미생물과 식물에 의해 생성된 탄산칼슘의 광물학적 특성 관찰
남인현1)* · 전철민1)· 김재곤1)
Mineralogical Observation of Calcium Carbonate Precipitation Properties by Bacterial and Plant Ureases
In-Hyun Nam*, Chul-Min Chon and Jae-Gon Kim
(Received 4 October 2016; Final version Received 26 October 2016; Accepted 26 October 2016)
Abstract : Calcium carbonate(CaCO3) can be precipitated by urea hydrolysis by bacterial and plant ureases.
Sporosarcina sp. KM-01, KM-07, and KM-12 strains were shown to be able to precipitate CaCO3 using urea amended with calcium chloride. Also, an extract of the plant, Canavalia ensiformis, was shown to produce CaCO3 as an effective biomaterial. The mineralogical formation of CaCO3 using bacteria was compared to that of plant urease in this study. The results revealed that urease in the Canavalia ensiformis extracts precipitated mainly calcite, however, the ureases in the bacterial strains produced vaterite in liquid-state cultures. Considering acidic habitats of the isolated bacterial strains, these results could be attributed to their evolutionary capabilities or the addition of specific amino acid. This makes the general application of biomineralization process based on stimulation of urea hydrolysis and potentially useful for a variety of soil bioengineering and environmental bioremediation.
Key words : Calcium carbonate, Urease, Bacteria and plant, Calcite, Vaterite
요 약 : 본 연구에서는 미생물과 식물이 가지는 요소분해효소 작용에 의해 침전되는 탄산칼슘(CaCO3)의 광물학적
특성을 관찰하고 비교하였다. 중금속으로 오염된 산성 토양에서 분리한 Sporosarcina sp. KM-01, KM-07, KM-12 등 의 박테리아 균주 및 Canavalia ensiformis 식물 추출액에 의해 침전되는 CaCO3를 X-선회절분석(XRD) 및 전자현미
경(SEM) 분석을 통해 확인하였다. 박테리아 균주로부터 침전되는 CaCO3은 바테라이트가 주 광물상이었으나, 식물
에 의해 생성되는 CaCO3의 주 광물상은 방해석으로 확인되었고, 일부 바테라이트가 검출되었다. 박테리아의 경우 진
화적 요인에 의해 낮은 pH에 잘 적응된 균주의 요소분해효소 특이적 활성의 차이로 바테라이트가 생성되고, 아직 명 확하게 규명되지 않은 바테라이트 결정 구조와 특정 아미노산의 영향으로 해당 구조가 유지되었을 가능성을 예상할 수 있다. 이러한 결과는 생광물화작용 및 토양환경복원 등의 분야에 적용 시 유용한 자료로 활용될 것을 기대한다.
주요어 : 탄산칼슘, 요소분해효소, 박테리아와 식물, 칼사이트, 바테라이트
1) 한국지질자원연구원 지질환경연구본부
*Corresponding Author(남인현) E-mail; [email protected]
Address; Geologic Environment Research Division, Korea Institute of Geoscience and Mineral Resources (KIGAM), Daejeon, Korea
서 론
토양에 존재하는 일부 미생물은 요소(Urea)와 칼슘원 등 이 주어지는 적합한 환경일 경우, 요소분해효소(Urease)를 유도한다. 요소분해효소의 작용은 아래 화학식과 같이 요 소가 가수분해되어 NH4+ 이온과 CO32- 이온으로 해리되어 주변의 pH를 높여주고(식 (1)), 세포(Cell)에 붙어있는 Ca2+
이온이 생성된 CO32-
이온과의 결합으로 CaCO3 침전을 유 도하며(식 (2)), 이러한 생물학적 과정을 기존 연구들에서 MIC(C)P(Microbially Induced Calcium Carbonate Preci- pitation)라고 명명하였다(DeJong et al., 2006; Kawasaki et al., 2010; Mitchell and Santamarina, 2005; Mobley and Hausinger, 1989; Terajima et al., 2009; Van Paassen et al., 2010; Whiffin et al., 2007).
CO(NH2)2 + 2H2O → 2NH4+ + CO32- (1) CO32-
+ Ca2+ → CaCO3 (2) 연구논문
요소분해효소는 박테리아를 포함한 일부 미생물과 식물 등 여러 생물 종에 분포하며, 식물 중에는 콩과류 식물 작두 콩(Jack bean)에 많이 함유된 것으로 알려져 있고 농경지의 질소순환 등 자연계에 존재하는 여러 형태의 질소화합물 변환에 중요한 역할을 수행하여(Min and Lee, 2007) 일부 세균의 병원성과 산성 환경에서의 생존에 필수적인 요소로 알려져 있다(Mobley and Hausinger, 1989). 박테리아의 요 소분해효소 유전자 염기서열은 1989년 요로감염의 원인세 균인 Proteus mirabilis에서 처음으로 밝혀진 이래 Klebsiella aerogenes, Helicobacter pylori, Yersinia enterocolitica, Streptococcus salivarius, Vibrio parahaemolyticus 등 다 수의 균주로부터 확인되었다(Chen et al., 1996; De Koning- Ward et al., 1994; Jones and Mobley, 1989; Labigne et al., 1991; Mulrooney and Hausinger, 1990; Park et al., 2000).
지금까지 박테리아로부터 보고된 요소분해효소 유전자는 구조적 배열이 유사한 7 개 이상의 형태로 구성되어 있고, 크게 구조단백질(UreA, UreB, UreC)과 보조단백질(UreD, UreE, UreF, UreG)로 나뉜다. 구조단백질은 효소가 활성 을 갖는 데에 중심역할을 하고 보조단백질은 구조단백질에 Ni2+을 운반하여 이를 조합시키는 역할을 담당한다(Kim et al., 2009). 특히, UreC 단백질은 요소분해효소 구조단백질 의 Subunit으로 가장 큰 Gene coding을 가지며, PCR priming site가 안정하고 염기서열이 가장 보존적인 부위이 기 때문에 요소분해효소의 Target 유전자 부위로 이용된다 (Grasham and Sheridan, 2007; Koper et al., 2004). 일부 박 테리아에서는 특이적으로 요소분해효소 유전자의 전사를 조절하는 전사조절 단백질 UreR 및 세포내로 Ni2+를 운반 하는 NixA와 UreH 등의 보조단백질도 발견된 바 있다 (Kim et al., 2009). 요소분해효소 활성을 저해시킬 수 있는 물질로 유기화합물의 경우에는 Catechol이 평균 74%, Phenyl mercuric acetate가 67%, Hydroquinone이 64% 정 도로 요소분해효소 활성을 저해시킬 수 있고, 무기화합물 로는 Silver sulphate가 48%, Mercuric chloride가 37%, Gold chloride가 18% 정도 요소분해효소 활성을 저해할 수 있으며, 여러 중금속으로 활성저해 실험을 수행한 결과 대 부분의 중금속이 요소분해효소 활성을 저해하는 것으로 확 인된 바 있다(Bremner and Douglas, 1971; Tabatabai, 1973).
탄산칼슘(Calcium carbonate, CaCO3)은 여러 광물자원 중 풍부하게 존재하며, 이를 생성시키는 연구는 화학적, 생 물학적으로 다양하게 진행되어 왔다. 기존 연구에서 다양 하게 알려진 바와 같이 일반적으로 CaCO3은 이를 만드는 방법에 따라 원석의 파쇄 및 분쇄로부터 기인하는 중질 탄 산칼슘(Ground calcium carbonate)과 화학적 혹은 생물학 적인 방법으로 침전 반응에 의해 얻어지는 침강성 탄산칼 슘(Precipitated calcium carbonate)으로 크게 나눌 수 있다
(Park et al., 2006; Song et al., 2011). 또한, CaCO3은 반응 조건 및 상(phase)에 따라 무정형 탄산칼슘 ACC(Amorphous Calcium Carbonate), 결정형 탄산칼슘 방해석(Calcite), 아 라고나이트(Aragonite), 바테라이트(Vaterite) 등 세 종류 의 형태로 변할 수 있다. 이들의 결정 구조는 각기 다른 형태 를 지니는데, 방해석 결정은 직사각형 모양을 가지고 아라 고나이트 결정은 바늘 모양, 바테라이트는 원형 모양으로 각각 독특한 형태를 지니고 있어서 이를 형태학적으로 관 찰할 때 구별하기 용이하다. 결정을 형성하는 조건으로는 가장 열역학적으로 안정한 방해석은 높은 pH와 낮은 온도 에서 결정이 잘 형성되며, 아라고나이트와 바테라이트는 방해석에 비해 불안정하며 낮은 pH와 높은 온도가 결정 형 성이 잘 되는 조건이다. 이들의 결정형태 변화는 상전이에 의해서 이루어지고 대부분의 반응이 지속되면 자체적으로 재결정이 일어나 안정한 방해석으로 변화한다(Gu et al., 2001).
이러한 CaCO3이 생성되는 조건 중 하나인 요소분해효소 의 생화학적인 반응은 온도, 습도, pH, 이온의 농도 등 주어 진 조건이 적합할 경우 CaCO3광물질을 침전시킨다. 특정 재료 표면이나 분리된 공간 사이에 미생물로 인하여 광물 질이 쌓이거나 형성되는 것은 Biodeposition이라고 하고, 입자 사이에 미생물 작용으로 생성된 광물질이 고결을 일 으키는 것을 Biocementation이라고 하는데, 이는 미생물이 요소분해효소 작용으로 요소를 분해하여 암모니아와 탄산 이온으로 바뀌면서 주변이 있는 칼슘이온과 결합하여 CaCO3광물질이 생성되는 과정을 나타낸 바 있다(De Muynck et al., 2010). 기존 연구에서 요소분해효소에 의해 침전되 는 CaCO3이 시멘트나 토양의 고결작용을 유도하는 것으 로 알려져 있으며 토양 박테리아의 하나인 Sporosarcina pasteurii의 Urease가 대표적이다(DeJong et al., 2006;
Mobley and Hausinger, 1989; Park et al., 2011; Van Paassen et al., 2010; Whiffin et al., 2007). 또한, 박테리아 등의 미생물 외 콩과류 식물 내 요소분해효소를 이용하여 CaCO3을 생성시키고 이 반응을 활용하여 토양 내 중금속 을 안정화하고, 이를 통해 미생물 군집이 변화하여 식생을 원활하게 유도한 연구가 최근 진행된 바 있다(Nam et al., 2016; Park et al., 2014; Roh et al., 2015). 이렇게 미생물에 의해 분비되거나 식물 내에 존재하는 요소분해효소를 활용 하여 CaCO3의 침전을 유도할 경우, 약화된 지반을 친환경 적으로 메우거나 고결시켜 지반의 강도 증가 및 토양 입자 간 공극을 효과적으로 메울 수 있다(Lee and Park, 2012;
Park et al., 2011). 그러나, 생화학적인 효소에 의해 생성되 는 CaCO3이 보다 제어된 환경에서 침전되는 것이 아니어 서 생성되는 CaCO3에 대한 상(Phase)에 대한 정확한 규명 은 이루어진 바가 없다.
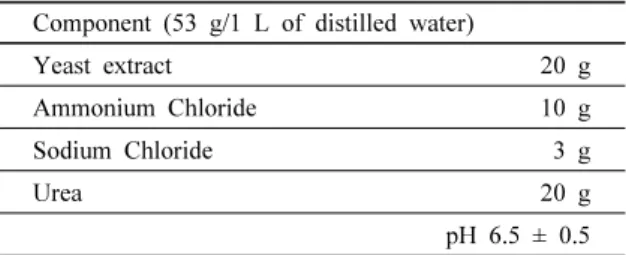
Table 1. Urea medium composition used for urease-producing bacteria cultivation
Component (53 g/1 L of distilled water)
Yeast extract 20 g
Ammonium Chloride 10 g
Sodium Chloride 3 g
Urea 20 g
pH 6.5 ± 0.5 따라서, 본 연구에서는 분리된 요소분해 활성을 가지는 미생물에 의해 침전되는 CaCO3을 확인하여 그 성상을 파 악하고, 식물에서 유래한 요소분해효소에 의해 침전되는 CaCO3과의 비교를 통해 광물학적인 차이를 규명함으로써 생물학적으로 침전될 수 있는 CaCO3의 다양성을 고찰하 고자하였다.
연구방법
미생물에 의한 탄산칼슘 생성 실험
본 연구에서 활용한 요소분해효소 생산 박테리아 균주 3 종은 중금속으로 오염된 산성 토양으로부터 멸균된 50 mL 원심분리 튜브에 채취하여 Urea 배지(Table 1)를 사용하여 분리하였고(Park et al., 2014), 고체배지로는 1.5% Agar를 첨가한 Urea 배지를 사용하였다(De Muynck et al., 2010).
분리 균주들은 50% Glycerol이 첨가된 Cryogenic vial에 보관하여 –70°C에서 동결보존 하면서 각 실험에 사용하였 으며, 실험 전 Table 1에 나타낸 조성의 배지에서 2회 전 배 양(pre-culture)을 실시한 후 사용하였다. CaCO3침전 실험 은 분리된 박테리아 균주들의 전 배양액 1 mL를 Urea 액체 배지 400 mL에 접종하여 Shaking incubator에서 진탕 배 양하였다(28°C, 160 rpm). 시간 별로 침천물 생성량을 비 교하기 위해 총 96 시간 동안 12 시간마다 배양액 25 mL를 회수하여 10 분간 원심분리 (4°C, 8,000 rpm)하여 상등액 을 제거하고 남은 균체와 침전물을 50°C Dry oven에서 24 시간 건조한 후 무게를 측정하였다.
식물에 의한 탄산칼슘 생성 실험
식물에 의한 CaCO3생성 실험은 요소분해효소가 많이 함유된 것으로 알려져 있는 콩과류 식물 중 Canavalia ensiformis 50 g을 분쇄기(Homogenizer)로 20 mM Phosphate buffer(pH 7.0)와 함께 분쇄시킨 후, 원심분리(12,000 rpm, 20 min)하여 그 상등액을 식물추출액 시료로 사용하였다.
20 mM Phosphate buffer 는 기존 연구와 동일하게 제조하 여 사용하였고(Nam et al., 2006; 2008), 칼슘원인 CaCl2과
기질 요소 용액의 농도는 각각 CaCl2 4g/100 mL, 요소 2 g/100 mL의 농도로 제조하여 순차적으로 공급하였다. 실 험 방법은 CaCl2, 요소 용액을 100 mL씩 주입한 후 식물추 출액을 50 mL 주입하였다. 용액 주입 후부터 96 시간까지 상온에서 반응시키면서 시간에 따라 CaCO3의 확인 및 정 량 실험을 진행하였다.
침전된 탄산칼슘 분석 방법
미생물과 식물에 의해 각각 생성된 CaCO3의 X-선회절 분석 및 정량은 리트벨트법에 기반한 DIFFRAC PLUS TOPAS 소프트웨어를 이용하여 실험결과로 얻은 CaCO3
및 다양한 물질의 구성광물 조성 및 비정질 함량을 정량화 하고 이에 따라 발생하는 정량 산출값 변화를 평가하였다.
분석 대상 시료에 대하여 각각 분말시료 3 g과 내부표준시 료(Calcium fluoride, CaF2, ultra dry, 99.99%, metal basis, Alfa Aesar) 0.3333 g을 아게이트 막자사발에서 잘 혼합하여 분말화하였다. X-선회절분석 조건은 Cu target과 LynxEye position sensitive detector가 부착된 D8 Advance diffractometer (Bruker-AXS, Germany)를 이용하여 수행되었다. 회절패 턴은 5° to 100° 2 theta 구간, 0.01° step size, step 당 1 sec의 조건으로 취득되었으며, 0.3° divergence slit이 사용되었 다. 시료의 정성분석 및 기기인자의 표준화(Fundamental parameter)를 위해 동일한 조건으로 원시료와 표준시료 (Calcium Fluoride, CaF2, ultra dry, 99.99%, metals basis, Alfa Aesar)에 대하여 각각 X-선 회절패턴을 취득하였다.
분석결과는 리트벨트법에 기반을 둔 구성광물의 정량분석 을 위하여 TOPAS 4.2(Bruker-AXS, Germany) 소프트웨어 를 사용하였다. 또한, 미생물 및 식물에 의해 생성된 CaCO3
침전물의 형태학적인 확인을 위해 전자현미경 SEM(Scanning Electron Microscope, JSM-7000F, Jeol, Japan) 및 EDS(Energy Dispersive Spectrometer) 분석을 실시하였다. 각 시료를 완전히 건조시킨 후 0.5 cm cube pieces로 제작하고 Platinum sputter 코팅을 통해 관찰하였다. 가속전압 범위는 10 kV로 수행하였다.
결과 및 토의
미생물에 의한 탄산칼슘 생성 실험 결과
분리 박테리아 균주 3종(Sporosarcina sp. KM-01, KM- 07, KM-12)에 대해 CaCO3침전능을 확인하기 위해 Urea 액체배지에 염화칼슘(Calcium Chloride, CaCl2)을 첨가하 여 배양한 후, 각 균주 내에서 생성되는 요소분해효소에 의 해 생성된 CaCO3 침전물의 무게를 측정하였다. 그 결과, 3 종의 균주 모두에서 시간이 지남에 따라 무게가 증가하는 경향성을 나타내었다(Fig. 1).
Fig. 1. Time course of the precipitated solid materialweight by culture of Sporosarcina sp. KM-01, KM-07, and KM-12.
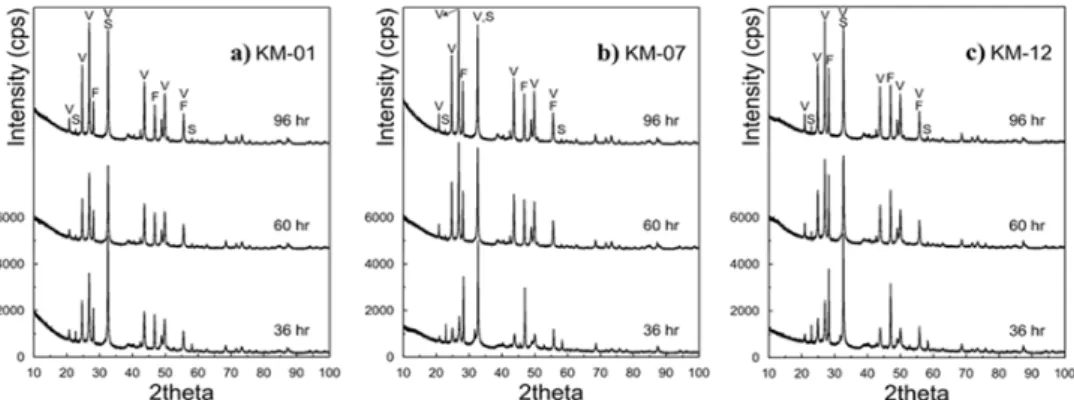
Fig. 2. X-Ray diffraction (XRD) patterns of the precipitated CaCO3 by Sporosarcina sp. KM-01(a), KM-07(b), and KM-12(c). V, Vaterite; F, Fluorite; S, Salammoniac).
Fig. 1에 나타난 바와 같이, KM-01 균주는 배양 24 시간 이 지나면서 침전물 생성량이 급격하게 증가하였고, KM-07 균주는 36 시간이 지나는 시점에서 침전물의 양이 급격히 증가하는 경향성을 나타내었다. 반면에, KM-12 균 주의 경우에는 KM-01 및 KM-07 균주들이 나타낸 경향성 과는 다소 차이가 있게 침전물이 급격하게 증가하는 시점 없이 꾸준하게 침전물 양이 증가하는 경향을 나타내었다.
본 균주들을 분리하여 특성을 연구한 기존 연구에서 KM-01 균주의 생장 및 활성이 초기에 가장 높게 나타났는 데(Park et al., 2014), 이를 토대로 침전물 양의 증가 시점이 가장 빠른 원인은 균주의 생장이 가장 초기에 활발해지기 때문인 것으로 판단된다. 또한, KM-01 및 KM-07 균주의 경우, 96 시간 이후에는 침전물의 양이 감소하는 경향을 보 이는데, 이는 초기 생장이 매우 활발하여 배지에 첨가된 칼 슘원 및 균주 생장에 필요한 영양분(Nutrients)의 고갈로 인해 요소분해효소 활성이 감소하였기 때문으로 예상할 수 있다.
박테리아 균주들에 의해 생성된 침전물의 분석을 위해 배양 시간에 따라 얻어진 침전물을 각 36 시간, 60 시간, 96 시간에 시료 채취 후, X-선회절분석(XRD)을 실시하여 침 전물의 광물조성을 분석하였다(Fig. 2). 각 균주에 의해 생 성된 침전물들의 주 구성광물은 바테라이트(CaCO3)였고, 정량분석을 위해 내부표준물질로 첨가한 CaF2의 영향으로 Fluorite 가 관찰되었으며, Salammoniac이 일부 관찰되었 다. 또한, 침전물의 주 구성광물인 바테라이트(CaCO3) 정 량 분석을 실시한 결과를 Fig. 3에 나타내었는데, 전체의 침 전물 측정 결과(Fig. 1)와 동일한 경향으로 KM-01 균주에 의해 생성된 바테라이트 양이 가장 많았으며, 반응 36 시간 에 회수한 침전물에서 KM-07, KM-12에 의해 생성된 바테 라이트 양이 오차범위 내에서 비슷한 수준인 것을 관찰할 수 있었다. 또한, 배양 60 시간 후에는 세 종 모든 박테리아 균주에 의해 생성된 바테라이트 정량 값이 유사하게 나타 남을 확인하였다. 상기한 바와 같이 이 결과는 Fig. 1에 나 타낸 침전물 무게 측정 결과와 경향성이 유사하여 침전물 의 주 구성광물이 바테라이트라는 것을 의미하고, 생장곡 선 측정 결과와도 동일한 경향을 나타낸 것으로 균주에서 발생하는 요소분해효소 활성이 바테라이트 침전에 직접적 으로 영향을 미친 것이라고 할 수 있다.
박테리아에 의해 침전된 CaCO3형태를 시각적으로 확인 하기 위해 각 침전물을 전자현미경(SEM) 및 EDS 원소 분 석 등을 실시하였다. XRD 분석 결과(Fig. 3)에서 나타낸 바 와 같이 분리한 3 종의 균주 모두 바테라이트 침전물을 생 성시키는 것을 형태학적으로 확인하였고, 대체로 CaCO3
의 원소비율과 비슷한 Ca:C:O 의 비율인 1:1:3을 나타냈다 (Fig. 4). 모든 시료에서 CaCO3침전물이 바테라이트의 전 형적인 결정 형태인 둥근 모양의 형태를 나타냄을 확인 할 수 있었다. 기존에 수행된 MICP 관련 연구들에서는
Fig. 3. X-Ray diffraction (XRD) quantification analytical results of precipitated CaCO3 by 3 bacterial cultures.
Fig. 4. Scanning Electron Microscope (SEM) and Energy Dispersive Spectrometer (EDS) results of the precipitated CaCO3 by Sporosarcina sp. KM-01(a), KM-07(b), and KM-12(c).
Sporosarcina pasteurii 균주를 중심으로 방해석을 박테리 아 배양에 의해 침전시키는 연구를 수행하였다(DeJong et al., 2006; Kawasaki et al., 2010; Mitchell and Santamarina, 2005; Mobley and Hausinger, 1989; Terajima et al., 2009;
Van Paassen et al., 2010; Whiffin et al., 2007). 거의 모든 연구에서 바테라이트 생성에 대한 고찰은 이루어지지 않았 고, 일부 연구에서 박테리아 또는 식물 내 요소분해효소에 의해 생성된 바테라이트가 시간이 지남에 따라 보다 열역 학적으로 안정한 방해석으로 상 전이가 이루어진다는 연구 결과를 보고한 바 있다(Sondi and Salopek-Sondi, 2005).
그러나, 본 연구에서 관찰한 결과에서는 배양 96 시간이 지 난 후에도 바테라이트 생성이 두드러졌으며(Fig. 2), 이를 1 개월간 방치한 후에도 방해석으로의 전이가 이루어지지 않 았다. 기존 연구들에서 분리 및 활용한 박테리아 종과 본 연 구에서 활용한 Sporosarcina sp. KM-01, KM-07, KM-12 등의 균주 간 유연관계가 높기는 하나, 연구방법에 기술한 바와 같이 본 연구에서 활용한 박테리아 균주들은 중금속 으로 오염된 산성 토양에서 분리하였다. 일반적으로 바테 라이트는 방해석과 달리 낮은 pH와 높은 온도가 결정 형성 이 잘 되는 조건이며, 바테라이트 결정의 유지는 특정 아미 노산이 영향을 미칠 수 있다(Gu et al., 2001). 또한, 박테리 아 균주가 생존하는 초기의 산성 조건에서 수소이온의 세 포질 내 유입은 요소분해효소 활성도를 증가시켜 세포질 내 Urea를 Ammonia와 Carbonic acid로 가수분해하여 생 성된 암모니아는 수소이온을 받아들여 암모늄이 되고, 세 포질 내의 산성도를 중화시킨다. 또한, 생성된 암모늄은 세 포질내로 유입된 수소이온과 함께 공수송(Symport)을 통 하여 세포 밖으로 유출되어 세포내의 pH 감소를 막는다 (Young et al., 1996). 따라서, 진화적인 요인에 의해 낮은 pH에 잘 적응된 균주의 요소분해효소 특이적 활성 및 영향 성의 차이로 바테라이트가 생성되고 아직 명확하게 규명되
지 않은 바테라이트 결정 구조와 특정 아미노산의 영향(Gu et al., 2001; Kabalah-Amitai et al., 2013)으로 해당 구조가 유지되었을 가능성을 예상할 수 있겠다.
이와 같이 미생물을 활용하여 이산화탄소와 암모니아를 생성하는 요소 가수분해 과정과 요소 분해를 통한 생산물 로 박테리아 주변의 pH를 변화시키는 과정은 생태학적으 로 요소분해효소의 중요한 특이성이다(Koper et al., 2004). 따 라서, 이러한 특이성을 활용한 MICP 기법을 Bioremediation 과정에 적용한 연구가 진행되었는데(Nam et al., 2016; Roh et al., 2015), 이는 요소분해효소를 생성하는 Ureolytic bacteria 가 다양한 환경에 널리 존재하며, in-situ 환경복원 과정에 서 요소 가수분해 반응의 특이성을 바로 활용할 시에 완전 히 새로운 미생물을 적용할 필요가 없는 장점이 있기 때문 이다. 또한, 요소분해효소를 생성하는 여러 미생물을 통해 손쉽게 환경복원에 적용이 가능한 이유는 일반적인 박테리 아 세포 표면이 음전하를 띠고 있으므로 Ca2+와 Mg2+와 같 은 2가 양이온과 잘 반응하여 세포 표면에 잘 흡착되므로 미생물 주변에서 광물 형성이 일어나기에 유리하기 때문이 다(Okwadha and Li, 2010).
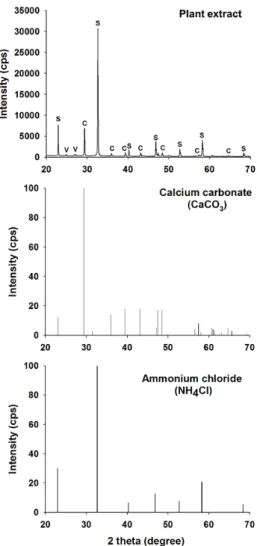
Fig. 5. X-Ray diffraction (XRD) patterns of the precipitated CaCO3 by plant extract. The spectra were compared with authentic spectra as depicted in CaCO3, in specific, calcite(c) and NH4Cl(d). C, Calcite; V, Vaterite; S, Salammoniac.
Fig. 6. Scanning electron microscope (SEM) images for showing CaCO3 precipitation by plant urease. Calcite(a) was mainly observed and the small amounts of vaterite were observed in this study. All data were obtained after 72 hours incubation (bar, 10 μm).
식물에 의한 탄산칼슘 생성 실험 결과
식물에 의한 CaCO3생성 실험은 요소분해효소가 많이 함유되어 있는 것으로 잘 알려져 있는 콩과류 식물 중 한 종 (Canavalia ensiformis)을 분쇄하여 그 상등액을 활용하여 수행하였고, 침전 반응물의 확인을 위해 X-선 회절분석 (XRD) 및 전자현미경(SEM) 분석 등을 수행하였다. XRD 분석 결과, Fig. 5에 나타낸 것과 같이 CaCO3이 생성됨을 확인하였고, 반응 시간에 따라 침전물이 증가하는 경향성 도 리트벨트 법에 기반한 정량 분석을 통해 확인하였다. 이 는 식물추출액을 포함하는 요소분해효소 작용이 일정 시간 동안 지속적인 반응을 유지하는 것을 의미하며, 향후 환경 매체에 적용하는 연구에서 응용성이 확대될 것으로 사료된 다. CaCO3생성 확인은 Fig. 5에 나타낸 바와 같이 표준물 질을 동시에 분석하여 피크를 확인하였고, 칼슘원인 CaCl2 과 기질로 사용한 요소가 반응하여 생성된 NH4Cl도 동일 분석에서 확인하였다(Fig. 5).
XRD 분석을 통해 확인한 바와 같이 식물추출액에 의해 생성된 CaCO3를 시각적으로 확인하기 위해 전자현미경 (SEM)을 이용하여 그 형태를 Fig. 6과 같이 관찰하였다.
Fig. 6에 나타낸 바와 같이 식물추출액을 이용해 반응을 진 행시킨 결과, CaCO3생성을 직접 확인할 수 있었으나, 상기 한 바와 같이 박테리아에 의해 생성된 CaCO3, 즉 바테라이 트와는 다른 형태, 방해석의 전형적인 직사각형 결정 형태 를 나타내고 있다(Fig. 6(a)). 침전물의 일부가 방해석 형태 와 다소 다르게 관찰되어 이를 확대해 본 결과, 바테라이트 결정 형태인 둥근 모양이 검출되었으나, 그 검출 빈도가 매 우 낮았다(Fig. 6(b)). 이러한 CaCO3 침전 특성은 XRD 분 석 결과에서도 확인할 수 있었는데(Fig. 5), 일부 바테라이 트 피크가 검출되기는 하였으나, 대부분의 피크가 침전된 생성물이 방해석임을 지시하고 있다(Fig. 5).
이렇게 박테리아에 의해 침전된 CaCO3과 식물에 의해 생성된 CaCO3의 결정 형태가 다른 것에 대한 이유는 아직 명확하지는 않다. 다만, 기존 연구에서 박테리아로부터 발
생하는 요소분해효소와 식물 내에 있는 요소분해효소의 아 미노산 서열(Amino acids sequence) 차이를 분석해 본 결 과, 동일한 활성을 가진 효소임에도 불구하고 서열의 차이 가 확연하게 나타난 바 있다(Sondi and Salopek-Sondi, 2005). 또한, CaCO3결정화 및 상 전이에 있어서 특정 아미 노산을 첨가하여 CaCO3결정의 성질이나 형태, 특성의 변화 를 연구한 다른 기존 연구에서 CaCO3의 Biomineralization 반응을 위해 다양한 아미노산을 첨가한 결과, CaCO3용액 에 Glutamic acid가 첨가되면 바테라이트 상을 안정하게 만들고(Manoli and Dalas, 2001), Aspartic acid도 바테라 이트를 형성할 수 있는 활성이 있으며, 형성된 결정을 안정 하게 유지하는 능력이 있다고 알려진 바 있다(Xie et al., 2005). 대부분의 CaCO3결정에 첨가된 아미노산은 바테라 이트 형성에 효율적이고, 결정이 장기간 유지될 수 있도록 하여 충분한 양의 아미노산이나 Glycine의 첨가는 CaCO3
결정화 반응에서 바테라이트의 침전물을 발생시킨다고 보 고하였다(Shivkumara et al., 2006). 이를 토대로 본 연구에 서 나타낸 박테리아 및 식물 요소분해효소에 의해 생성되 는 CaCO3의 결정 생성의 차이를 유추하면, 각 효소가 가지 는 활성은 유사하지만 그 활성이 미치는 영향력의 범위 및 조건이 다르므로 결정 상의 차이가 나타날 가능성이 있다 고 생각할 수 있으며, 해당 조건은 반응에 관여하는 효소로 부터의 아미노산의 차이, 박테리아의 경우 해당 균주가 지 니고 있는 기타 유전자로부터 발현되는 기타 단백질의 영 향 등을 포함할 수 있다.
방해석, 바테라이트와 같은 MICP 기법의 생산물은 표면 에 중금속 이온을 매우 강하게 흡착할 수 있어 그들의 광물 구조에 중금속이온을 포함하기도 한다. CaCO3가 침전되 는 동안 Ca2+과 비슷한 이온반지름을 갖는 Sr2+, Pb2+, Cd2+, Cu2+같은 중금속 이온이 CaCO3광물 구조 내에서 Ca2+대 신 치환되거나, 광물의 입자 사이에 중금속 이온이 침투하 여 동시에 침전 될 수 있다. 따라서, 본 연구에서 관찰한 MICP 기법은 환경 연구 분야에 적용할 수 있고, 중금속을 잘 흡착하므로 오염된 토양을 in-situ로 정화하는 방법으로 여겨진다(Achal et al., 2011). 또한, MICP 기법은 지하수 거동을 통해 지하에 존재하는 바륨(Ba), 스트론튬(Sr)과 같 은 방사성원소와 미량원소 오염을 잡아내는 데에도 효율적 인 방법이다. 금속이온과 결합하여 광물질을 형성하는 미 생물 활동 과정은 다양한 응용 범위에서 영향을 일으킬 수 있어서 생지화학측면에서 주요한 부분으로 여겨지고 있고, 특히 CaCO3 침전은 Biomineralization 측면에서 중요한 연 구 분야로 인식되고 있다. 본 연구에서 살펴본 바와 같이 Biomineral은 무기적으로 생성된 광물과 비교하면 독특한 크기, 결정도, 동위원소와 미량원소를 포함하는 등의 특성 을 갖는다. 생물학적으로 CaCO3 침전을 시킬 수 있는 대표
적인 2 가지의 방법을 박테리아와 식물이 발생시키는 요소 분해효소 작용에 의한 기작으로 알아본 결과, 방해석과 바 테라이트로 대표되는 서로 다른 결정 형태를 나타내었다.
이 원인으로 본 연구에서 몇 가지의 가능성을 제기하기도 하였지만, 보다 명확한 원인을 규명하기 위해서 각 효소들 의 3-D 구조 및 유전자 규명 등의 심화된 연구가 지속되어 야 할 것으로 판단된다.
결 론
본 연구에서는 미생물(Sporosarcina sp. KM-01, KM-07, KM-12)과 식물(Canavalia ensiformis)이 생성하는 요소 분해효소에 의해 침전되는 탄산칼슘(CaCO3)의 광물학적 특성을 관찰하고 비교하였다. 박테리아에 의해 침전되는 CaCO3은 바테라이트였으며, 식물에 의해 생성되는 CaCO3
의 주요한 결정 상태는 방해석으로 확인되었고, 일부 바테 라이트가 검출되었다. 본 결과는 X-선회절분석(XRD) 및 전자현미경(SEM-EDS) 분석을 통해 CaCO3침전 형태 및 결정상의 차이를 확인했으며, 시간에 따라 침전물의 양이 증가하는 경향성을 확인하여 중금속 안정화, 지반고결화 등 향후 다양한 환경 연구 분야에 활용될 수 있음을 제시하 였다. 박테리아 및 식물 요소분해효소에 의해 생성되는 CaCO3의 결정 생성의 차이는 유사한 각 효소의 활성 대비 그 활성이 미치는 영향력의 범위 및 조건이 상이하여 결정 형태의 차이가 나타날 가능성이 있다고 유추된다. 개별적 으로 차이가 발생하는 조건은 반응에 관여하는 효소로부터 의 아미노산의 차이, 박테리아의 경우 해당 균주가 지니고 있는 기타 유전자로부터 발현되는 기타 단백질의 영향 등 을 포함할 수 있다. 또한, 박테리아의 경우 본 연구에서 활 용한 균주들의 분리 장소가 산성 조건임을 감안하여 해당 서식처에서 진화적으로 잘 적응된 특정 요인에 의해 분비 하는 요소분해효소 특이적 활성 및 영향성의 차이로 바테 라이트가 생성되고 아직 명확하게 규명되지 않은 바테라이 트 결정 구조 전이 형태와 특정 아미노산의 영향으로 해당 구조가 유지되었을 가능성을 예상할 수 있다. 결정 형태의 차이는 있으나 본 연구에서 관찰한 생물학적인 CaCO3침 전 기법은 기타 중금속을 동시에 침전할 수 있어 다양한 환 경 연구 분야에 적용할 수 있고 오염된 토양을 in-situ로 정 화하는 방법으로 활용될 수 있는 가능성을 제시한다. 또한, 생지화학적인 측면에서 금속이온과 결합하여 광물질을 형 성하는 요소분해효소의 활성은 Biomineralization 측면에 서 주요한 연구 분야로 인식되며, 이와 같은 Biomineral은 무기적으로 생성된 광물과 비교하여 독특한 크기, 결정도, 동위원소와 미량원소를 포함하는 등의 특성을 가져 다양한 분야에서 활용될 수 있다.
사 사
본 연구는 한국지질자원연구원의 주요사업(16-3414)의 지원으로 수행되었습니다.
References
Achal, V., Pan, X. and Zhang, D., 2011, “Remediation of copper-contaminated soil by Kocuria flava CR1, based on microbially induced calcite precipitation,” Ecol. Eng., Vol.
37, No. 10, pp. 1601-1605.
Bremner, J.M. and Douglas, L.A., 1971, “Inhibition of urease activity in soil,” Soil Biol. Biochem., Vol. 3, No. 4, pp. 297-307.
Chen, Y.Y., Clancy, K.A. and Burne, R.A., 1996, “Streptococcus salivarius urease: genetic and biochemical characterization and expression in a dental plaque Streptococcus,” Infect Immun., Vol. 64, No. 2, pp. 585-592.
DeJong, J.T., Fritzges, M.B. and Nüslein, K., 2006, “Microbially induced cementation to control sand response to undrained shear,” J. Geotech. Geoenviron. Eng., Vol. 132, No. 11, pp.
1381-1392.
De Koning-Ward T.F., Ward, A.C. and Robins-Browne, R.M., 1994, “Characterization of the urease-encoding gene complex of Yersinia enterocolitica,” Gene, Vol. 145, No. 1, pp. 25-32.
De Muynck, W., De Belie, N. and Verstraete, W., 2010,
“Microbial carbonate precipitation in construction materials:
A review,” Ecol. Eng., Vol. 36, No. 2, pp. 118-136.
Grasham, T.L.T. and Sheridan, P.P., 2007, “Design and validation of ureC-based primers for groundwater detection of urea-hydrolyzing bacteria,” Geomicrobiol. J., Vol. 24, No. 3, pp. 353-364.
Gu, C.H., Victor, Y.J., David, J. and Grant, W., 2001,
“Polymorph Screening: Influence of Solvents on the Rate of Solvent-mediated Polymorphic Transformation,” J. Pharm.
Sci., Vol. 90, No. 11, pp. 1878-1890.
Jones, B.D. and Mobley, H.L., 1989, “Proteus mirabilis urease: nucleotide sequence determination and comparison with Jack bean urease,” J. Bacteriol., Vol. 171, No. 12, pp.
6414-6422.
Kabalah-Amitai, L., Mayzel, B., Kauffmann, Y., Fitch, A.N., Bloch, L., Gilbert, P.U.P.A. and Pokroy, B., 2013, “Vaterite crystals contain two interspersed crystal structures,” Science, Vol. 340, pp. 454-457.
Kawasaki, S., Ogata, S., Hiroyoshi, N., Tsunekawa, M., Kaneko, K. and Terajima, R., 2010, “Effect of temperature on preci- pitation of calcium carbonate using soil microorganisms,” J.
Japan Soc. Eng. Geol., Vol. 51, No. 1, pp. 10-18.
Kim, K.J., No, A.R. and Park, K.S., 2009, “Isolation and
identification of urease-positive Photobacterium sp. strain HA-2 from sea water,” Kor. J. Fish Aquat. Sci., Vol. 42, No.
6, pp. 531-536.
Koper, T.E., El-Sheikh, A.F., Norton, J.M. and Klotz, M.G., 2004, “Urease-encoding genes in ammonia-oxidizing bacteria,”
Appl. Environ. Microbiol., Vol. 70, No. 4, pp. 2342-2348.
Labigne, A., Cussac, V. and Courcoux, P., 1991, “Shuttle cloning and nucleotide sequences of Helicobacter pylori genes responsible for urease activity,” J. Bacteriol, Vol.
173, No. 6, pp. 1920-1931.
Lee, M.N. and Park, H.D., 2012, “Isolation and characterization of acidophilic yeasts producing urease from Korean traditional Nuruk,” Korean J. Food Preserv., Vol. 19, No. 2, pp. 308-314.
Manoli, F. and Dalas, E., 2001, “Calcium carbonate crystallization in the presence of Glutamic Acid,” J. Cryst. Growth, Vol.
222, pp. 293-297.
Min, S.H. and Lee, M.H., 2007, “Purification and characterization of the Staphylococcus epidermidis urease,” J. Life Sci., Vol.
17, No. 4, pp. 581-586.
Mitchell, J.K. and Santamarina, J.C., 2005, “Biological considerations in geotechnical engineering,” J. Geotech.
Geoenviron. Eng., Vol. 131, No. 10, pp. 1222-1233.
Mobley, H.L. and Hausinger, R.P., 1989, “Microbial ureases:
significance, regulation, and molecular characterization,”
Microbial. Rev., Vol. 53, No. 1, pp. 85-108.
Mulrooney, S.B. and Hausinger, R.P., 1990, “Sequence of the Klebsiella aerogenes urease genes and evidence for accessory proteins facilitating nickel incorporation,” J.
Bacteriol., Vol. 172, No. 10, pp. 5837-5843.
Nam, I.H., Kim, Y.M., Murugesan, K., Jeon, J.R., Chang, Y.Y.
and Chang, Y.S., 2008, “Bioremediation of PCDD/Fs- contaminated municipal solid waste incinerator fly ash by a potent microbial biocatalyst,” J. Hazard. Mater., Vol. 157, pp. 114-121.
Nam, I.H., Kim, Y.M., Schmidt, S. and Chang, Y.S., 2006,
“Biotransformation of 1,2,3-tri- and 1,2,3,4,7,8-hexachlo- rodibenzo-p-dioxin by Sphingomonas wittichii strain RW1,” Appl. Environ. Microbiol., Vol. 72, pp. 112-116.
Nam, I.H., Roh, S.B., Park, M.J., Chon, C.M., Kim, J.G., Jeong, S.W., Song, H. and Yoon, M.H., 2016, “Immobilization of heavy metal contaminated mine wastes using Canavalia ensiformis extract,” Catena, Vol. 136, pp. 53–58.
Okwadha, G.D. and Li, J., 2010, “Optimum conditions for microbial carbonate precipitation,” Chemosphere, Vol. 81, No. 9, pp. 1143-1148.
Park, J.W., Kim, J.S., Ahn, J.W. and Han, C., 2006, “A Study on characteristics of precipitated calcium carbonate prepared by the nozzle spouting method,” J. Korean Ind.
남 인 현
2000년 성균관대학교 유전공학과 이학사 2002년 포항공과대학교 환경공학부 공학 2006년 포항공과대학교 환경공학부 공학석사
박사
현재 한국지질자원연구원 지질환경연구본부 책임연구원 (E-mail; [email protected])
김 재 곤
1988년 연세대학교 지질학과 이학사 1994년 The University of Georgia 이학 1998년 Texas A&M 이학박사석사
현재 한국지질자원연구원 지질환경연구본부 책임연구원 (E-mail; [email protected])
전 철 민
1994년 연세대학교 지질학과 이학사 1998년 연세대학교 지구시스템과학과 이 2003년 연세대학교 지구시스템과학과 이학석사
학박사
현재 한국지질자원연구원 지질환경연구본부 책임연구원 (E-mail; [email protected])
Eng. Chem., Vol. 17, No. 1, pp. 67-72.
Park, K.S., Iida, T., Yamaichi, Y., Oyagi, T., Yamamoto, K.
and Honda, T., 2000, “Genetic characterization of DNA region containing the trh and ure genes of Vibrio parahae- molyticus,” Infect Immun., Vol. 68, No. 10, pp. 5742-5748.
Park, M.J., Yoon, M.H. and Nam, I.H., 2014, “Characterization of urease-producing bacteria isolated from heavy metal contaminated mine soil,” Korean J. Soil Sci. Fert., Vol. 47, No. 6, pp. 391-397.
Park, S.S., Kim, W.J. and Lee, J.C., 2011, “Effect of biomineralization on the strength of cemented sands,” J.
Korean Geotech. Soc., Vol. 27, No. 5, pp. 75-84.
Roh, S.B., Park, M.J., Chon, C.M., Kim, J.G., Song, H., Yoon, M.H. and Nam, I.H., 2015, “Bioremediation of heavy metal contaminated mine wastes using urease based plant extract,”
J. Soil Groundw. Environ., Vol. 20, No. 1, pp. 56-64.
Shivkumara, C., Singh, P., Gupta, A. and Hegde, M.S., 2006,
“Synthesis of vaterite CaCO3 by direct precipitation using glycine and L-alanine as directing agents,” Mater. Res. Bull., Vol. 41, No. 8, pp. 1455-1460.
Sondi, I. and Salopek-Sondi, B., 2005, “Influence of the primary structure of enzymes on the formation of CaCO3 polymorphs: A comparison of plant (Canavalia ensiformis) and bacterial (Bacillus pasteurii) ureases,” Langmuir, Vol.
21, pp. 8876-8882.
Song, S.M., Seong, B.I., Koo, J.H. and Kim, I.H., 2011, “Effect
of Aspartic Acid and Lysine on Polymorphism of Calcium Carbonate Crystal Formed by Gas-Liquid Reaction,”
Korean Chem. Eng. Res., Vol. 49, No. 1, pp. 109-113.
Tabatabai M.A., 1973, “Michaelis constants of urease in soils and soil fraction,” Soil Sci. Soc. Am. Proc., Vol. 37, No. 5, pp. 701-707.
Terajima, R., Shimada, S., Oyama, T. and Kawasaki, S., 2009,
“Fundamental study of siliceous biogrout for eco-friendly soil improvement,” J. Japan Soc. Civil Eng. C., Vol. 65, No.
1, pp. 120-130.
Van Paassen, L.A., Ghose, R., Linden, T.J.M. van der, Star, W.R.L. van der, and Loosdrecht, M.C.M. van, 2010,
“Quantifying biomediated ground improvement by ureolysis:
large-scale biogrout experiment,” J. Geotech. Geoenviron.
Eng., Vol. 136, No. 12, pp. 1721-1728.
Whiffin, V.S., Paassen, L.A. van, and Harkes, M.P., 2007,
“Microbial carbonate precipitation as a soil improvement technique,” Geomicrobiol, J. Vo. 24, No. 5, pp. 417-423.
Xie, A.J., Shen, Y.H., Zhang, C.Y., Yuan, Z.W., Zhu, X.M.
and Yang, Y.M., 2005, “Crystal growth of calcium carbonate with various morphologies in different amino acid systems,”
J. Cryst. Growth, Vol. 285, pp. 436-443.
Young, G.M., Amid, D. and Miller, V.L., 1996, “A bifunctional urease enhances survival of pathogenic Yersinia enterocolitica and Morganella morganii at low pH,” J. Bacteriol., Vol.
178, No. 22, pp. 6487-6495.