiOH] equals the consumption of H 2 O by hydrolysis.
DPPH radical scavenging radical activity of jelly products added with different levels of Tribeli paprika powder.. ABTS radical scavenging radical activity of
