신두리 해안사구에 자생하는 사구식물 내생진균의 다양성 분석
유영현1†, 서영교1†, 윤혁준1, 김현1, 김예은1, 이리나 할무라토바1, 임순옥1, 김창무2, 김종국1*
1경북대학교생명과학부
2국립생물자원관미생물자원과
Received : May 20, 2013 / Revised : June 25, 2013 / Accepted : June 26, 2013
서 론
신두리해안사구는충청남도태안군원북면에위치하고있 으며, 생태학적으로매우중요한지형으로알려지게되어서 2001년에천연기념물 431호로지정되었다. 신두리해안사구 의규모는해안을따라북서방향으로약 3.4 km 길이와폭 은 0.2~1.3 km 정도이고, 면적은 2 km2로알려져있다[12].
해안사구(Coastal sand dune)는해류와연안류및바람에의 해모래가퇴적작용으로형성되어해안선을따라쌓인모래 언덕을말하며, 강한바람, 강한일조량, 물부족, 염도등의 열악한환경조건을가진다. 그리고해안사구는해양생태계 와육상생태계의이행대(Ecotone)를형성하고[3, 12, 21], 이 러한독특한환경지형으로서해안사구식물(Coastal sand-
dune plant)이분포하고자생하는식생으로이루어져있다. 해안사구식물은주로우리나라동해안의고래불해안사구 와서해안태안반도에위치한신두리해안사구등에서다양 하고많은식물종이자생하고있는것으로알려져있으며, 생태적자원과관광자원으로서매우중요시되고있다. 그러 나최근에는해안사구지역의훼손, 오염및난개발등으로 인하여수많은사구지역이사라져가고있는추세이고[21], 전세계적으로환경보전및복원에대하여관심을받고있으 며[1, 3], 생태학적으로이행대를형성하고있기때문에특이 한생물종의생태학적, 경제적·학문적및여러측면에서연
구와관심이집중되고있다[21]. 최근에는해안사구지역의
보존및복원을위하여해안사구식물의발아, 개화및생장 에대한연구와식물과미생물간의상호관계에대한연구가 점차적으로이루어지고있으며[5-8, 25], 해안사구식물과공 생하는유용미생물의 gibberellin (GA), auxin (IAA) 같은식 물호르몬연구[4, 7, 8, 25], 미생물에의한식물의염스트레
스연구[20] 및새로운미생물자원의개발등으로해안사구
식물의생리가해안사구환경의보호를위하여중요하다는 Endophytic Fungal Diversity Associated with the Roots of Coastal Sand-dune Plants in the Sindu-ri Coastal Sand Dune, Korea. You, Young-Hyun1†, Yeonggyo Seo1†, Hyeokjun Yoon1, Hyun Kim1, Ye-Eun Kim1, Irina Khalmuratova1, Soon-Ok Rim1, Changmu Kim2, and Jong-Guk Kim1*. 1School of Life Science, Kyungpook National University, Daegu 702- 701, Korea, 2National Institute of Biological Resource, Incheon 404-708, Korea
The coastal sand-dune plants of eight species; Argusia sibirica, Calystegia soldanella, Elymus mollis, Lithospermum zollingeri, Raphanus sativus, Salsola collina, Zoysia macrostachya, and Zoysia sinica were collected from the Shindu-ri coastal sand dune. Ninety-eight endophytic fungal strains were isolated from the roots of these plants, analyzed, and identified by sequences in their internal transcribed spacers (ITS) at the ITS1, 5.8S, and ITS2 regions. The diversity of endophytic fungi iso- lated from coastal sand-dune plants was confirmed with various diversity indices. The fungal strains belonged to thirteen orders: Capnodiales (3.09%), Eurotiales (70.10%), Glomerellales (1.03%), Helotiales (3.09%), Hypocreales (9.28%), Mortierel- lales (2.06%), Onygenales (1.03%), Ophiostomatales (1.03%), Pleosporales (1.03%), Polyporales (1.03%), Russulales (1.03%), Saccharomycetales (2.06%), and Xylariales (1.03%). Of the endophytic fungal strains collected, Penicillium (59.18%
in Eurotiales) and Fusarium (5.10% in Hypocreales) were the most abundant in coastal sand-dune plants. The endophytic fun- gal strains isolated from C. soldanella were more diverse compared to strains from the other coastal sand-dune plants.
Keywords: Coastal sand-dune plant, endophytic fungi, Shindu-ri sand dune, species richness, species diversity index
*Corresponding author
Tel: +82-53-950-5379, Fax: +82-53-955-5379 E-mail: [email protected]
†These authors contributed equally to this work.
© 2013, The Korean Society for Microbiology and Biotechnology
것이알려지고있다[8, 21, 25]. 그리고다양한식물과공생 하는내생진균에대한다양성연구및특성에대한연구들 이이루어지고있다[5, 6, 17, 24].
그러나몇몇의해안사구지역을대상으로해안미생물에관 한연구는토양으로부터분리된원핵미생물의군집분석등 이다소이루어져있지만, 신두리해안사구에자생하는해안 사구식물과상호관계에있는식물뿌리내생진균(Endophytic fungi)의연구는거의이루어져있지않다[14, 25].
본연구는우리나라최대의해안사구지역으로알려진신 두리해안사구에자생하고있는 8종의해안사구식물과공생 하는내생진균을분리및동정하였다. 그리고신두리해안사 구식물내생진균류를분류학적으로분석하였고, 내생진균 의종다양성을확인하기위하여, 6종류의다양한지수를적 용하여 종 풍부도(Species richness) 및 종 다양성 지수 (Species diversity index)를분석하였다.
재료 및 방법
식물시료의 채집
본연구는충청남도태안군신두리해안사구에자생하는 8종의해안사구식물을채취하였다. 채집한해안사구식물은 모래지치(Argusia sibirica), 갯메꽃(Calystegia soldanella), 갯그령(Elymus mollis), 반디지치(Lithospermum zollingeri), 갯무(Raphanus sativus), 솔장다리(Salsola collina), 왕잔디 (Zoysia macrostachya) 및갯잔디(Zoysia sinica)이며, 신두리 해안사구의다양한지역에서식물샘플들을채집하여실험재 료로사용하였다(Table 1). 그리고해안사구식물들은식물종 에따라식물코드(Plant code)로분류하여본연구에사용하 였다.
내생진균의 분리 및 배양조건
신두리해안사구에서채집한해안사구식물의뿌리의토양
을세척하고, 계면활성제(Tween 80, Duksan, South Korea) 를 10분간 2회 처리한 후에 멸균증류수(Sterile distilled water)로세척하였다. 그리고과염소산(Perchloric acid, 1%, Duksan, South Korea)을 10분간 2회처리한후, 멸균증류 수로 세척하여 수분을 제거하였으며, 식물 뿌리시료를 3~4 cm 길이로절단하여실험재료로사용하였다[24, 25]. 식 물뿌리시료로부터내생진균을분리하기위하여, 스트렙토마 이신(Streptomycin, Sigam-aldrich, USA) 80 ppm이 함유된 Hagem minimal (HM) 배지를사용하였고, 식물뿌리시료를 최소배지에올려 25oC 조건에서배양하였다[7, 19, 23, 25].
그리고식물뿌리시료단면에서내생진균류를 toothpicking 하였고, HM 배지에획선도말법(Streaking)을이용하여다 시 25oC 조건에서배양하였다. 그리고식물뿌리내생진균을 potato dextrose agar (PDA, Difco, USA)에계대배양하였고, 균사의형태, 색상등을확인하고선별하여순수분리하였다
[24, 25]. 해안사구식물의뿌리에서순수분리한내생진균류
의 동정을 위하여, potato dextrose broth (PDB, Difco, USA)에서 7일동안 120 rpm으로 25oC 조건하에서진탕배양 (shake culture) 하였으며, 여과지를사용하여내생진균의균 체(mycobiont)를동결건조하여동정을위한시료로사용하 였다[24, 25].
Genomic DNA의 추출과 PCR 조건
신두리해안사구에자생하는해안사구식물의뿌리로부터 분리된내생진균류의동정으로 ITS 영역의염기서열을확인 하기위하여, 동결건조상태의내생진균류의균체를 DNeasy Plant mini kit (QIAgen, Valencia, CA, USA)를사용하여 genomic DNA를추출하여 PCR의주형으로사용하였다[5, 7]. Genomic DNA로부터 rDNA-ITS 영역을증폭하기위해 ITS1과 ITS4 primer를 사용하였다[9, 10, 24, 25]. 그리고 내생진균의 동정을 위하여, polymerase chain reaction (PCR) 조건은 최종농도 Tris-HCl 10 mM (pH 8.5), MgCl2
Table 1. Geographic coordinates of the native plants in coastal sand dune.
Plant taxon
(Family) Scientific name of plantsa Plant
code Site of collection Originally reported habitatb Boraginaceae Argusia sibirica (L.) Dandy Asd Taean, Chungnam, Korea Coastal sand dune Convolvulaceae Calystegia soldanella (L.) Roem. & Schultb Cs Taean, Chungnam, Korea Coastal sand dune
Gramineae Elymus mollis Trin. Em Taean, Chungnam, Korea Coastal sand dune
Boraginaceae Lithospermum zollingeri A.DC. Lz Taean, Chungnam, Korea Coastal sand dune Cruciferae Raphanus sativus var. raphnistroides Makino Rs Taean, Chungnam, Korea Coastal sand dune Chenopodiaceae Salsola collina Pall. Sc Taean, Chungnam, Korea Coastal sand dune Gramineae Zoysia macrostachya Franch. & Sav. Zm Taean, Chungnam, Korea Coastal sand dune
Gramineae Zoysia sinica Hance Zs Taean, Chungnam, Korea Coastal sand dune
aScientific name of native plants in coastal sand dune.
bThe habitat of native plants in coastal sand dune.
1.5 mM, dNTPs 200 nM, KCl 40 mM이고, 10 pmol의 primer와 0.1 unit의 Ex-Taq DNA polymerase (Takara, Japan)를 사용하였다. PCR 반응조건은 (95oC, 2 min) predenaturation, (95oC, 30 sec) denaturation, (49.5-54.5oC, 1 min) annealing, (72oC, 1 min) extension, (35 cycles) total cycles, (72oC, 7 min) final extension으로수행하였고, 증폭 된 PCR 산물은 1.5% agarose gel에서 전기영동 후, ethidium bromide (EtBr, Sigam-aldrich, USA)을사용하여 20분염색한후, UV transilluminator로전기영동밴드를확 인하였다[24, 25]. PCR 반응후, 전기영동단편들은 QIAquick PCR purification kit (Qiagen Inc., Germany)를사용하여 정제하였으며, ABI PRISM BigDye Terminator Cycle Sequencing Kit (PE Biosystems, Foster City, CA, USA) 를사용하여염기서열결정반응을수행하였다. 반응 DNA 단편들은 ABI 310 DNA sequencer (Perkin Elmer, Foster City, CA, USA)를사용하여염기서열을결정하였다[24].
식물뿌리 내생진균류의 계통학적 분석 및 다양성 분석 해안사구식물의뿌리로부터분리된내생진균의염기서열 을 ClustalX 프로그램으로다중정렬을수행한후, Lasergene 7과 BioEdit 프로그램을사용하여염기서열을확인하였다. 그리고 내생진균의 계통학적 분석은 MEGA 5.1프로그램 Neighbor-Joining (NJ: 1000 bootstrap replicates) 방법을 이용하여분석하였으며[18], 내생진균의유전적인동정을위
하여 rDNA-ITS 영역의 염기서열을 NCBI의 nucleotide blast 데이터와비교하여확인하였다[24].
태안군신두리해안사구에자생하는해안사구식물에따른 내생진균의다양성분석을위하여동정된내생진균류의속 (Genus)을확인하였다. 그리고내생진균들의종풍부도와종 다양성지수를적용하여비교하였다. 종풍부도는 Mehinick’s index (Dmn) [22]와 Margalef's richness (Dmg) [13]를적용 하여분석하였고, 종다양성지수는 Fisher’s alpha index (α) [2], Shannon’s diversity index (H’) [11, 15, 17], Simpson's index (D) [11, 16], Simpson's diversity index (1-D) [11, 16]
를적용하여분석하였다.
결 과
사구식물 내생진균의 동정 및 계통학적 분석
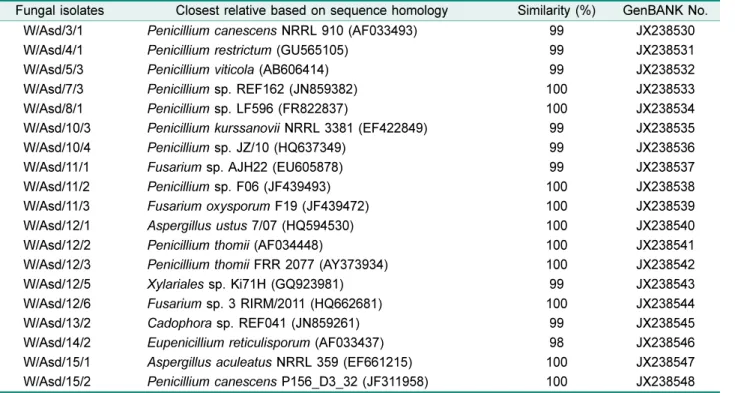
태안군신두리해안사구에자생하고있는 8종의해안사구 식물의뿌리로부터 254주를분리하였고, 형태적으로다른내 생진균 98주를최종선별하였다. 그리고해안사구식물로부 터분리된균주는 모래지치로부터 19주, 갯메꽃으로부터 11주, 갯그령으로부터 8주, 반디지치로부터 18주, 갯무로부 터 9주, 솔장다리로부터 14주, 왕잔디로부터 15주, 그리고갯 잔디로부터 4주를선별하였다.
해안사구식물의뿌리로부터분리된내생진균의동정을수
행하였고, 각 식물의 내생진균 ITS 염기서열을 이용하여
Table 2. The identification of endophytic fungi isolated in this study.
Fungal isolates Closest relative based on sequence homology Similarity (%) GenBANK No.
W/Asd/3/1 Penicillium canescens NRRL 910 (AF033493) 99 JX238530
W/Asd/4/1 Penicillium restrictum (GU565105) 99 JX238531
W/Asd/5/3 Penicillium viticola (AB606414) 99 JX238532
W/Asd/7/3 Penicillium sp. REF162 (JN859382) 100 JX238533
W/Asd/8/1 Penicillium sp. LF596 (FR822837) 100 JX238534
W/Asd/10/3 Penicillium kurssanovii NRRL 3381 (EF422849) 99 JX238535
W/Asd/10/4 Penicillium sp. JZ/10 (HQ637349) 99 JX238536
W/Asd/11/1 Fusarium sp. AJH22 (EU605878) 99 JX238537
W/Asd/11/2 Penicillium sp. F06 (JF439493) 100 JX238538
W/Asd/11/3 Fusarium oxysporum F19 (JF439472) 100 JX238539
W/Asd/12/1 Aspergillus ustus 7/07 (HQ594530) 100 JX238540
W/Asd/12/2 Penicillium thomii (AF034448) 100 JX238541
W/Asd/12/3 Penicillium thomii FRR 2077 (AY373934) 100 JX238542
W/Asd/12/5 Xylariales sp. Ki71H (GQ923981) 99 JX238543
W/Asd/12/6 Fusarium sp. 3 RIRM/2011 (HQ662681) 100 JX238544
W/Asd/13/2 Cadophora sp. REF041 (JN859261) 99 JX238545
W/Asd/14/2 Eupenicillium reticulisporum (AF033437) 98 JX238546
W/Asd/15/1 Aspergillus aculeatus NRRL 359 (EF661215) 100 JX238547
W/Asd/15/2 Penicillium canescens P156_D3_32 (JF311958) 100 JX238548
Table 2. Continued.
Fungal isolates Closest relative based on sequence homology Similarity (%) GenBANK No.
W/Cs/1/1 Fusarium sp. 3 RIRM/2011 (HQ662681) 100 JX238562
W/Cs/1/2 Nectriaceae sp. CHTAE36 (JF773610) 96 JX238563
W/Cs/4/1 Arthrinium sacchari A09 (HQ115646.) 100 JX238564
W/Cs/6/1 Cladosporium sp. CPC 15516 (GU214631) 100 JX238565
W/Cs/8/1 Penicillium kojigenum NRRL 3442 (AF033489) 99 JX238566
W/Cs/10/1 Nectria mauritiicola (AJ558115) 96 JX238567
W/Cs/10/2 Penicillium montanense (AF527058) 100 JX238568
W/Cs/11/2 Oidiodendron chlamydosporicum (AF062789) 99 JX238569
W/Cs/12/1 Penicillium restrictum FRR 332 (AY373928) 99 JX238570
W/Cs/13/4 Talaromyces barcinensis CBS 649.95 (JN899349) 99 JX238571
W/Cs/13/4/1 Talaromyces helicus NRRL 2106 (AF033396) 99 JX238572
W/Em/2/1 Gibberella intermedia (AB587007) 100 JX238576
W/Em/3/2 Irpex lacteus 12 (HQ656793) 99 JX238577
W/Em/7/1 Nectria mauritiicola (AJ558115) 96 JX238578
W/Em/10/1 Penicillium corylophilum TBG1/14 (JQ082506) 100 JX238579
W/Em/10/1/1 Penicillium pinophilum SGE75 (JQ776546) 100 JX238580
W/Em/11/1 Penicillium solitum 20/01 (JN642222) 100 JX238581
W/Em/12/2 Geomyces sp. OUCMBIII091010 (HQ914918) 99 JX238582
W/Em/14/3 Leotiomycetes sp. (JQ761483) 95 JX238583
W/Lz/3/1 Penicillium kurssanovii NRRL 3381 (EF422849) 99 JX238589
W/Lz/3/3 Penicillium sp. EMA/2011f (JF429679) 99 JX238590
W/Lz/4/6 Penicillium viticola (AB606414) 99 JX238591
W/Lz/5/3 Mortierella sp. dd08032 (FJ810149) 100 JX238592
W/Lz/6/1 Penicillium quercetorum NRRL 3758 (AY443471) 98 JX238593
W/Lz/6/3 Peniophora sp. 9/2 (HM535362) 99 JX238594
W/Lz/6/4 Eupenicillium pinetorum NRRL 3008 (AF033411) 100 JX238595
W/Lz/6/5 Penicillium montanense (AF527058) 100 JX238596
W/Lz/7/1 Penicillium restrictum (GU565105) 99 JX238597
W/Lz/7/2 Mortierella sp. dd08032 (FJ810149) 100 JX238598
W/Lz/10/1 Penicillium asperosporum (AF033412) 100 JX238599
W/Lz/10/2 Sporothrix sp. 1/CMW9488 (AY546720) 98 JX238600
W/Lz/11/2 Cadophora sp. 6e51/2 (DQ317329) 99 JX238601
W/Lz/14/2 Penicillium restrictum FRR 332 (AY373928) 99 JX238602
W/Lz/14/5 Penicillium purpurascens NRRL 720 (AF033408) 99 JX238603
W/Lz/15/1 Penicillium glabrum KUC1553 (HM469402) 99 JX238604
W/Lz/15/4 Penicillium purpurascens KUC1788 (HM469419) 99 JX238605
W/Lz/16/1 Penicillium restrictum NRRL 25744 (AF033459) 99 JX238606
W/Rs/3/1 Cladosporium sp. SS/S12 (GU797141) 100 JX238615
W/Rs/3/2 Penicillium sp. FGTR9 (JF689825) 99 JX238616
W/Rs/7/2 Penicillium viridicatum CBS 390.48 (JN942697) 100 JX238617
W/Rs/7/4 Penicillium pinophilum SGE75 (JQ776546) 100 JX238618
W/Rs/9/1/1 Penicillium multicolor KUC1626 (HM469407) 99 JX238619
W/Rs/9/2 Paraconiothyrium sporulosum 248 (FJ224106) 98 JX238620
W/Rs/10/3 Penicillium polonicum NRRL 995 (AF033475) 100 JX238621
W/Rs/12/1 Penicillium pinophilum NRRL 58691 (GQ337428) 99 JX238622
W/Rs/12/2 Penicillium sp. MS/2011/F37 (HE608803) 99 JX238623
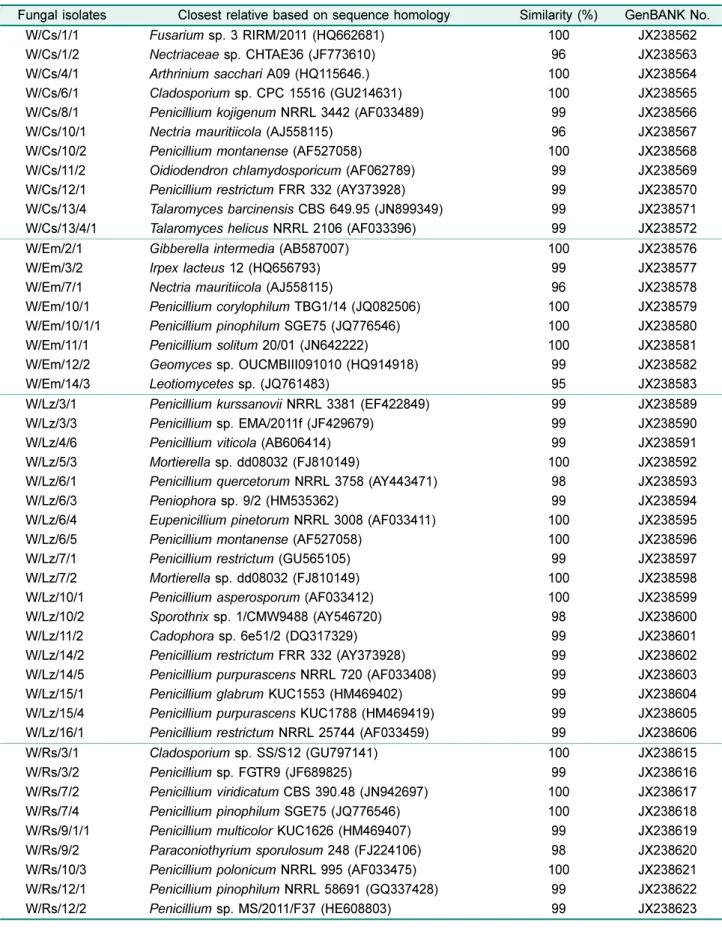
NCBI의 GenBank에서 blast search를수행하였다. 그결과, 가장 유의성이 높은 균주들을 확인하였으며 염기서열을 GenBank에등록하여, 모래지치(JX238530-JX238548), 갯메 꽃(JX238562-JX238572), 갯그령(JX238576-JX238583), 반 디지치(JX238589-JX238606), 갯무(JX238615- JX238623), 솔장다리(JX238624-JX238637), 왕잔디(JX238667- JX238681), 그리고갯잔디(JX238682-JX238685)로부터분리된균주들 의 accession number를제공받았다(Table 2).
해안사구식물로부터분리된내생진균은모래지치에서분
리된내생진균은자낭균문(Ascomycota)에속하는 Eurotio- mycetes강(Class)에속하는 Eurotiales목(Order)의 Aspergillus 속, Eupenicillium속, Penicillium속과 Leotiomycetes강에 속하는 Helotiales목의 Cadophora속, 그리고 Sordariomycetes 강에 속하는 Hypocreales목의 Fusarium속과 Xylariales 목의 Xylariales속이분석되었다. 갯메꽃에서분리된내생 진균은 자낭균문에속하는 Dothideomycetes강에속하는 Capnodiales목의 Cladosporium속과 Eurotiomycetes강 에속하는 Eurotiales목의 Penicillium속, Talaromyces속 Table 2. Continued.
Fungal isolates Closest relative based on sequence homology Similarity (%) GenBANK No.
W/Sc/1/4 Meyerozyma guilliermondii PH5 (JN974767) 99 JX238624
W/Sc/1/5 Paecilomyces lilacinus NRRL896 (GU980017) 99 JX238625
W/Sc/2/2 Penicillium paneum Z8M/2 (JN226928) 99 JX238626
W/Sc/2/5 Talaromyces trachyspermus Ln/11/1 (EU287809) 100 JX238627
W/Sc/3/1 Penicillium paneum M/17 (JQ422609) 100 JX238628
W/Sc/3/2 Penicillium minioluteum E3 (GU566240) 98 JX238629
W/Sc/4/1 Penicillium paneum SZ8M/15 (JN226992) 100 JX238630
W/Sc/4/2 Cladosporium silenes CMT48 (JQ754032) 100 JX238631
W/Sc/5/1 Penicillium brasilianum KUC1433 (HM469396) 100 JX238632
W/Sc/5/2 Plectosphaerella cucumerina (JQ796755) 99 JX238633
W/Sc/8/1 Penicillium paneum M/18 (JQ422610) 98 JX238634
W/Sc/9/2 Penicillium samsonii CBS 137.84 (JN899369) 98 JX238635
W/Sc/12/1 Aspergillus ochraceopetaliformis (JQ647894) 99 JX238636
W/Sc/13/3 Penicillium multicolor KUC1626 (HM469407) 99 JX238637
W/Zm/1/3 Penicillium verruculosum AT010 (HQ607791) 99 JX238667
W/Zm/5/1 Penicillium verruculosum AT010 (HQ607791) 99 JX238668
W/Zm/6/1 Penicillium pinophilum CB40 (JQ228237) 99 JX238669
W/Zm/6/3 Lachnum sp. 1 MV/2011 PB 003 (JN655650) 98 JX238670
W/Zm/7/1 Paecilomyces lilacinus MY543 (GU980026) 100 JX238671
W/Zm/8/1 Meyerozyma guilliermondii GK8 (JF508433) 100 JX238672
W/Zm/9/1 Penicillium verruculosum ATT281 (HQ607919) 99 JX238673
W/Zm/10/3 Penicillium verruculosum PTC06 (JF682635) 99 JX238674
W/Zm/11/1 Penicillium aculeatum LP67 (HQ392497) 98 JX238675
W/Zm/12/1 Penicillium sp. 22/M/5 (EU076945) 99 JX238676
W/Zm/12/2 Penicillium pinophilum CB42 (JQ228239) 99 JX238677
W/Zm/13/1 Penicillium aculeatum F1001 (HQ647326) 99 JX238678
W/Zm/14/1 Penicillium sclerotiorum FRR 1202 (AY373931) 99 JX238679
W/Zm/14/2 Penicillium samsonii CBS 137.84 (JN899369) 98 JX238680
W/Zm/15/3 Penicillium pinophilum SGE75 (JQ776546) 100 JX238681
W/Zs/3/1 Penicillium urticae JZ/56 (HQ637345) 99 JX238682
W/Zs/10/1 Talaromyces helicus NRRL 2106 (AF033396.1) 99 JX238683
W/Zs/14/1 Fusarium oxysporum BWH/F (JQ301897) 100 JX238684
W/Zs/15/1 Penicillium thomii (AF034460) 100 JX238685
Fig. 1. Phylogenetic relationships among sixty-nine endophytic fungi from coastal sand-dune plants belonging to Eurotio- mycetes (Ascomycota).
Phylogenetic tree was generated from the rDNA-ITS sequences of endophytic fungi using Mega program (version: 5.1). Bootstrap values (≥50%) are indicated at relevant nodes.
과 Leotiomycetes강에속하지만분류체계가명확하지 않 은 Incertae sedis목으로 분류되는 Oidiodendron속과 Sordariomycetes강에속하는 Hypocreales목의 Fusarium 속, Nectriaceae속과 분류체계가 명확하지 않은 Incertae sedis목으로분류되는 Arthrinium속이분석되었다. 갯그령 에서분리된내생진균은자낭균문에속하는 Eurotiomycetes 강에 속하는 Eurotiales목의 Nectria속, Penicillium속과 Onygenales 목의 Geomyces속과 Leotiomycetes강에속하 지만분류체계가 명확하지않은 Incertae sedis목으로 분 류되는 Leotiomycetes속과 Sordariomycetes강에 속하는 Hypocreales목의 Gibberella속, 그리고담자균문(Basidiomycota) 에 속하는 Agaricomycetes강에 속하는 Polyporales목의
Irpex속이분석되었다. 반디지치에서분리된내생진균은자
낭균문에속하는 Eurotiomycetes강에속하는 Eurotiales목
의 Eupenicillium속, Penicillium속과 Sordariomycetes강 에속하는 Ophiostomatales목에속하는 Sporothrix속이확 인되었으며, 분류체계가명확하지않은 Incertae sedis강에 속하는 Mortierellales목의 Mortierella속이확인되었다. 그 리고 담자균문에 속하는 Agaricomycetes강에 속하는 Russulales목의 Peniophora속이확인되었다. 갯무에서분 리된내생진균은자낭균문에속하는 Dothideomycetes강에 속하는 Capnodiales목의 Cladosporium속과 Pleosporales 목의 Paraconiothyrium속과 Eurotiomycetes강에 속하는 Eurotiales목의 Penicillium속이확인되었다. 솔장다리에서 분리된내생진균은자낭균문에속하는 Dothideomycetes강 에속하는 Capnodiales목의 Cladosporium속과 Eurotiomycetes 강에 속하는 Eurotiales목의 Aspergillus속, Penicillium속, Talaromyces속이확인되었고, Saccharomycetes강에속하는
Fig. 2. Phylogenetic relationships among twenty-nine endophytic fungi from coastal sand-dune plants belonging to Agarico- mycetes (Basidiomycota), Dothideomycetes, Saccharomycetes, Sordariomycetes, and incertae sedis (Ascomycota).
Bootstrap values (≥50%) are indicated at relevant nodes.
Saccharomycetales목의 Meyerozyma속과 Sordariomycetes강 에속하는 Hypocreales목의 Paecilomyces속과 Glomerellales목 의 Plectosphaerella속이확인되었다. 왕잔디에서 분리된 내생진균은 Eurotiomycetes강에 속하는 Eurotiales목의 Penicillium속과 Leotiomycetes강에속하는 Helotiales목의 Lachnum속과 Saccharomycetes강에 속하는 Saccharo- mycetales목의 Meyerozyma속과 Hypocreales목의 Paecilo-
myces속이확인되었다. 그리고갯잔디에서분리된내생진균
은 Eurotiomycetes강에속하는 Eurotiales목의 Penicillium속, Talaromyces속과 Sordariomycetes강에속하는 Hypocreales 목에속하는 Fusarium속이확인되었다. 본연구에서분리 및동정된내생진균들의유연관계를확인하기위하여계통
수를작성하여확인하였다. 작성된계통수는가장많은비율 을나타내고있는자낭균문에속하는 Eurotiomycetes강의 Eurotiales목과 그리고 Eurotiales목과가까운유연관계인 Onygenales목에속하는내생진균을나타내었고(Fig. 1), 분 리된 내생진균 중에서 비율이 낮은 Dothideomycetes강, Leotiomycetes강, Saccharomycetes강, Sordariomycetes강 및담자균문에속하는 Agaricomycetes강에속하는다양한 목에속하는내생진균에대하여계통수를확인하였다(Fig. 2).
신두리 해안사구식물의 내생진균 다양성 분석
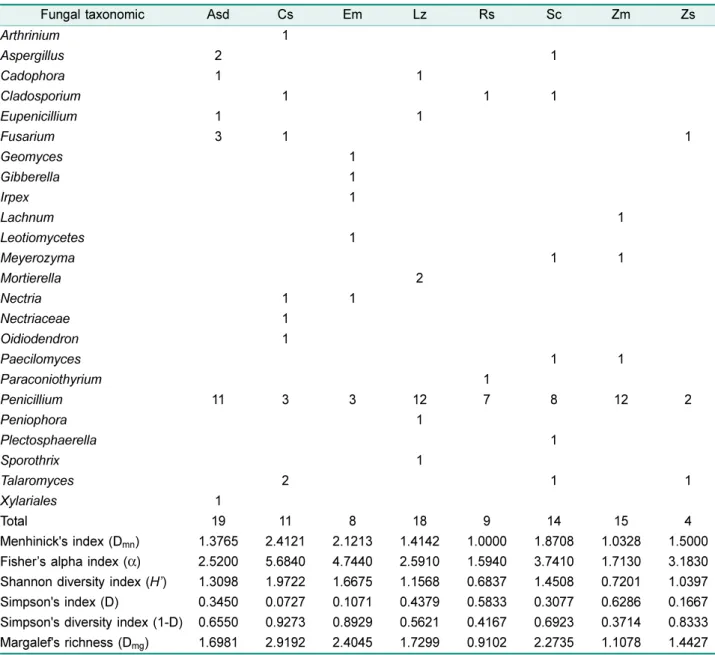
해안사구에자생하는해안사구식물의뿌리로부터분리된내 생진균의유전학적다양성을확인하기위하여, 분리된모든내
Table 3. Diversity and distribution of endophytic fungi isolated from the roots of native plants in coastal sand dune.
Fungal taxonomic Asd Cs Em Lz Rs Sc Zm Zs
Arthrinium 1
Aspergillus 2 1
Cadophora 1 1
Cladosporium 1 1 1
Eupenicillium 1 1
Fusarium 3 1 1
Geomyces 1
Gibberella 1
Irpex 1
Lachnum 1
Leotiomycetes 1
Meyerozyma 1 1
Mortierella 2
Nectria 1 1
Nectriaceae 1
Oidiodendron 1
Paecilomyces 1 1
Paraconiothyrium 1
Penicillium 11 3 3 12 7 8 12 2
Peniophora 1
Plectosphaerella 1
Sporothrix 1
Talaromyces 2 1 1
Xylariales 1
Total 19 11 8 18 9 14 15 4
Menhinick's index (Dmn) 1.3765 2.4121 2.1213 1.4142 1.0000 1.8708 1.0328 1.5000 Fisher’s alpha index (α) 2.5200 5.6840 4.7440 2.5910 1.5940 3.7410 1.7130 3.1830 Shannon diversity index (H’) 1.3098 1.9722 1.6675 1.1568 0.6837 1.4508 0.7201 1.0397 Simpson's index (D) 0.3450 0.0727 0.1071 0.4379 0.5833 0.3077 0.6286 0.1667 Simpson's diversity index (1-D) 0.6550 0.9273 0.8929 0.5621 0.4167 0.6923 0.3714 0.8333 Margalef's richness (Dmg) 1.6981 2.9192 2.4045 1.7299 0.9102 2.2735 1.1078 1.4427
생진균류를 확인하였으며, Asd에서 6속 9종(Unidentified species: 8 strains), Cs에서 8속 8종 (Unidentified species:
3 strains), Em에서 6속 6종(Unidentified species: 2 strains), Lz에서 6속 9종(Unidentified species: 6 strains), Rs에서 3속 5종(Unidentified species: 3 strains), Sc에서 7 속 11종, Zm에서 4속 7종(Unidentified species: 2 strains), Zs에서 3속 4종이확인되었다.
내생진균류의종풍부도를확인하기위하여 Mehinick’s index (Dmn)를적용하였을때, Asd (1.377), Cs (2.412), Em (2.121), Lz (1.414), Rs (1.000), Sc (1.871), Zm (1.033), Zs (1.500)로확인되었으며, Margalef's richness (Dmg) 다양성 지수의 적용은 Asd (1.700), Cs (2.920), Em (2.410), Lz (1.730), Rs (0.910), Sc (2.274), Zm (1.110), Zs (1.443)으로 확인되었다. 종다양성지수인 Fisher’s alpha index (α)를 적용하였을 때, Asd (2.520), Cs (5.684), Em (4.744), Lz (2.591), Rs (1.594), Sc (3.741), Zm (1.713), Zs (3.183)으로 확인되었다. Shannon’s diversity index (H’)의적용은 Asd (1.310), Cs (1.972), Em (1.668), Lz (1.157), Rs (0.684), Sc (1.451), Zm (0.720), Zs (1.040)으로확인되었다. Simpson's index (D)를적용하였을때는 Asd (0.345), Cs (0.073), Em (0.107), Lz (0.438), Rs (0.583), Sc (0.308), Zm (0.629), Zs (0.167)으로확인되었다. Simpson's diversity index (1-D)의 적용은 Asd (0.655), Cs (0.927), Em (0.893), Lz (0.562), Rs (0.417), Sc (0.693), Zm (0.371), Zs (0.833)으로확인되 었다(Table 3).
고 찰
본연구에서는우리나라 최대의해안사구로 알려진신두 리해안사구에자생하는해안사구식물들의내생진균에대 한연구로서, 해안사구식물과공생하고있는내생진균류를 분리및동정을수행하여분리된내생진균들의유연관계를 확인하기위하여계통분석을수행하였으며, 다양성지수를 적용하여사구식물 8종에따른내생진균의다양성을확인 하였다.
해안사구식물로부터분리된 모든내생진균은 2문, 6강,
13목으로분류가되었고, 24속으로분류되는것을확인하
였으며, 분류체계가명확하게연구되지않은 incertae sedis 에속하는내생진균들이포함되는것을할수있었다. 분리 된내생진균은크게자낭균문과담자균문으로분류되었고, Agaricomycetes강, Dothideomycetes강, Eurotiomycetes강, Leotiomycetes강, Saccharomycetes강, Sordariomycetes강 및분류체계가명확하지않은 incertae sedis강으로분류되 었다. 그리고내생진균의목수준에서분류는 Polyporales목, Russulales목, Capnodiales목, Pleosporales목, Eurotiales
목, Onygenales목, Mortierellales목, Helotiales목, Saccharo- mycetales목, Hypocreales목, Xylariales목, Ophiostomatales 목, Glomerellales목및분류체계가명확하지않은 incertae sedis목으로 확인되었다. 모든 내생진균은 Arthrinium 속(1.02%), Aspergillus속(3.06%), Cadophora속(2.04%), Cladosporium 속(3.06%), Eupenicillium속(2.04%), Fusarium 속(5.10%), Geomyces속(1.02%), Gibberella속(1.02%), Irpex 속(1.02%), Lachnum속(1.02%), Leotiomycetes속(1.02%), Meyerozyma속(2.04%), Mortierella속(2.04%), Nectria속 (2.04%), Nectriaceae속(1.02%), Oidiodendron속(1.02%), Paecilomyces속(2.04%), Paraconiothyrium속(1.02%), Pen- icillium속(59.18%), Peniophora속(1.02%), Plectosphaerella속 (1.02%), Sporothrix속(1.02%), Talaromyces속(4.08%), Xylariales속(1.02%)으로분류되는것을확인하였다. 분리 된내생진균중에서분류체계가명확하지않은 Mortierella 속, Arthrinium속, Oidiodendron속, Leotiomycetes속이 확인 되었으며, 내생진균류중에서종이확인되지않은 24균주가 확인되었다(Fig. 3).
내생진균류의종풍부도 Menhinick's index (Dmn)의적용 시에 Cs에서 2.412로가장높았고, Rs에서가장낮은수치 를 나타내었으며, Margalef's richness (Dmg)는 Cs에서 2.290으로가장높은지수와 Rs에서 0.910으로가장낮은지 수를나타내었다. 종다양성지수를확인하기위하여 Fisher’s alpha index (α)를적용하였을때, Cs에서 5.684로가장높 은지수와 Rs에서 1.594로가장 낮은지수를 나타내었고, Shannon diversity index (H’)의적용시에 Cs에서 1.972로 가장높은지수와 Rs에서 0.684로가장낮은지수를나타내 었다. Simpson's diversity index (1-D)는 Cs에서 0.927로 가장높은지수와 Zm에서 0.371로가장낮은지수를나타 내었다. 그리고종풍부도와종다양성지수에서 Cs가높은 지수(Menhinick's index, Fisher’s alpha index, Shannon diversity index, Margalef's richness)로확인된이유는다른 식물보다내생진균류의분포가가장다양하기때문에높은 지수로확인된것으로생각되며, Simpson's index에서 Zm 이가장높은지수로확인된이유는내생진균류의다양성은 낮으면서특정한속의분포비율이가장높기때문인것으로 판단된다. Simpson's index (D)는다양성지수를나타내지 만우점종(Dominant species)에대한의미를가지고있다. 그 리고 Simpson's diversity index (1-D)는다양성지수를나타 내지만균등성지수(Evenness index)에대한의미를가지고 있다. 그리고종다양성지수의값이높은수치가나올수록 다양성은높고, 낮은수치가나올수록종다양성은낮게분 포하는것을의미하고있다. 본연구에서는 6종류의종풍부 도와종다양성지수로나타내었으며, 지수마다나타내는식 의척도가조금씩다르기때문에 6종류의지수를적용하여
비교분석하였다.
본연구는우리나라최대의해안사구지역으로알려진신 두리해안사구에자생하는해안사구식물과공생관계를이루 고있는내생진균의분포및다양성을확인하여, 우리나라 특수환경지역으로분류되는염환경지역의미생물분포상과 분류체계가명확하지않은미생물의분류체계및염환경에 자생하는균류자원연구에기초자료가될것이라생각한다.
요 약
98주의내생진균은신두리해안사구에자생하고있는해안 사구식물의뿌리로부터분리하였다. 8종의해안사구식물샘플 은모래지치(Argusia sibirica), 갯메꽃(Calystegia soldanella), 갯그령(Elymus mollis), 반디지치(Lithospermum zollingeri), 갯무(Raphanus sativus), 솔장다리(Salsola collina), 왕잔디 (Zoysia macrostachya) 및갯잔디(Zoysia sinica)이며, 신두 리해안사구로부터채집되었다. 그리고분리된내생진균들 은 ITS1, 5.8S와 ITS2를포함하는 ITS-rDNA 영역에의해 분석되었다. 해안사구식물로부터분리된내생진균에대하여 다양한지수를적용하여분석하였다. 해안사구식물로부터분 리된 모든 내생진균은 Capnodiales (3.09%), Eurotiales (70.10%), Glomerellales (1.03%), Helotiales (3.09%), Hypocreales (9.28%), Mortierellales (2.06%), Onygenales (1.03%), Ophiostomatales (1.03%), Pleosporales (1.03%), Polyporales (1.03%), Russulales (1.03%), Saccharomycetales
(2.06%), Xylariales (1.03%)로 13개목과분류체계가명확하 지않은 Incertae sedis (3.09%)으로확인되었다. 본연구에서 는 8종의식물로부터내생진균을분석한결과 Eurotiales 목 과 Hypocreales 목의 Penicillium 속(59.18%)과 Fusarium속 (5.10%)이가장많이분포하는것으로나타났다. 갯메꽃으로 부터분리된내생진균이다른해안사구식물들로부터분리된 내생진균의다양성보다높은것으로확인되었다.
Acknowledgments
This work was supported by Korea Ministry of Environment as
“The Eco-Innovation Project” and a grant from National Institute of Biological Resources (NIBR) through Ministry of Environment (MOE) of the Republic of Korea (NIBR No. 2013-02-056).
References
1. Carter RWG. 1991. Near future sea level impacts on coastal dunes landscapes. Landscape. Ecol. 6: 29-39.
2. Fisher RA, Corbet AS, Williams CB. 1943. The relation between the number of species and the number of individuals in a random sample of an animal population. J. Anim. Ecol.
12: 42-58.
3. Garia-Mora MR, Gallego-Fernandez JB, Garcia-Novo F.
2000. Plant diversity as a suitable tool for coastal dune vulner- ability assessment. J. Coastal Res. 16: 990-995.
4. Hasan HA. 2002. Gibberellin and auxin-production by plant root-fungi and their biosynthesis under salinity-calcium inter- Fig. 3. Composition of all endophytic fungi isolated from the Shindu-ri coastal sand dune.
The graph represents the genus (%) which all isolated endophytic fungi belong to.
action. Acta Microbiol. Immunol. Hung. 49: 105-18.
5. Hwang JS, You YH, Bae JJ, Khan SA, Kim JG, Choo YS.
2011. Effects of endophytic fungal secondary metabolites on the growth and physiological response of Carex kobomugi Ohwi. J. Coastal. Res. 27: 544-548.
6. Khan SA, Hamayun M, Kim HY, Yoon HJ, Lee IJ, Kim JG.
2009. Gibberellin production and plant growth promotion by a newly isolated strain of Gliomastix murorum. World J. Micro- biol. Biotechnol. 25: 829-833.
7. Khan SA, Hamayun M, Kim HY, Yoon HJ, Seo JC, Choo YS, et al. 2009. A new strain of Arthrinium phaeospermum iso- lated from Carex kobomugi Ohwi is capable of gibberellin pro- duction. Biotechnol. Lett. 31: 283-287.
8. Khan SA, Hamayun M, Yoon HJ, Kim HY, Suh SJ, Hwang SK, et al. 2008. Plant growth promotion and Penicillium citrinum.
BMC Microbiol. 8: 231.
9. Kim BS, Oh HM, Kang H, Park S, Chun J. 2004. Remarkable bacterial diversity in the tidal flat sediment as revealed by 16S rDNA analysis. Microb. Biotechnol. 14: 205-211.
10. Kil YJ, Eo JK, Eom AH. 2009. Molecular identification and diveristy of endophytic fungi isolated from Pinus densiflora in Boeun, Korea. Korean J. Mycol. 37: 130-133.
11. Lambshead PJD, Platt HM, Shaw, KM. 1983. Detection of dif- ferences among assemblages of marine benthic species based on anssessment of dominance and diversity. J. Nat.
Hist. 17: 859-874.
12. Lim DO, Chekar EK, Choi HW, Hwang IC. 2010. The specific plant species and naturalized plants in the area of Taean- haean National Park, Korea. Korean J. Env. Eco. 24: 117-129.
13. Margalef R. 1958. Information theory in ecology. Gen. Syst. 3:
36-71.
14. Opelt K, Berg G. 2004. Diversity and antagonistic potential of bacteria associated with bryophytes from nutrient poor habi- tats of baltic sea coast. Appl. Microbiol. 70: 6569-6579.
15. Pielou EC. 1975. Ecological diversity. p 165. John Wiley, New York.
16. Simpson EH. 1949. Measurement of diversity. Nature 163:
688.
17. Tao G, Liu ZY, Hyde KD, Liu XZ, Yu XN. 2008. Whole rDNA analysis reveals novel and endophytic fungi in Bletilla ochra- cea (Orchidaceae). Fungal. Divers. 33: 101-122.
18. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar, S. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maxi- mum parsimony methods. Mol. Biol. Evol. 28: 2731-2739.
19. Vazquez MM, Cesar S, Azcon R, Barea JM. 2000. Interaction between arbuscular mycorrhizal fungi and other microbial inoculants (Azospirillum, Pseudomonas, Trichoderma) and their effects on microbial population and enzyme activities in the rhizosphere of maize plants. Appl. Soil. Ecol. 15: 261-272.
20. Waller F, Achatz B, Baltruscha TH, Fodor J, Becker K, Fischer M, et al. 2005. The endophytic fungus Piriformospora indica reprograms barley to saltstress tolerance, disease resistance, and higher yield. PNAS 102: 13386-13391.
21. Williams AT. 1998. Integrated management methods monitor- ing environmental changes in coastal dune ecosystem. pp.
642-653, In Baether KG, Barth H, Bohle-Carbonell M, Fragakis C, Lipiatou E, Martin P, Ollier G, Weydart M (eds.), Porc. 3rd European Marine Science and Technology Confer- ence, Brusells, European Commission 2.
22. Whittaker RH. 1977. Evolution of species diversity in land communities. Evol. Biol. 10: 1-67.
23. Yamada A, Takeo O, Yosuke D, Masatake O. 2001. Isolation of Tricholoma matsutake and T. bakamatsutake cultures from field-collected ectomycorrhizas. Mycoscience 42: 43-50.
24. You YH, Yoon H, Lee GS, Woo JR, Shin JH, Lee IJ, et al.
2011. Diversity and plant growth/promotion of endophytic fungi isolated from the roots of plants in Dokdo islands.
Korean J. Life Sci. 21: 992-996.
25. You YH, Yoon H, Kang SM, Woo JR, Choo YS, Lee IJ, et al.
2012. Cadophora malorum Cs-8-1 as a new fungal strain pro- ducing gibberellins isolated from Calystegia soldanella. J.
Basic Microbiol. 52: 1-5.