167 책임저자:황원덕
614-052, 부산시 진구 양정동 동의대학교 한의과대학 내과학교실 Tel: 051-850-8625, Fax: 051-853-4306 E-mail: [email protected]
접수일:2011년 5월 11일, 1차 수정일:2011년 5월 17일, 2차 수정일:2011년 5월 20일, 게재승인일:2011년 5월 28일
Correspondence to:Won Deok Hwang
Department of Internal Medicine, Dongeui University College of Oriental Medicine, Yangjeong-dong, Jin-gu, Busan 614-052, Korea
Tel: +82-51-850-8625, Fax: +82-51-853-4306 E-mail: [email protected]
활성산소종 형성을 통한 협죽도 잎 에탄올 추출물의 Caki-1 인체 신세포암 Apoptosis 유발
동의대학교 한의과대학 1내과학교실, 2생화학교실 및 3대학원 바이오물질제어학과
장자원1ㆍ김성윤2ㆍ최영현2,3ㆍ황원덕1
Induction of Apoptosis of Caki-1 Human Renal Carcinoma Cells by Ethanol Extract of Nerium indicum Leaf through Generation of Reactive Oxygen Species
Ja Won Jang1, Cheng Yun Jin2, Yung Hyun Choi2,3 and Won Deok Hwang1 Departments of 1Internal Medicine, 1Biochemistry, Dongeui University College of
Oriental Medicine, Busan 614-052, 2Department of Biomaterial Control, Graduate School and Blue-Bio Industry Regional Innovation Center, Dongeui University, Busan 614-714, Korea
Nerium indicum, an India-Pakistan-originated shrub belonging to the oleander family, is reported to possess many pharmacological activities including stimulation of cardiac muscle, anti-diabetes, anti-angiogenesis and neuroprotection activities. However, the cellular and molecular mechanisms of apoptosis induction by N. indicum extract in human cancer cells are poorly understood. In the present study, it was investigated the biochemical mechanisms of the pro-apoptotic effects by ethanol extract of N. indicum leaf (ENIL) in Caki-1 human renal carcinoma cells. It was found that ENIL could inhibit the cell proliferation of Caki-1 cells in a concentration- and time-dependent manner, which was associated with the induction of apoptosis as determined using DAPI staining and flow cytometric analysis. The induction of apoptotic cell death by ENIL was connected with a loss of mitochondrial membrane potential (MMP), proteolytic activation of caspase-3 and -9, and degradation of poly (ADP-ribose) polymerase (PARP) in Caki-1 cells. However, ENIL did not significantly affect the activity of caspase-8 and the levels of the inhibitor of apoptosis protein (IAP) family members. In addition, following exposure to ENIL, the production of reactive oxygen species (ROS) were increased, and inhibition of ROS generation by ROS scavenger, N-acetylcysteine (NAC), attenuated ENIL-induced apoptosis. Taken together, these findings suggest that ENIL may be a potential chemotherapeutic agent for use in control of Caki-1 human renal carcinoma cells and further studies will be needed for identification of active compounds that confer the anti-cancer activity of ENIL. (Cancer Prev Res 16, 167-179, 2011)
Key Words: Nerium indicum, Apoptosis, ROS, Caki-1
서 론
신장 세포암은 원발성 신종양의 약 85%를 차지하는 악성종양으로 약 25∼30%에서 진단 시 이미 전이가 일 어난 것으로 알려져 있다. 신장에 국한된 신세포암의 경 우는 근치적 신적출술로 약 60∼70%의 환자에서 5년 생 존율을 기대할 수 있으나 전이 신세포암의 경우는 현재 까지 뚜렷한 치료법이 없는 것으로 알려져 있다.1) 현재 까지 vinblastine이 유용한 화학요법제로 알려져 있으나 반응률은 10% 내외가 고작이며 전이 신세포암은 대개 급속히 진행하여 대부분이 2년 이내에 사망한다.2,3) 따라 서 신세포암의 경우에 화학치료가 왜 효과가 없는지에 대한 원인 분석과 함께 특별한 치료법이 없는 전이 신세 포암의 생존을 억제시키기 위해서는 이러한 세포들의 사멸을 촉진할 수 있는 약물개발이 시급하다.
유도화(柳桃花)라고도 불리는 협죽도(Nerium indicum, 夾 竹桃)는 협죽도과에 속하는 넓은 잎 늘푸른떨기나무로 서, 인도가 원산지로서 한국의 제주도에 자생하며,4) 특 히 파키스탄 지역에서는 제2형 당뇨병 치료를 위한 민간 요법으로 사용되고 있다.5) 기존의 연구에 따르면 협죽도 에는 여러 종류의 cardiac glycoside, alkaloid 및 carbohydrate 가 함유되어 있어 신생혈관형성 억제, 항암효과 및 신경 세포 보호작용 등과 같은 약리학적인 효과를 가지는 것 으로 보고되어지고 있다.6∼8) 또한 협죽도의 잎에서 분리 된 arabinogalactan은 mitogen에 의해 유도된 T 및 B lym- phocyte의 증식을 자극하며, 꽃에서 분리된 rhamnoga- lacturonan 및 xyloglucan은 PC12 세포의 증식 및 분화를 유도하는 것으로 보고된 바 있다.5,7) 최근의 연구결과에 의하면 낮은 농도의 협죽도 추출물이 암세포의 항전이 효과와 더불어 심장마비와 뇌졸중을 포함한 심혈관계 질환에 탁월한 보호 효과를 가지고 있다고 알려져 있
다.9,10) 또한, 유방암 세포의 세포사멸(apoptosis)을 유도하
여 암세포의 성장을 억제한 반면 마우스에서 유래된 정 상적인 유방세포에는 독성이 없는 것으로 보고되었다.11) 따라서 협죽도의 다양한 질병 예방과 치료에 효과적일 것으로 추정되지만 현재까지 apoptosis 유도와 관련된 생 화학적 기전에 대한 연구는 미비한 실정이다.
인체 내에서는 지속적으로 일어나는 세포의 죽음 기 전으로는 apoptosis와 necrosis가 있으며, 후자는 면역반응 을 활성화시키지만 전자는 그렇지 않다.12,13) Necrosis는 감염이나 조직손상에 의하여 발생하는 비정상적인 과정 이므로 인체의 면역계는 apoptosis에 대해서는 관용 (tolerance)을 보이지만 necrosis에 대해서는 반응을 나타낸
다. Apoptosis는 programmed cell death라고 불리는 생리학 적 과정이며 mitochondria의 기능 변화, 세포의 수축, 염색 질 응축, 핵의 단편화 현상, 세포막의 수포화 현상, cas- pases 활성화, 세포 표면에 phosphatidylserine의 발현 및 apoptotic body의 형성 등과 같은 형태적 또는 생화학적인 변화를 동반하고, 세포의 종류와 손상된 형태에 따라 여 러 유전자의 증가 또는 감소 등과 같은 상호작용에 의해 유발된다.14,15) 또한 apoptosis는 대부분의 조직에서 손상 을 입었거나 감염된 세포들을 제거하는 중요한 과정으 로서 이러한 apoptosis 과정이 실패하게 되면 암과 같은 여러 가지 질병의 원인이 된다.15) 따라서 apoptosis는 암 화 과정의 여러 단계에서 암을 치료하는 중요한 표적이 되고 있다.
포유동물에서 apoptosis의 조절에 관여하는 유전자로서 암유발유전자(proto-oncogene)인 B cell lymphoma-2 (Bcl-2) 가 발견된 이후 Bcl-2와 구조적으로 유사한 많은 단백질 들이 밝혀짐으로서 거대한 Bcl-2 family가 구성되었다.16) 지금까지의 연구결과를 바탕으로 이들 Bcl-2 family의 종 류를 살펴보면, apoptosis 유도에 관여하는 것으로 알려진 Bad, Bax, Bid, Bik 및 Bim 등과 같은 pro-apoptotic 유전자 와, 세포의 생존에 관여하는 것으로 알려진 Bcl-2, Bcl-xL 및 Bcl-wL 등과 같은 anti-apoptotic 유전자가 보고되었다.
Mitochondria에서 pro-apoptotic Bcl-2 family는 cytochrom c를 세포질로 방출시킴으로서 caspase를 활성화시키는데, 이 과정은 활성 산소종(reactive oxygen species, ROS) 생산에 의존 및 비의존적으로 나타날 수 있다.17,18) 일반적으로 ROS는 인체의 건강에 해로운 것으로 알려져 있지만 apoptosis 유발의 경우에 면역계의 과잉반응에 의하여 건 강한 세포가 파괴되는 것을 막아주는 역할을 한다.
본 연구에서는 협죽도 추출물이 인체 신세포암인 Caki-1 세포에 미치는 항암 효과의 생화학적 기전의 해 석을 위하여 암세포의 증식에 미치는 협죽도 잎의 에탄 올 추출물의 영향을 조사하였고, apoptosis 과정 중 caspase 활성 및 mitochondria에 의한 ROS의 생성에 관여하는 몇 가지 중요한 유전자들의 발현 변화를 조사하여 유의적 인 결과를 얻었기에 이를 보고하는 바이다.
재료 및 방법 1. 실험재료
본 실험에 사용된 협죽도는 부산광역시 영도구에서 채집된 것을 동정하여 사용하였다. 협죽도 잎의 에탄올 추출물(ethanol extract of N. indicum leaf, ENIL)을 얻기 위하 여 채집된 협죽도의 잎을 흐르는 물로 충분히 세척하고
건조시킨 후 잘게 분쇄하였다. 건조된 협죽도의 잎 100 g에 ethanol 1 l를 첨가하여 60oC, 150 rpm으로 3일간 교반 후 상층액만 분리하여 Whatman 필터(No. 2)로 걸러내고 감압 농축과정을 통하여 고형성분을 얻어내어 막자사발 로 잘게 마쇄하고 밀봉시켜 −70oC 초저온 냉동고에 보 관하였으며, 실험 시에는 dimethyl sulfoxide (DMSO)를 이 용하여 10 mg/ml의 농도로 만든 다음 이를 적정 농도로 배지에 희석하여 처리하였다.
2. 세포의 배양
실험에 사용한 Caki-1 신세포암세포는 생명공학연구 소(KRIBB, Taejeon, Korea)에서 분양 받았으며 90%의 RPMI-1640 배지(Gibco BRL, Grand Island, NY, USA), 10%
fetal bovine serum (FBS, Gibco BRL)에 1%의 penicillin 및 streptomycin (Gibco BRL)이 포함된 성장배지를 사용하여 5% CO2, 37oC의 조건하에서 배양하였다. 배지는 매 48시 간마다 교환해주었고, 세포 수의 증식에 따른 과밀도 현 상을 해소하기 위하여 0.05% trypsin-ethylenediamine tetra- acetic acid (EDTA, Gibco BRL)를 처리하여 세포를 부유시 킨 다음 적정수의 세포를 분주하여 재배양하였다.
3. MTT assay에 의한 세포 성장억제 조사
세포 배양용 6 well plate에 Caki-1 세포를 3×104개/ml로 분주하여 안정화시킨 다음 ENIL을 각각 배지에 희석하 여 각 well 당 적정농도 혹은 시간별로 처리하였다. 일정 시간 경과 후 0.5 mg/ml 농도의 tetrazolium bromide salt (MTT, Amresco, Solon, OH, USA)를 2 ml씩 분주하고 2시 간 동안 CO2 incubator에서 배양시킨 다음 MTT 시약을 깨끗하게 제거하고 DMSO를 1 ml씩 분주하여 well에 생 성된 formazin을 모두 녹인 후 96 well plate에 200 μl씩 옮겨서 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)로 540 nm에서 흡광도를 측정하였다. 측정은 모두 세 번을 하였으며, 그에 대한 평균값과 표준 오차를 Microsoft EXCEL program을 사용하여 분석하였다.
4. 위상차 현미경을 이용한 세포의 성장과 형태의 관찰
세포 배양용 100 mm petri dishes에 세포를 3×104개/ml 정도로 분주하여 24시간 동안 안정화시킨 다음 ENIL을 적정농도로 희석 처리하여 24시간 동안 배양한 후, 위상 차 현미경(Carl Zeiss, Germany)을 이용하여 200배의 배율 로 각 농도에 따른 형태의 변화를 관찰하였다.
5. DAPI staining에 의한 세포핵의 형태 관찰 ENIL 처리에 의한 암세포의 apoptosis 유발 여부 확인
을 위한 핵의 형태적 변화를 관찰하기 위하여 준비된 세 포를 모은 다음 37% formaldehyde 용액과 PBS를 1:9의 비율로 섞은 fixing solution을 모아진 세포에 500 μl 첨가 하여 충분히 섞은 후, 상온에서 10분 동안 고정하였다.
2,000 rpm으로 5분간 원심 분리한 후 상층액을 제거하고 PBS 200 μl를 넣어서 충분히 섞은 후, slide glass 위에 80 μl 정도 떨어뜨려 1,000 rpm에서 5분간 cytospin하였다.
PBS로 2∼3회 washing하고 PBS가 다 마르기 전에 0.2%의 Triton X-100 (Amresco)을 첨가하여 상온에서 10분간 고정 하였다. 그 후 다시 PBS로 washing하고 4',6-diamidino-2- phenylindole (DAPI, Sigma, St. Luis, MO, USA) 용액을 세포 가 고정된 slide glass 위에 적당량을 떨어뜨린 후 빛을 차 단하고 상온에서 염색시켰다. 15분 정도 염색시킨 후, PBS로 DAPI 용액을 충분하게 세척하고 증류수로 재빨 리 세척한 다음 absolute alcohol을 이용하여 탈수과정을 거친 slide glass 위에 mounting solution을 처리한 후 형광 현미경(Carl Zeiss)을 이용하여 400배의 배율로 각 농도에 따른 암세포의 핵의 형태 변화를 관찰하였다.
6. Flow cytometry 분석
정상 및 ENIL이 함유된 배지에서 24시간 동안 배양시 킨 암세포들을 씻어 내고 0.05% trypsin-EDTA를 처리하 여 부유시킨 다음 2,000 rpm으로 5분간 원심 분리하여 상층액을 버리고 세포들만 모았다. 여기에 다시 PBS를 첨가하여 충분히 씻은 다음 2,000 rpm으로 5분간 원심분 리 한 후 상층액만 버리고 남은 세포에 CycleTEST PLUS DNA REAGENT Kit (Becton Dickinson, San Jose, CA, USA) 를 이용하여 고정 및 염색을 하여 4oC, 암실에서 30분 동 안 반응을 시킨 다음 DNA flow cytometry (Becton Dickin- son)에 적용시켜 형광반응에 따른 histogram을 ModiFit LT (Becton Dickinson) 프로그램으로 분석하였다.
7. Rverse transcriptase polymerase chain reac- tion (RT-PCR)에 의한 mRNA 발현 변화의 분석
동일한 조건에서 준비된 암세포를 대상으로 TRIzol re- agent (Invitrogen Co., Carlsbad, CA, USA)를 이용하여 total RNA를 분리하였다. 분리된 RNA를 정량한 후, 각각의 primer, DEPC water 그리고 ONE-STEP RT-PCR PreMix Kit (Intron, Korea)를 넣고 Mastercycler gradient (Eppendorf, Hamburg, Germany)를 이용하여 증폭하였다. 각 PCR 산물 들의 양적 차이를 확인하기 위하여 1X TAE buffer로 1%
agarose gel을 만들고 well당 각각의 primer에 해당하는 PCR 산물에 DNA gel loading solution을 섞어서 loading 한 후 100 V에서 전기영동을 하였다. 전기영동으로 DNA 분
리가 끝난 gel을 ethidium bromide (EtBr, Sigma)를 이용하 여 염색한 후 UV 하에서 확인하였으며, housekeeping 유 전자인 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) 를 internal control로 사용하였다.
8. Western blot analysis에 의한 단백질 발현 변화의 분석
정상 및 ENIL이 처리된 배지에서 자란 세포들을 PBS 로 씻어 내고 0.05% trypsin-EDTA를 처리하여 부유시킨 다음 원심분리를 하여 세포를 모았다. 이렇게 모아진 세 포에 적당량의 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenymethyl- sulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]를 첨가 하여 4oC에서 1시간 동안 반응시킨 후, 14,000 rpm으로 30분간 원심분리하여 그 상층액을 취하였다. 상층액의 단백질 농도는 Bio-Rad 단백질 정량 시약(Bio-Rad, Hercules, CA, USA)과 그 사용방법에 따라 정량한 다음 동량의 Laemmli sample buffer (Bio-Rad)를 섞어서 sample을 만들었 다. 이렇게 만든 동량의 단백질을 sodium dodecyl sulphate (SDS)-polyacrylamide gel을 이용하여 전기영동으로 분리하 였다. 분리된 단백질을 함유한 acrylamide gel을 nitro- cellulose membrane (Schleicher and Schuell, Keene, NH, USA) 으로 electroblotting에 의해 전이시킨 후, 5% skim milk를 함유한 PBS-T (0.1% Tween 20 in PBS)에 담구어 상온에서 1시간 정도 incubation하여 비특이적인 단백질들에 대한 blocking을 실시하고 PBS-T로 15분 정도 세척하였다. 준 비된 membrane에 1차 antibody를 처리하여 상온에서 2시 간 이상 또는 4oC에서 over night 시킨 다음 PBS-T로 세척 하고 처리된 1차 antibody에 맞는 2차 antibody를 사용하 여 상온에서 1시간 정도 반응시켰다. 다시 PBS-T로 세척 하고 Enhanced Chemiluminoesence (ECL) 용액(Amersham Life Science Corp., Arlington Heights, IL, USA)을 적용시킨 다음 암실에서 X-ray film에 감광시켜 특정단백질의 양을 분석하였다.
9. In vitro caspase 활성 변화 측정
ENIL 처리에 의한 caspase의 활성화 정도를 알아보기 위하여 정상 및 ENIL이 처리된 배지에서 24시간 배양된 세포를 모은 뒤 단백질을 추출하고 정량하여 각각 150 μg의 단백질을 fluorogenic peptide 기질 100 μM이 함유 된 extraction buffer [40 mM HEPES (pH 7.4), 20% glycerol (v/v), 1 mM EDTA, 0.2% NP-40 and 10 mM DL-DTT] 50 μl에 혼합하였으며, microtiter plate에 다시 extraction buf- fer에 희석하여 각 sample당 총 volume이 100 μl가 되게
하였다. 실험에 사용된 기질은 caspase-3의 경우에는 Asp- Glu-Val-Asp (DEVD)-p-nitroaniline (pNA)이었고, caspase-8 의 경우에는 Ile-Glu-Thr-Asp (IETD)-pNA이었으며, cas- pase-9는 Leu-Glu-His-Asp (LEHD)-pNA였다. 준비된 plate 를 37oC에서 3시간동안 incubation 시킨 후 ELISA reader를 이용하여 405 nm의 흡광도를 이용하여 반응의 정도를 측정하였다.
10. ROS 생성 변화 측정
6 well plate에 Caki-1 세포를 3×104개/ml로 분주하고, ENIL 80 μg/ml을 각각 제시된 시간동안 처리한 후, 배양 액에 5-(and 6)-carboxy-2’7’-dichloro-dihydrofluorescein diac- etate (DCFDA, sigma)를 10 μM 농도가 되게 분주하여 20 분간 37oC, 5% CO2 조건에서 배양시켰다. 그 후 세포들 을 모아 3,000 rpm에서 원심분리하여 상층액을 제거한 후 PBS로 세척하고 DNA flow cytometer를 이용하여 상대 적인 reactive oxygen species (ROS)의 생성 변화를 관찰하였 다.
결 과
1. 암세포의 성장에 미치는 ENIL의 영향
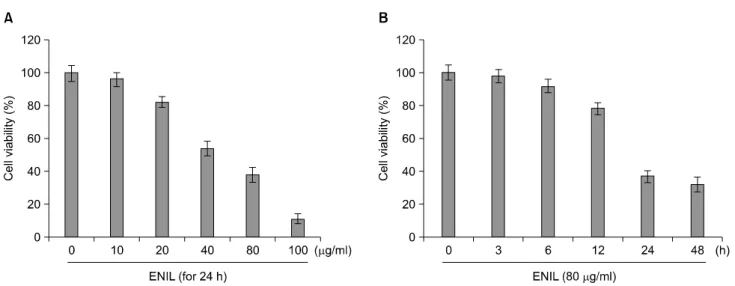
ENIL의 처리에 따른 Caki-1 세포의 성장억제 정도를 알아보기 위하여 ENIL을 적정농도 및 적정 시간 동안 처 리한 후, MTT assay를 이용하여 조사한 결과는 Fig. 1에 나타낸 바와 같다. 결과에서 알 수 있듯이 Caki-1 세포에 0∼100 μg/ml의 ENIL를 24시간 처리 시 농도 의존적으 로 세포의 성장이 억제되는 것으로 나타났다(Fig. 1A). 또 한 ENIL의 농도를 80 μg/ml로 고정하고 0∼48시간까지 처리하였을 경우 12시간 처리군까지는 큰 변화가 나타 나지 않았지만 24시간 처리군에서부터 급격한 생존율의 감소가 관찰되어 48시간 처리군에서는 60% 이상의 생존 율 감소 현상이 나타나는 것으로 조사되었다(Fig. 1B). 이 상의 결과에서 Caki-1 세포에 ENIL을 처리하였을 경우 처리시간에 따른 생존율 감소현상에 비교하여 처리농도 에 따른 생존율 감소현상이 민감하게 나타나는 것으로 나타났으므로 이후의 실험은 농도 의존적으로 ENIL를 처리하여 진행하였다.
2. ENIL를 처리에 의한 apoptosis의 유발
ENIL 처리에 의하여 유발되는 암세포의 전체적인 형 태변화를 알아보기 위하여 Caki-1 세포에 다양한 농도의 ENIL을 처리하여 24시간 동안 배양한 후 도립 현미경을 이용하여 관찰하였다. Fig. 2A에서 볼 수 있듯이 ENIL의
Fig. 1. Effects of ENIL on the cell viability of Caki-1 human renal carcinoma cells. Cells were seeded at 3×104/ml in a 24-well plate and incubated for 24 h. The cells were treated with the indicated concentrations of fucoidan for 24 h (A) or 80 μg/ml of ENIL for the indicated times (B). The cell viability was measured by the metabolic-dye-based MTT assay. Each point represents the mean±SD of three independent experiments.
처리 농도가 증가할수록 암세포의 밀도가 감소하면서 길고 분지를 형성하는 듯한 dendrite-like한 구조로 바뀌 었으며, 부착력을 상실하고 배지 위로 부유되는 경향성 이 증가하는 것과 같은 심한 형태적 변화가 관찰되었다.
다음으로 Caki-1 세포의 형태변화가 apoptosis 유발과 연 관성이 있는지 알아보기 위하여 정상 및 ENIL이 처리된 배지에서 배양된 암세포를 고정시킨 후 DAPI 염색을 실 시하여 핵의 형태변화를 형광현미경 하에서 관찰하였 다. Fig. 2B의 결과에서 알 수 있듯이 정상배지의 경우 핵의 형태에 아무런 변화가 나타나지 않았지만 ENIL를 처리한 농도가 높아짐에 따라 apoptosis가 일어난 세포에 서 전형적으로 관찰되는 염색질 응축에 의한 apoptotic body가 관찰되었다. 따라서 ENIL 처리에 따른 apoptosis 유발의 정도를 정량적으로 비교 평가하기 위하여 상기 와 동일한 조건으로 배양된 세포들을 대상으로 DNA flow cytometry 분석을 통한 세포주기의 sub-G1기에 해당 되는 세포들의 빈도를 조사한 결과, ENIL 처리 농도의 증가에 따라 sub-G1기에 해당하는 세포의 빈도가 현저 히 증가함을 알 수 있었다(Fig. 2C). 특히 20 μg/ml의 ENIL을 처리하였을 경우 sub-G1기에 해당하는 세포의 빈도가 소폭 증가하였으나 40 μg/ml 처리군에서는 약 19% 정도로 증가하였으며 80 μg/ml 처리군에서는 약 32% 정도로 급격하게 증가하는 것으로 관찰되었다. 이 상의 결과를 살펴볼 때 ENIL 처리에 의한 Caki-1 인체신 세포암의 생존율 감소는 apoptosis 유발과 밀접한 관련이 있음을 알 수 있었다.
3. Bcl-2 family의 발현 및 mitochondrial mem- brane potential (MMP)에 미치는 ENIL의 영향
다음으로 Bcl-2 family가 ENIL에 의해 유발되는 apopto- sis에 있어서 어떠한 영향을 미치는지를 확인하였다. Fig.
3A 및 B에 나타낸 바와 같이 Caki-1 세포에 ENIL을 처리 하였을 경우 anti-apoptotic 유전자인 Bcl-2 및 Bcl-xL 유전 자의 발현이 전사 및 번역수준 모두에서 감소되는 것으 로 나타났으며, pro-apoptotic 유전자인 Bax의 경우에는 전사 및 번역수준 모두에서 증가되는 것으로 나타났다.
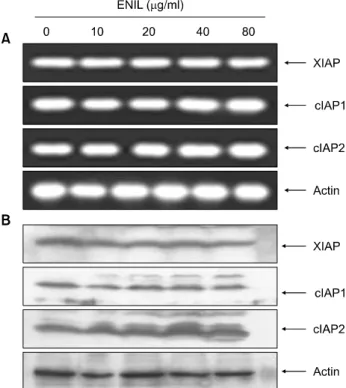
하지만 또 다른 pro-apoptotic 유전자인 Bad의 경우에는 큰 변화가 관찰되지 않았다. 또한 mitochondrial 외막에 존재하는 Bcl-2 family의 발현 변화에 의하여 유발되는 것 으로 알려진 MMP의 loss가 ENIL 처리에 의하여 현저하 게 증가되어(Fig. 3C), ENIL 처리에 따른 apoptosis 유발에 있어서 Bcl-2 family가 중요한 역할을 한다는 것을 알 수 있었다. 그러나 caspase의 활성을 억제하는 것으로 알려 진 inhibitors of apoptosis (IAP) family가 ENIL 처리에 의하 여 유발되는 apoptosis에 어떠한 영향을 미치는 지를 조사 한 결과, ENIL 처리에 의한 IAP family에 속하는 유전자 의 발현의 변화는 전사와 번역수준 모두에서 관찰되지 않았다.
4. Caspase의 발현 및 활성에 미치는 ENIL의 영향
Apoptosis 유발에 있어서 중요한 역할을 하는 것으로 알려진 caspase-3, -8 및 -9의 발현 및 활성에 미치는 ENIL 의 영향을 조사하였다. Fig. 5A 및 B에 나타난 바와 같이
Fig. 2. Induction of apoptosis by ENIL treatment in Caki-1 cells. Cells were incubated with the indicated concentrations of ENIL for 24 h. (A) Cell morphology was visualized by light microscopy (original magnification, ×200). (B) The cells were fixed and stained with DAPI solution. After 10 min incubation at room temperature, stained nuclei were then observed under a fluorescent microscope (original magnification, ×400). (C) To quantify the degree of apoptosis induced by ENIL, cells grown under the same conditions as (A) were evaluated by flow cytometry for sub-G1 DNA content, which represents the cells undergoing apoptotic DNA degradation. Data are the mean±SD of two different experiments.
ENIL의 처리에 의하여 initiator caspase로 알려진 caspase-9 와 effector caspase로 알려진 caspase-3의 활성형 단백질은 나타나지 않았지만 전체 단백질의 발현이 처리농도 의 존적으로 현저히 감소되는 것으로 관찰되었으며, cas- pase-3의 기질단백질로서 DNA repair와 genomic stability에 관여하는 poly (ADP-ribose) polymerase (PARP)의 단편화 현 상이 관찰되었다. 하지만 또 다른 initiator caspase인 cas- pase-8의 경우는 ENIL 처리에 따른 변화가 관찰되지 않
았다. 이상의 Western blotting에 의한 결과를 재확인하기 위하여 caspases의 활성 정도를 직접 분석한 결과, ENIL 처리 농도 증가에 따라 caspase-3이 현저히 증가되었고 caspase-9의 활성은 조금 증가된 반면, caspase-8의 경우에 는 큰 변화가 없었음을 알 수 있었다. 이상의 결과를 살 펴볼 때 ENIL 처리에 따른 Caki-1 세포에서의 apoptosis 유발은 caspase-8과 무관하게 caspase-3 및 -9의 활성화가 중요한 역할을 한다는 것을 알 수 있었다.
Fig. 3. Effects of ENIL on levels of Bcl-2 family proteins and MMP values in Caki-1 cells. (A) After 24 h incubation with the indicated concentrations of ENIL, total RNAs were isolated and reverse-transcribed. The resulting cDNAs were subjected to PCR with the indicated primers and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining.
GAPDH was used as an internal control. (B) The cells were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (C) The cells grown under the same conditions as (A) were collected and incubated with JC-1 (10 μM) for 20 min at 37oC in the dark. The cells were the washed once with PBS and analyzed by a DNA flow cytometer. The results are expressed as the mean±SD of three independent experiments.
5. JNK의 발현 및 ROS의 생성에 미치는 ENIL의 영향
마지막으로 ENIL의 처리에 따른 apoptosis의 유발에 있 어서 mitogen-activated protein kinases (MAPKs)의 한 종류 로서 스트레스와 연관된 유전자인 c-Jun NH(2) -terminal kinase (JNK)와의 연관성을 알아본 결과, Fig. 6A에 나타 난 바와 같이 ENIL이 처리된 Caki-1 세포에서 JNK의 활 성형인 phosphoryated JNK (p-JNK) 발현양이 현저히 증가 되었다. 따라서 ENIL에 의하여 유발되는 apoptosis에 JNK 경로가 관여하는지를 확인하기 위하여 JNK inhibitor인 SP600125를 1시간 선처리한 후 ENIL을 처리한 다음 DNA flow cytometry를 이용하여 apoptosis의 유발 정도를
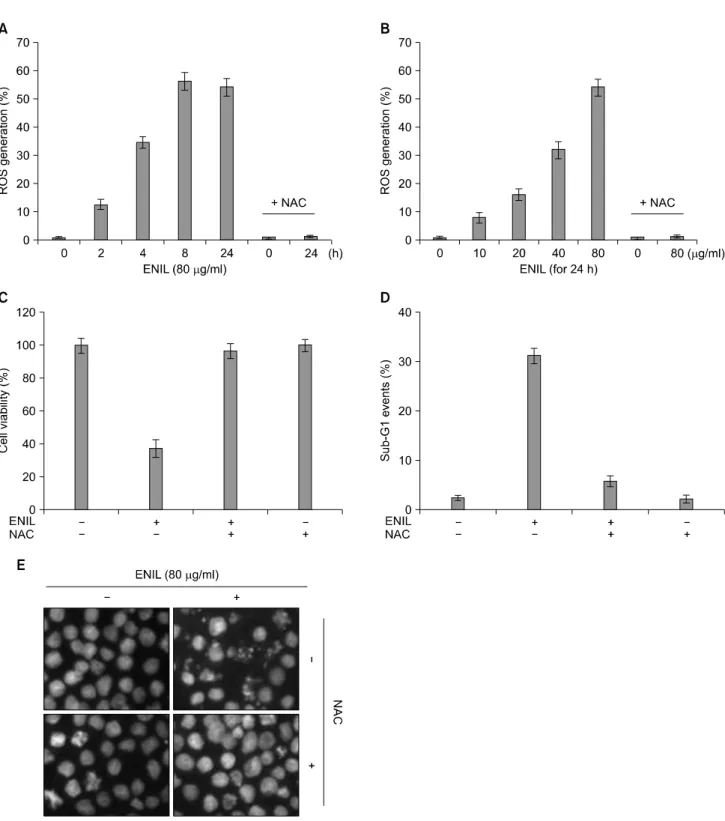
정량적으로 분석한 결과, sub-G1기의 비율이 ENIL 단독 처리군과 비교하여 큰 차이가 없는 것으로 나타났으며 (Fig. 6B), DAPI 염색을 통한 결과에서도 ENIL에 의한 JNK의 활성증가는 apoptosis 유발에 직접적인 연관성이 없음을 알 수 있었다(Fig. 6C). 다음으로 ENIL의 처리에 따른 ROS 생성 및 apoptosis 유도와의 연관성을 확인해보 았다. Fig. 7A 및 B에 나타난 바와 같이 Caki-1 세포에 80 μg/ml의 ENIL을 처리하였을 경우 2시간부터 8시간까지 ROS의 생성이 현저히 증가되었고 24시간 이후 감소하였 고, ENIL 처리에 의한 ROS 생성은 ENIL 처리 농도 의존 적으로 증가되었으며, 이러한 ROS의 생성은 ROS scav- enger인 N-acetylcysteine (NAC)에 의하여 완벽하게 억제되
Fig. 4. Effects of ENIL on levels of IAP family proteins in Caki-1 cells. (A) After 24 h incubation with the indicated concentra- tions of ENIL, total RNAs were isolated and reverse-trans- cribed. The resulting cDNAs were subjected to PCR with the indicated primers and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells grown under the same conditions as (A) were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The mem- branes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
었다. 따라서 ROS의 생성이 ENIL에 의하여 유발되는 apoptosis에 직접 관여하는지를 확인하기 위하여 NAC을 1시간 선처리 한 후 ENIL을 처리한 다음 apoptosis의 유발 정도를 정량적으로 분석하였다. Fig. 6C∼E의 결과에서 알 수 있듯이 ENIL에 의하여 유발되었던 apoptosis가 ROS scavenger인 NAC 선처리에 의하여 완벽하게 억제되었으 며, 이에 따른 생존율도 정상수준으로 회복되는 것으로 나타났다. 이상의 결과에서 Caki-1 세포에서 ENIL 처리 에 의한 apoptosis 유발은 ROS 생성을 통하여 일어난다는 것을 알 수 있었다.
고 찰
본 연구에서는 인체 신세포암인 Caki-1 세포의 증식에 미치는 ENIL의 영향과 이와 연관된 apoptosis 유발 여부 를 조사하였다. 이를 위하여 Caki-1 세포에서 ENIL 처리 농도 및 시간에 따른 생존율의 정도를 비교한 결과, Caki-1 세포에서 ENIL의 처리농도 및 시간이 증가할수록 생존율이 감소하는 것을 확인할 수 있었으며(Fig. 1), Fig.
2A에서와 같이 이러한 생존율의 감소는 세포의 형태변 화와도 연관이 있었다. ENIL에 의한 생존율의 감소 및 형태변화가 apoptosis 유발과 관련이 있는지를 확인하기 위하여 핵의 형태 및 sub-G1의 정략적인 분석을 실시한 결과, Fig. 2B 및 C에서 나타난 바와 같이 ENIL 처리에 의하여 apoptosis가 유발된 세포에서 특이적으로 나타나 는 염색질 응축에 의한 apoptotic body의 증가가 관찰되었 고, sub-G1의 비율이 농도의존적으로 현저하게 증가하는 것으로 나타났다. 이는 Caki-1 세포에서 ENIL이 유발하 는 생존율의 감소는 apoptosis의 유발과 밀접한 관계가 있 음을 의미한다.
일반적으로 apoptosis 과정은 Bcl-2 및 IAP family 인자들 및 caspase 등과 같은 여러 가지 유전자들이 관여하는 것 으로 알려져 있다. 먼저 Bcl-2 family에 속하는 단백질들 은 네 가지의 Bcl homology (BH) domains (BH1-BH4) 중 최소한 한 개의 domain을 포함하고 있으며, mitochondria 보존과 mitochondria에 의해 유도되는 apoptosis를 조절하 는 중요한 인자들이다.19) 이 단백질들은 Bcl-2, Bcl-XL, Bcl-w 및 Mcl-1과 같이 apoptosis를 억제하는 anti-apoptotic member와 Bax, Bad, Bak, Bid 및 Bcl-XS과 같이 apoptosis를 유발하는 pro-apoptotic member로 구성되어 있다.20) 이러 한 anti-apoptotic 및 pro-apoptotic member들 사이의 비율에 의하여 생리적 및 병리적인 조건하에서 세포의 생존 및 죽음이 결정되는데,21) 이 member들 간의 균형이 깨어지 게 되면 mitochondria로부터 cytosol로 cytochrome c가 방출
되어 cysteine-related proteases인 caspase, 종양억제 유전자 인 p53, DNA의 단편화와 연관된 endonuclease 등의 활성 을 증가시켜 apoptosis가 유발되는 것으로 알려져 있다.22) 따라서 본 실험에서 사용한 Caki-1 세포에 ENIL을 처리 하였을 경우 Bcl-2 family에 속하는 인자들의 발현 변화가 유발되는지를 먼저 확인한 결과, Fig. 3에 나타난 바와 같이 ENIL에 의해 Caki-1 세포에서는 pro-apoptotic 인자 인 Bax의 발현 증가와 함께 anti-apoptotic 인자인 Bcl-2 및 Bcl-XL의 전사 및 번역수준에서의 발현감소가 관찰되었 으며 이로 인하여 MMP의 loss가 증가하는 것으로 확인되 었다. 이상의 결과를 살펴볼 때 ENIL에 의한 apoptosis 유 발은 Bcl-2 family에 속하는 유전자의 발현 차이에 의한 mitochondria membrane의 permeability 변화에 의하여 cyto- chrome c가 mitochondria에서 cytosol로 방출되면서 개시하
Fig. 5. Activation of caspases and degradation of PARP protein by ENIL in Caki-1 cells. (A) Caki-1cells were treated with the indicated concentrations of ENIL for 24 h. The cells were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (B) After 24 h incubation with the indicated concentrations of ENIL, the cells were lysed and aliquots (50 mg protein) were assayed for in vitro caspase-3, -8, and -9 activity using DEVD-pNA, IETD-pNA, and LEHD-pNA as substrates, respectively, at 37oC for 1 h. The released fluorescent products were measured. Data are expressed as mean±SD of three independent experiments.
Fig. 6. Effects of JNK activation on ENIL-induced apoptosis in Caki-1 cells. (A) Cells were treated with the indicated concentrations of ENIL for 24 h. Cells were then lysed, and equal amounts of cell lysates were resolved by SDS-polyacrylamide gels, transferred to nitrocellulose, and probed with anti-p-JNK antibody. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (B) Cells were pretreated with SP600125 (40 mM) for 1 h and then treated with ENIL (80 μg/ml) for 24 h. The percentage of sub-G1 cell population was evaluated by a flow cytometer. Each point represents the mean±SD of three independent experiments. (C) The cells grown under the same conditions as (B) were fixed and stained with DAPI solution. After 10 min incubation at room temperature, stained nuclei were then observed under a fluorescent microscope (original magnification,
×400).
Fig. 7. ENIL-induced apoptosis is associated with ROS generation in Caki-1 cells. (A, B) Cells were treated with or without NAC (10 mM) for 2 h before being challenged with 80 μg/ml of Caki-1 for the indicated times (A) or with the indicated concentrations of ENIL for 24 h (B). For the measurement of ROS generation, the cells were incubated with 10 μM DCF-DA at 37oC for 30 min, and ROS generation was measured by flow cytometer. (C) Cells were treated with or without NAC for 1 h before being challenged with 80 μg/ml of ENIL for 24 h. The cell viability was measured by the metabolic-dye-based MTT assay. (D) Following incubation of cells under the same conditions as (C), the degree of apoptosis were evaluated by flow cytometry for sub-G1 DNA content, which represents the cells undergoing apoptotic DNA degradation. Data are the mean±SD of two different experiments.
(E) The cells grown under the same conditions as (C) were fixed and stained with DAPI solution. After 10 min incubation at room temperature, stained nuclei were then observed under a fluorescent microscope (original magnification, ×400).
는 것으로 생각된다.
Caspase 활성 의존적인 apoptosis는 여러 단계로 조절되 는데 그 중 IAP family는 caspases의 활성을 직․간접적으로 억제하는 것으로 알려져 있다.23,24) 현재까지 밝혀진 여덟 종류의 human IAPs는 한 개 이상의 baculovirus IAP repeat (BIR) domain을 가진다. Human IAPs 중 일부는 caspases의 ubiquitination 및 degradation을 조절하는 RING finger do- main을 가지며, 또 다른 일부는 protein-protein interaction 기능을 하는 caspase-recruitment domain (CARD)을 가지는 것으로 알려져 있다.25) IAPs의 중요한 기능은 BIR domain 의존적인 상호작용 및 caspase-3, -7 및 -9의 억제를 통한 apoptosis의 억제이다. 가장 강력한 IAP로 알려진 X-linked IAP (XIAP)는 활성화된 caspases와 높은 친화력을 가지며 apoptosis를 억제한다. XIAP의 BIR3 domain은 caspase-9과 결합하는 반면에 BIR1 및 BIR2 domain은 활성형 cas- pases-3 및 -7와 결합하여 apoptosis를 억제한다.26,27) XIAP 외에도 cIAP-1 및 cIAP-2도 caspases와 결합하여 apoptosis 를 억제하는 것으로 알려져 있다.28,29) 본 실험에서는 ENIL 처리에 따른 IAP family의 발현 변화를 조사해본 결 과, Fig. 4에 나타난 바와 같이 전사 및 번역 수준 모두에 서 큰 변화가 관찰되지 않았으므로 ENIL가 apoptosis에는 아무런 영향을 미치지 못하는 것으로 생각된다.
Caspase protease라는 효소 역시 apoptosis 유발에 중요한 조절인자로서 작용하는 것으로 알려져 있으며 이 단백 질들은 세포에서 핵과 mitochondria의 외막에 불활성 상 태인 proenzyme 형태로 존재한다.30) 많은 caspases 중 ini- tiator caspase인 caspase-8은 활성화된 death receptor에 의해 형성된 DISC에 의해 활성화 되어 effector caspase인 cas- pase-3을 활성화시킨다. 이렇게 활성화된 caspase-3은 PARP 등과 같은 많은 표적 단백질들을 분해함으로서 apoptosis를 유발한다.31) 또 다른 caspase인 caspase-9의 활 성화는 mitochondria에서 유리된 cytochrome c에 의해서 형 성된 apoptosome에 의하여 유발되며 이렇게 활성화된 caspase-9는 caspase-3을 활성화시켜 apoptosis를 일으키는 것으로 알려져 있다.32) 따라서 본 실험에서는 여러 종류 의 caspases 중 apoptosis 유발에 직접적으로 관여하는 것 으로 알려진 caspase-3, -8 및 -9의 발현 및 기질단백질에 미치는 ENIL의 영향에 대해서 조사하였다. Fig. 5A의 결 과에서와 같이 death receptor에 의하여 활성화 되는 것으 로 알려진 caspase-8의 경우는 아무런 변화가 관찰되지 않았지만, mitochondria에 의한 apoptosis 유발에 있어서 중 요한 역할을 하는 caspase-3 및 -9의 경우는 ENIL의 농도 가 증가할수록 비활성형 단백질의 발현이 감소하였으 며, 기질단백질인 PARP의 단편화 현상이 유발되는 것으
로 나타났다. 다음으로 활성형 단백질의 증가를 확인하 기 위해서 in vitro assay를 이용하여 caspases의 활성 정도 를 직접적으로 측정해본 결과, Fig. 5B에 나타난 바와 같 이 Caki-1 세포에서 ENIL 처리에 의해 caspase-3 및 -9의 활성 정도가 증가되었음을 재확인하였다. 이상의 결과 를 살펴보면 ENIL 처리에 의한 apoptosis 유발은 Bcl-2 family의 발현 변화에 의하여 유발되는 mitochondria mem- brane의 permeability의 변화에 따른 caspases의 활성화와 직접 관련이 있음을 알 수 있었다.
한편 호흡과정에서 몸속으로 들어간 산소가 산화과정 에 이용되면서 여러 대사과정에서 생성되는 ROS는 주위 의 화합물과 아주 쉽게 반응하며 전자를 잃거나 얻으려 하기 때문에 높은 반응성을 띠고 있다.33) 병원체나 이물 질을 제거하기 위한 생체방어과정에서 ROS가 많이 발생 하는데, 이들의 강한 살균작용으로 병원체로부터 인체 를 보호하기도 하며, 다양한 암세포의 세포주기 조절작 용에 관여하고 mitochondrial 의존적인 세포사멸을 유도 하여 Bcl-2 member 단백질의 발현을 조절하기도 하는 것 으로 알려져 있다.34,35) 또한 ROS는 다양한 인자에 의하 여 유발될 수 있는데 대표적인 것이 JNK와 같은 스트레 스성 인자들의 조절작용에 의하여 생기는 것으로 알려 져 있다.36,37) 따라서 ENIL에 의한 pJNK 단백질의 발현 변화를 살펴 본 결과, Fig. 6에 나타난 바와 같이 ENIL 처리에 의하여 pJNK의 발현이 현저하게 증가하였지만 JNK 활성의 억제자인 SP600125를 선처리하였을 경우 apoptosis의 억제현상이 나타나지 않았다. 하지만 ENIL 처리에 의하여 ROS가 현저하게 증가하였으며, ROS scav- enger인 NAC에 의하여 apoptosis의 억제가 유발되는 것으 로 보아 ROS가 apoptosis 조절에 아주 밀접한 연관성이 있는 것을 확인할 수가 있었다(Fig. 7). 즉 Caki-1 세포에 ENIL을 처리했을 경우 ROS의 생성됨으로 인하여 Bcl-2 family의 발현변화에 따른 mitochondrial membrane의 per- meability 변화와 그에 따른 caspases의 활성화와 연관된 기질 단백질들의 분해에 의하여 apoptosis가 유발된다는 것을 알 수 있었다.
이상의 결과에서 ENIL을 인체 신세포암세포주인 Caki-1 세포에 처리하였을 경우 처리 농도 및 시간 의존 적으로 생존율 감소 현상이 나타났고, 이러한 생존율의 감소는 apoptotic body의 형성 및 apoptotic cell의 증가와 밀접한 관련이 있는 것으로 조사되었다. 또한 이러한 과 정들이 ROS의 생성과 함께 Bax의 증가, Bcl-2와 Bcl-XL의 발현 감소에 의한 MMP의 소실에 따른 caspase-9의 활성 화와 같은 intrinsic pathway를 거쳐 caspase-3의 활성화 및 기질 단백질들의 분해가 유발되는 것으로 조사되었다.
이러한 연구 결과는 ENIL의 생화학적 항암기전 해석을 이해하고 향후 지속적인 연구를 위한 귀중한 자료로 사 용 될 것으로 생각된다.
감사의 글
본 논문은 2011년도 동의대학교 교내연구비 지원으로 이루어졌음.
참 고 문 헌
1) Motzer RJ, Bander NH, Nanus DM. Renal-cell carcinoma. N Engl J Med 335, 865-875, 1996.
2) Escudier B, Eisen T, Stadler WM. Sorafenib in advanced clear-cell renal-cell carcinoma. N Engl J Med 356, 125-134, 2007.
3) Ratain MJ, Eisen T, Stadler WM. Phase II placebo-controlled randomized discontinuation trial of sorafenib in patients with metastatic renal cell carcinoma. J Clin Oncol 24, 2505-2512, 2006.
4) Ishikawa A, Yamashita H, Hiemori M, Inagaki E, Kimoto M, Okamoto M, Tsuji H, Memon AN, Mohammadio A, Natori Y. Characterization of inhibitors of postprandial hyperglycemia from the leaves of Nerium indicum. J Nutr Sci Vitaminol 53, 166-173, 2007.
5) Ding K, Fang JN, Dong T, Tsim KW, Wu H. Characteri- zation of a rhamnogalacturonan and a xyloglucan from Neri- um indicum and their activities on PC12 pheochromocytoma cells. J Nat Prod 66, 7-10, 2003.
6) Mae SH, Sofia M, Bolhuis RL, Nooter K, Oostrum RG, Subagus W, Ibnu GG. Selectivity of compounds isolated from the leaves of Nerium indicum Mill. on various human cancer cell lines. Med J Malaysia 63, S24-25, 2008.
7) Dong Q, Fang JN. Structural elucidation of a new arabino- galactan from the leaves of Nerium indicum. Carbohydr Res 332, 109-114, 2001.
8) Yu MS, Wong AY, So KF, Fang JN, Yuen WH, Chang RC.
New polysaccharide from Nerium indicum protects neurons via stress kinase signaling pathway. Brain Res 1153, 221-230, 2007.
9) Gayathri V, Ananthi S, Chandronitha C, Ramakrishnan G, Sundaram RL, Vasanthi HR. Cardioprotective effect of Nerium oleander flower against isoproterenol-induced myocardial oxidative stress in experimental rats. J Cardiovasc Pharmacol Ther 16, 96-104, 2011.
10) Harrison JS, Mondor EB. Evidence for an invasive aphid
"Superclone": Extremely low genetic diversity in Oleander aphid (Aphis nerii) populations in the southern United States.
PLoS One 6, e17524, 2011.
11) Rashan LJ, Franke K, Khine MM, Kelter G, Fiebig HH,
Neumann J, Wessjohann LA. Characterization of the anti- cancer properties of monoglycosidic cardenolides isolated from Nerium oleander and Streptocaulon tomentosum. J Ethnopharmacol 134, 781-788, 2011
12) Leist M, Jäättelä M. Four deaths and a funeral: from caspases to alternative mechanisms. Nat Rev Mol Cell Biol 2, 589-598, 2001.
13) Vilches Troya J. Understanding cell death: a challenge for biomedicine. An R Acad Nac Med 122, 631-656, 2005.
14) Huerta S, Goulet EJ, Huerta-Yepez S, Livingston EH.
Screening and detection of apoptosis. J Surg Res 139, 143-156, 2007.
15) Huerta S, Goulet EJ, Livingston EH. Colon cancer and apoptosis. Am J Surg 191, 517-526, 2006.
16) Nagata S. Apoptotic DNA fragmentation. Exp Cell Res 256, 12-18, 2000.
17) Lin S, Fujii M, Hou DX. Rhein induces apoptosis in HL-60 cells via reactive oxygen species-independent mitochondrial death pathway. Arch Biochem Biophys 418, 99-107, 2003.
18) Jacobson MD. Reactive oxygen species and programmed cell death. Trends Biochem Sci 21, 83-86, 1996.
19) Du J, Chen GG, Vlantis AC, Chan PK, Tsang RK, van Hasselt CA. Resistance to apoptosis of HPV 16-infected laryngeal cancer cells is associated with decreased Bak and increased Bcl-2 expression. Cancer Lett 205, 81-88 2004.
20) Reed JC. Bcl-2 family proteins. Oncogene 17, 3225-3236, 1998.
21) Kroemer G. The proto-oncogene Bcl-2 and its role in regulat- ing apoptosis. Nat Med 3, 614-620, 1997.
22) Rosse T, Olivier R, Monney L, Rager M, Conus S, Fellay I, Jansen B, Borner C. Bcl-2 prolongs cell survival after Bax- induced release of cytochrome c. Nature 391, 496-499, 1998.
23) Salvesen GS, Duckett CS. IAP proteins: blocking the road to death's door. Nat Rev Mol Cell Biol 3, 401-410, 2002.
24) Deveraux QL, Reed JC. IAP family proteins-suppressors of apoptosis. Genes Dev 13, 239-252, 1999.
25) Checinska A, Hoogeland BS, Rodriguez JA, Giaccone G, Kruyt FA. Role of XIAP in inhibiting cisplatin-induced caspase activation in non-small cell lung cancer cells: a small molecule Smac mimic sensitizes for chemotherapy-induced apoptosis by enhancing caspase-3 activation. Exp Cell Res 313, 1215-1224, 2007.
26) Chai J, Shiozaki E, Srinivasula SM, Wu Q, Datta P, Alnemri ES, Shi Y. Structural basis of caspase-7 inhibition by XIAP.
Cell 104, 769-780, 2001.
27) Huang Y, Park YC, Rich RL, Segal D, Myszka DG, Wu H.
Structural basis of caspase inhibition by XIAP: differential roles of the linker versus the BIR domain. Cell 104, 781-790, 2001.
28) Deveraux QL, Roy N, Stennicke HR, Van Arsdale T, Zhou Q, Srinivasula SM, Alnemri ES, Salvesen GS, Reed JC. IAPs block apoptotic events induced by caspase-8 and cytochrome c by direct inhibition of distinct caspases. EMBO J 17, 2215- 2223, 1998.
29) Roy N, Deveraux QL, Takahashi R, Salvesen GS, Reed JC.
The c-IAP-1 and c-IAP-2 proteins are direct inhibitors of specific caspases. EMBO J 16, 6914-6925, 1997.
30) Nagata S. Apoptosis by death factor. Cell 88, 355-365, 1997.
31) Chai F, Truong-Tran AQ, Ho LH, Zalewski PD. Regulation of caspase activation and apoptosis by cellular zinc fluxes and zinc deprivation: a review. Immunol Cell Biol 77, 272-278, 1999.
32) Green DR, Reed JC. Mitochondra and apoptosis. Science 281, 1309-1312, 1998.
33) Sauer H, Wartenberg M, Hescheler J. Reactive oxygen species as intracellular messengers during cell growth and differen- tiation. Cell Physiol Biochem 11, 173-186, 2001.
34) Ling YH, Liebes L, Zou Y, Perez-Soler R. Reactive oxygen species generation and mitochondrial dysfunction in the apoptotic response to Bortezomib, a novel proteasome inhibi-
tor, in human H460 non-small cell lung cancer cells. J Biol Chem 278, 33714-33723, 2003.
35) Zhang Y, Chen F. Reactive oxygen species (ROS), troublema- kers between nuclear factor-kappaB (NF-kappaB) and c-Jun NH(2)-terminal kinase (JNK). Cancer Res 64, 1902-1905, 2004.
36) Moungjaroen J, Nimmannit U, Callery PS, Wang L, Azad N, Lipipun V, Chanvorachote P, Rojanasakul Y. Reactive oxygen species mediate caspase activation and apoptosis induced by lipoic acid in human lung epithelial cancer cells through Bcl-2 down-regulation. J Pharmacol Exp Ther 319, 1062-1069, 2006.
37) Kim BC, Kim HG, Lee SA, Lim S, Park EH, Kim SJ, Lim CJ. Genipin-induced apoptosis in hepatoma cells is mediated by reactive oxygen species/c-Jun NH2-terminal kinase-depen- dent activation of mitochondrial pathway. Biochem Pharmacol 70, 1398-1407, 2005.