Studies of cold resistant glycine betaine effect on cold sensitive Bacillus subtilis mutant strains
8
0
0
전체 글
(2) Cold resistant glycine betaine effect ∙. 201. Bacillus subtilis는 glycine betaine을 생산하지 못하고 외부. 결손은 저온 민감성을 보이며 RNA 분자의 misfolding을 억제. 로부터 흡수하여 사용하거나 전구체인 choline으로부터 합성. 하여 저온 충격에 대응하는 것으로 알려져 있다(Oh and Lee,. 하여 사용한다(Bremer, 2002). Glycine betaine은 3개의 삼투. 2010). 이들 DEAD-box RNA helicase들 중 YdbR과 YqfR은. 압 유도성 수송 시스템인 Opu 수송체가 있는데, 이들은 OpuA,. 저온에서 유도되는 단백질로 이들 두 유전자의 발현은 생장에. OpuC, OpuD로 glycine betaine를 수송한다(Bremer and Krämer,. 필수적으로 작용한다고 보고된 바 있다(Hunger et al., 2006).. 2000; Bremer, 2002; Ziegler, 2010). 이와 같이 Opu 수송체에. 따라서 저온 민감성 돌연변이로 세포막의 지질 성분과 관련. 의하여 수송된 glycine betaine은 삼투보호제로 작용하여 다양. 이 있는 유전자로 bkdR과 des와 RNA 구조 형성과 관련이 있. 한 염 농도 조건에서도 생장이 지속된다. 적절한 삼투보호제. 는 RNA helicase 유전자인 ydbR과 yqfR을 대상으로 glycine. 가 없을 경우 삼투조절이 가능한 proline를 합성한다. Proline. betaine의 효과를 조사하였다.. 생합성에 관여하는 유전자들로는 proJ, proA와 proH들이 연 속적으로 작용한다(Whatmore et al., 1990; Brill et al., 2011a,. 재료 및 방법. 2011b). 고농도의 염분이 존재할 때 Bacillus subtilis는 삼투보 호제를 흡수하거나 아니면 생합성하여 세포 내의 삼투압을 유 지시킨다. 더불어 Bacillus subtilis는 전구물질인 choline으로. 박테리아 배양액 및 생장조건. 부터 GbsB와 GbsA 효소들에 의해 glycine betaine을 합성하. Bacillus subtilis는 JH642 (W168 trpC2 PheA1)를 사용하. 여 삼투압을 조절한다(Boch et al., 1996; Nau-Wagner et al.,. 였으며 SMM (Spizizen’s Minimal Media) 배지에서 배양하여. 2012) Choline은 OpuB와 OpuC ABC 수송 시스템에 의하여. 생장곡선 실험에 사용하였다. SMM은 1.4% K2HPO4, 0.6%. 세포 내로 이동된다(Kappes et al., 1999).. KH2PO4, 0.2% (NH4)2SO4, 0.1% sodium citrate dehydrate,. Glycine betaine과 같은 물질은 세포 내에 축적되어 단백질. 0.02% MgSO4 ․ 7H2O, 0.5% glucose, 50 µg/ml tryptophan, 50. 을 안정화시키는 역할을 한다. 이와 같이 단백질을 안정화시. µg/ml phenylalanine으로 조성하였다. 생장 조건은 37°C 및. 키는 성질은 고온이나 저온에서도 세포들을 유지하고 생장시. 20°C에서 200 rpm으로 진탕 배양하였다. 여러 Bacillus subtilis. 키는 것과 관련되는 것으로 알려져 있다(Holtmann and Bremer,. 균주 선별을 위해 사용한 항생제는 spectinomycin (100 µg/ml),. 2004, 2011). 이와 같은 glycine betaine의 저온에서의 역할에. kanamycin (10 µg/ml), erythromycin (1 µg/ml)과 lincomycin. 대하여 알아보기 위하여 야생형 Bacillus subtilis와 저온에 민. (25 µg/ml) (macrolide-lincosamide-streptogram B 저항성을. 감한 생장을(37°C에서 15~20°C로 온도 변화에 민감성을 보. 테스트)이다.. 이는 생장) 보이는 여러 돌연변이 균주들을 대상으로 저온 조 건에서 glycine betaine과 그 전구체인 choline의 효과에 대해 서도 알아보고자 하였다.. Bacillus subtilis 유전체 정제 및 형질전환 Bacillus subtilis JH642를 5 ml LB 배지에 접종하여 37°C에. 저온 민감성을 보이는 유전자로 세포막의 지질 성분과 관. 서 200 rpm으로 12시간 동안 진탕 배양한 후 1 ml 정도 튜브에. 련이 있는 유전자로 bkdR과 des 등이 알려져 있다. 유전자. 담고 8,000 rpm에서 2분간 원심분리하여 상층 액을 버리고 세. bkdR은 bkd 오페론을 조절하고 저온에서 발현된다(Debarbouille. 포 침전물을 얻는다. 이것을 500 µl TE buffer (10 mM Tris pH. et al., 1999). 발현된 유전자들은 branched-chain fatty acid의. 8.0 / 1 mM EDTA)를 넣고 침전물을 천천히 풀어준다. 풀어 준. 전구물질에 해당하는 keto acid를 제공하여 branched-chain. 뒤 70 µl lysozyme (10 mg/ml)과 2 µl RNase (10 mg/ml) 넣어. fatty acid의 새로운 합성을 촉진한다(Beckering et al., 2002;. 주고 37°C에서 30분간 반응한다. 반응 후에 20 µl proteinase K. Kaan et al., 2002). 합성된 branched-chain fatty acid는 세포막. (10 mg/ml)를 넣고 다시 37°C로 15분간 반응한다. 20 µl 10%. 의 유동성을 증진 시킨다. 반면에 des는 지방산 불포화효소에. SDS와 phenol 500 µl을 넣고 1분간 흔들어 주고 10,000 rpm. 해당하는 유전자로 지방산을 불포화지방산으로 전환시켜 막. 에서 2분간 원심분리를 한다. 위와 같은 방법으로 phenol/. 유동성을 높인다(Altabe et al., 2003; Cybulski et al., 2002). 따. chloroform으로 추출하고 차례로 chloroform으로 추출한 후. 라서 bkdR 혹은 des의 결손이 일어나면 저온에서 세포막의 유. 상층 액을 모은다. 이 상층 액에 에탄올을 넣고 유전체 DNA를. 동성이 떨어지고 저온 민감성 생장을 보이게 된다.. 침전시키고 70% 에탄올로 씻어준 후에 침전물을 말리고 증류. Bacillus subtilis의 putative DEAD RNA helicase로서는. 수를 50 µl에 녹여준다. Bacillus subtilis에 형질전환하기 위해. deaD, ydbR, yfmL, yqfR의 4가지 유전자들이 있으며, 이들의. LB broth 5 ml에 JH642를 배양해서 12시간 정도 키운 후에 Korean Journal of Microbiology, Vol. 54, No. 3.
(3) 202. ∙ Kim and Lee. MC (Modified Competence) media 5 ml에 1/500 비율로. 성시키고 30초, 55°C에서 30초 동안 annealing, 그리고 72°C 6. JH642를 접종하여 OD₆₀₀ = 0.6~0.7이 될 때까지 37°C에서 진. 분간 extention한다. PCR에서 얻은 DNA를 B. subtilis 형질전. 탕 배양한다. MC media는 100 mM phosphate buffer (pH 7),. 환에 사용하였다. 형질전한 후에 각각의 항생제 저항성 선별. 3 mM trisodium citrate, 3 mM MgSO4, 2% glucose, 0.2%. 표지를 이용하여 1차 선별하였다. 결손 돌연변이체의 확인은. potassium glutamate, 0.1% casein hydolysate, 22 µg/ml ferric. 추가적으로 PCR 확인 및 DNA 서열을 조사하여 최종적으로. ammonium citrate, 50 µg/ml tryptophan으로 조성하여 사용하. 유전자 결손을 확인하였다(Oh and Lee, 2010; Kim and Lee,. 였다. OD값을 확인 한 후, double-joint PCR product를 8 µl 정. 2018).. 도 넣고 1시간 정도 37°C에서 진탕 배양한다. 그 후 각각의 LB 항생제 plate에 300 µl, 150 µl를 spreading 하여 37°C에서 배양 한다.. LB 배양액에서의 저온 조건에서의 세포 생장 측정 하루 전에 LB 배양액으로 야생형 균주 JH642와 bkdR, des, ydbR, yqfR 유전자들의 결손 균주들을 키운다. 250 ml 삼각플. 유전자 결손 돌연변이체의 제조. 라스크에 SMM 배양액을 30 ml 넣은 후 각각의 삼각플라스크. Bacillus subtilis JH642로 유전자 결손 돌연변이체를 제조. 에 같은 양의 세포로 접종하고 20°C 조건에서 200 rpm으로 진. 하는 방법으로 LFH (Long-flanking homology) PCR을 이용. 탕 배양한다. 저온 조건에서의 세포 생장은 OD600값을 측정하. 하였다(Kim and Lee, 2018). JH642 유전체 DNA를 주형으. 여 알아보았다.. 로 PCR하여 각각의 up-stream, down-stream, 항생제 저항성 cassette 부분의 DNA를 합성한다(Table 1). 첫 번째 PCR에서. Disk diffusion assay. 얻은 각 upstream, cassette, down stream의 비율은 1:2:1로 넣. 하루 전에 LB broth에서 배양한 Jh642와 bkdR, des, bkdR/des. 고 프라이머로는 upstream F와 downstream R을 사용하여 두. 유전자들의 결손 균주들을 LB broth에 다시 접종하여, OD600 =. 번째 PCR을 수행한다. 이 때 반응 조건은 94°C로 30초 동안 변. 0.4일 때 clean bench에서 각각의 배양액 100 µl 취하여 0.7%. Table 1. Oligonucleotide sequence used for LFH-PCR Oligo sequence bkdR Up F : 5′- CGCTCCTTGCGAATATGACT -3′ bkdR Up-mlsR : 5′- GAGGGTTGCCAGAGTTAAAGGATCCCCGATACCCCTTTGTATGA -3′ bkdRDn-mlsF : 5′- CGATTATGTCTTTTGCGCAGTCGGCCTTGCAAATGAAGGCATGC -3′ bkdRDnR : 5′- GTCGCTTCCATTTTAGGGTT -3′ mls-F : 5′- GATCCTTTAACTCTGGCAACCCTC -3′ mls-R : 5′- GCCGACTGCGCAAAAGACATAATC G -3′ des Up F : 5′- TGCTGCGCTTCAGACGAT C -3′ des Up-kanaR : 5′- CCTATCACCTCAAATGGTTCGCTGGAGCAAATGCAGCGACTTGC -3′ des Dn-kana F : 5′- CGAGCGCCTACGAGGAATTTGTATCGAAGCCTTCCGCCTGATTCAC -3′ des DnR : 5′- GGCAATTCGCTGACGTTCTT -3′ kana-F : 5′- CAGCGAACCATTTGAGGTGATAGG -3′ kana-R : 5′- CGATACAAATTCCTCGTAGGCGCTCGG -3′ YdbR Up F : 5′- GACAGAAAGCCTTGAAGGCT-3′ YdbR Up-spec R: 5′- CGTTACGTTATTAGCGAGCCAGTCTGCTTCTTCAAATCCCATACG -3′ YdbR Dn-spec F : 5′- CAATAAACCCTTGCCCTCGCTACGGGGAAAAGCAACAACCGTTC -3′ YdbR Dn R : 5′- CAGACATCATCGTCAGTCAC -3′ Sepc F : 5′- GACTGGCTCGCTAATAACGTAACGTGACTGGCAAGAG -3′ Sepc R : 5′- CGTAGCGGCAAGGGTTTATTGTTTTCTAAAATCTG -3′ YqfR Up F : 5′- CCTCATTGGCCTCTGTCT -3′ YqfR Up-kana R : 5′- CCTATCACCTCAAATGGTTCGCTGCAAGGCGATGTACTGCATCT -3′ YqfR Dn-kana F : 5′- CGAGCGCCTACGAGGAATTTGTATCGAAGCCGAAGAAGGTTAAGCC -3′ YqfR Dn R : 5′- AGCACTCCGTCAAAGTCTTC -3′ Kana F: 5′- CAGCGAACCATTTGAGGTGATAGG-3′ Kana R: 5′- CGATACAAATTCCTCGTAGGCGCTCGG-3′. 미생물학회지 제54권 제3호.
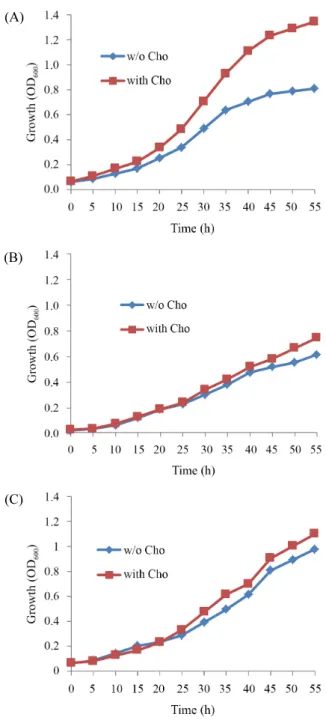
(4) Cold resistant glycine betaine effect ∙. 203. LB soft agar (50°C 유지) 4 ml 들어있는 test tube에 같이 넣고. 존재 유무에 따라 Td값이 780 min과 753 min로 비슷한 생장속. 섞어 준다. 그리고 즉시 LB plate에 (15 ml의 1.5% LB agar) 천. 도를 보였다(Fig. 2B and C). 이와 같이 bkdR과 des 결손 균주. 천히 붓는다. 그 다음 각각의 plate들을 20분 정도 clean bench. 의 경우 저온에서 glycine betaine에 의한 생장 지속이 일어나. 에서 건조한다. 건조한 후 paper disk를 각각의 plate 중앙에 올. 지 않았음을 확인할 수 있었다.. 린 뒤 paper disk에 25% Triton X-100과 N-lauryl sarcosine 10 µl을 넣고 20°C incubator에 넣고 하룻밤 동안 배양한다. (A). 결 과 야생형 및 bkdR와 des 결손 균주들의 저온 생장에 미치는 glycine betaine 효과 저온에서 glycine betaine의 Bacillus subtilis 생장에 미치는 영향을 알아보기 위하여 세포막의 지질 성분과 관련이 있는 저온 민감성 유전자 bkdR과 des가 결손된 균주와 야생형 균주 JH642를 37°C와 20°C에서 진탕 배양하여 생장을 측정하였 다. 37°C에서의 생장은 야생형 및 bkdR과 des가 결손된 균주 모두 거의 비슷한 생장을 보였으며(Td = 100~130 min) glycine. (B). betaine을 첨가한 배지에서도 생장 속도가 비슷하였다(Fig. 1). 반면에 저온 조건인 20°C에서 1 mM glycine betaine을 첨가한 것과 첨가 하지 않은 경우의 생장을 비교하였다. 이전에 보고 된 결과와 같이(Hoffmann and Bremer, 2011) 야생형 균주의 경우 저온에서 glycine betaine이 첨가된 경우 (Td = 420 min) 첨가되지 않은 것에(Td = 610 min) 비하여 생장이 촉진됨을 알 수 있었다(Fig. 2A). 반면에 유전자 bkdR 결손 균주는 glycine betaine이 있을 때 Td값이 699 min로 없을 때의 728 min와 유사 한 생장을 보인다. 또한 유전자 des 결손 균주는 glycine betaine (C). Fig. 1. Growth of B. subtilis JH642, bkdR and des deletion mutants. The cells of wild type and mutants strains were cultivated at 37°C in SMM in the absence and presence of 1 mM glycine betaine. Each growth curve represents the average of three independent experiments.. Fig. 2. Protection of B. subtilis JH642 (A) bkdR (B) and des (C) deletion mutants against cold stress by glycine betaine. The cells of wild type and mutants strains were inoculated (OD600 of about 0.1) from a preculture grown at 37°C in SMM and were then cultivated in a shaking water bath set at 200 rpm at 20°C in the absence and presence of 1 mM glycine betaine. Each growth curve represents the average of three independent experiments.. Korean Journal of Microbiology, Vol. 54, No. 3.
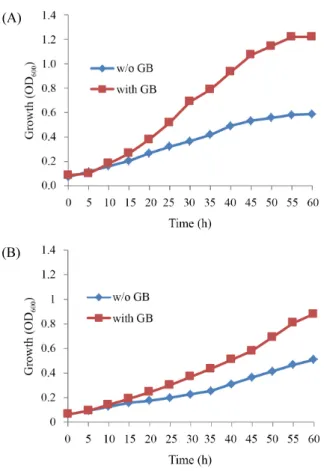
(5) 204. ∙ Kim and Lee. 야생형 및 bkdR와 des 결손 균주들의 저온 생장에 미치는 choline 효과. 이용된다. 따라서 choline을 배양액에 첨가하여 glycine betaine. Choline은 glycine betaine의 전구물질로 작용하며 Bacillus. 이 Bacillus subtilis 생장에 미치는 영향을 알아보기 위하여 저. subtilis 세포 내로 흡수되어 glycine betaine을 생합성 하는데. 과 같은 효과가 일어나는지 알아보고자 하였다. 저온에서 choline 온 민감성 유전자 bkdR과 des가 결손된 균주와 야생형 균주 JH642를 20°C에서 SMM broth로 진탕 배양하여 생장을 측정 하였다. 저온 조건인 20°C에서 20 mM choline을 첨가한 것과. (A). 첨가 하지 않은 경우의 생장을 비교 하였다(Fig. 3). 야생형 균 주의 경우 저온에서 choline이 첨가된 경우 첨가되지 않은 것 에 비하여 생장이 촉진됨을 알 수 있었다. 즉 choline이 없을 때 의 Td는 636 min인 반면에 choline이 있을 때는 Td가 552 min 로 다소 생장이 촉진되었다(Fig. 3A). 반면에 bkdR과 des 결손 균주의 경우 choline의 첨가 유무에 관계없이 Td값이 720 min 와 763 min 그리고 760 min와 725 min로 약간의 차이가 있지 만 비슷한 생장을 보여 야생형 균주와 달리 저온에서 생장 촉 진이 일어나지 않았다(Fig. 3B and C).. (B). ydbR과 yqfR 결손 균주들의 저온 생장에 미치는 glycine betaine 효과 Bacillus subtilis의 DEAD box RNA helicase 유전자를 결손 시켜 저온 민감성을 확인한 결과 ydbR, yfmL, yqfR 순으로 저온 민감성을 보인다(Oh and Lee, 2010). 이들 유전자들의 결손은 RNA의 생체 내 구조를 형성하는데 기여하며 특히 저온에서 는 이들의 역할이 리보솜이 정상적으로 작용하는 구조 형성에 중요한 역할을 한다. 이들 유전자들의 결손이 저온 조건에서 생장에 미치는 영향에 대한 glycine betaine의 효과를 알아보 기 위해 저온 민감성 유전자 ydbR, yqfR 결손 균주에 glycine betaine을 첨가하여 저온 생장을 확인하였다(Fig. 4). Glycine. (C). betaine이 없을 때는 ydbR과 yqfR 결손 균주들의 Td값이 1290 min와 1584 min로 매우 느리게 생장한다. 반면에 glycine betaine 이 첨가된 경우 ydbR과 yqfR 결손 균주들의 Td값이 700 min와 898 min으로 저온에서의 생장이 지속됨을 알 수 있었다. Glycine betaine은 저온에서 단백질 등의 생체물질 안정성에 기여한다 고 알려져 있다(Holtmann and Bremer, 2004, 2011). 이와 같이 생체물질인 RNA 혹은 단백질 안정성이 glycine betaine이 의 해 유도되어 ydbR, yqfR 결손 균주의 저온 지속 생장이 가능한 것으로 생각된다.. bkdR과 des 결손 균주들의 세포막 유동성과 안정성 Fig. 3. Protection of B. subtilis JH642 (A) bkdR (B) and des (C) deletion mutants against cold stress by choline. The cells of wild type and mutants strains were inoculated (OD600 of about 0.1) from a preculture grown at 37°C in SMM and were then cultivated in a shaking water bath set at 200 rpm at 20°C in the absence and presence of 1 mM choline. Each growth curve represents the average of three independent experiments.. 미생물학회지 제54권 제3호. 세포막의 유동성은 세포막 구조에 따라 변화한다. 저온에 서 세포막 유동성은 낮아지며 세포막 구조를 형성하는 분자 구조를 바꾸어 이에 대처한다. 이와 관련된 저온 민감성 유전 자 bkdR과 des는 세포막 유동성을 일으키는 branched-chain.
(6) Cold resistant glycine betaine effect ∙. (A). 205. (A). (B) (B). Fig. 4. Protection of B. subtilis ydbR (A) yqfR (B) deletion mutants against cold stress by glycine betaine. The cells of wild type and mutants strains were inoculated (OD600 of about 0.1) from a preculture grown at 37°C in SMM and were then cultivated in a shaking water bath set at 200 rpm at 20°C in the absence and presence of 1 mM glycine betaine. Each growth curve represents the average of three independent experiments.. Fig. 5. Disk diffusion assay for detergent susceptibility. B. subtilis JH642, bkdR, des and bkdR/des deletion mutants were tested with Triton X-100 (A) and N-lauryl sarcosine (B).. fatty acid (Beckering et al., 2002; Kaan et al., 2002)와 불포화. des 이중 결손의 경우는 1.15 cm로 bkdR 결손 균주와 거의 같. 지방산 합성하여 저온에서 생장이 가능하게 한다(Altabe et al., 2003; Cybulski et al., 2002). 세포막의 안정성과 관련하여 세제에 의한 세포막 용해에 대한 연구 보고에서 세포막의 성 분에 따라 용해 정도가 다르게 나타난다. 특히 이중결합이 있 는 지방산을 포함하거나 콜레스테롤이 있을 경우 세포막 안정 도가 증가하여 용해가 느리게 진행된다(Ahyayauch et al., 2006) 따라서 이들 bkdR과 des 결손 균주의 세포막 성분은 이중결합 이나 가지 달린 지방산이 없어서 세제에 대한 세포막의 안정. 은 크기의 clean zone이 형성 되었다. 세제를 N-lauryl sarcosine 으로 바꾸어 실험한 경우에는 야생형 균주 JH642 0.6 cm, des 결손 균주 0.7 cm, bkdR 결손 균주 0.8 cm, bkdR과 des 이중 결 손 균주의 경우 0.75 cm로 Triton X-100의 결과와 비슷한 결과 를 보였다. 이와 같은 실험 결과 유전자 bkdR의 결손 균주는 세 제에 불안정한 세포막 구조를 형성하는 것으로 보인다. 반면 에 야생형 균주 JH642와 des 결손 균주들은 세제에 민감하지 않았다.. 성이 변화될 것이라 예상된다. 따라서 이와 같은 세제의 세포 막에 대한 효과를 알아보기 위하여 LB plate의 disk에 Triton X-100, N-lauryl sarcosine (25%) 10 µl을 넣어 plate 주변의 세 포 생장에 방해되는 정도를 clean zone의 크기를 비교하였다 (Fig. 5). 이 경우 야생형 균주인 JH642의 zone 크기 1 cm에 비 해 bkdR 결손 균주의 경우 1.25 cm로 더 커졌으나 des 결손 균 주의 경우 0.95 cm로 야생형 균주보다 약간 작아졌다. bkdR과. 고 찰 지구상의 일반적인 생물들의 서식 환경은 낮은 온도에 노 출 될 수 있어(Rodrigues and Tiedje, 2008) 많은 미생물들은 단 백질 접힘, 생합성, 유전자 발현 등의 과정을 항상 추운 환경에. Korean Journal of Microbiology, Vol. 54, No. 3.
(7) 206. ∙ Kim and Lee. 서도 적응하여 생장할 수 있다(Feller and Gerday, 2003; Strocchi. 에 의한 손상이 더 잘 일어나고 있음을 알 수 있었다. 이는 저온. et al., 2006; Shivaji and Prakash, 2010) Bacillus subtilis는 추. 에서 bkdR과 des 결손 균주 세포막의 유동성이 감소되거나 불. 운 환경에서도 생장할 수 있는 전형적인 호저온성 미생물에. 안정하여 세제의 영향이 더 잘 받는 것으로 생각된다. Glycine. 속하지는 않지만(Budde et al., 2006) 일시적으로 낮은 온도에. betaine은 저온에서 야생형 Bacillus subtilis JH642 생장을 지. 노출되고 이에 적응하기 위하여 적당한 화학 물질을 이용한. 속시키는 반면 bkdR과 des 결손 균주에서는 저온에서의 생장. 다. Bacillus subtilis는 토양에 서식하며 식물 분해 산물을 이용. 이 지속되지 않는다. 이는 저온에서 세포막 유동성 감소로. 한다. 많은 식물들은 삼투압 스트레스에 대응하여 glycine. glycine betaine이 세포막 투과에 영향을 미치는 것으로 보인. betaine을 합성하고 뿌리를 통해 토양에 배출시킨다(Chen and. 다. 따라서 저온 조건 bkdR 결손 균주에서는 glycine betaine의. Murata, 2011). 이러한 토양 환경으로부터 Bacillus subtilis는. 막 통과가 잘 일어나는지 확인 할 필요가 있을 것으로 보인다.. glycine betaine을 얻을 수 있고 이를 이용하여 여러 스트레스 에 대응할 수 있다(Whatmore et al., 1990). 최근 보고에 의하면. 적 요. glycine betaine은 고농도 염분 조건에서의 생장뿐 아니라 고 온 및 저온 조건에서도 작동하여 세포생장이 멈추지 않고 유 지되는 것으로 알려져 있다(Brigulla et al., 2003; Holtmann and Bremer, 2004). 저온에서 glycine betaine의 Bacillus subtilis 생장에 미치는 영향을 알아보기 위해 bkdR, des, ydbR과 yqfR 각각의 유전자 결손 돌연변이 균주와 야생형 균주 JH642의 20°C에서의 생장 을 관찰하였다. 이 결과 야생형 균주 JH642는 glycine betaine 에 의해 저온에서 생장이 지속되는 것을 관찰 하였으나, 반면 에 bkdR과 des 결실 균주에서는 glycine betaine 첨가가 저온 생장을 지속시키지 못하는 것을 확인하였다. 그리고 이를 보 다 확인하기 위해 저온 민감성 유전자인 ydbR과 yqfR 결손 돌 연변이를 통하여 확인해본 결과 glycine betaine을 첨가하니 이들의 생장이 지속되는 것을 확인 할 수 있었다. 이는 결과적 으로 ydbR과 yqfR의 경우 RNA helicase 유전자로 RNA 구조 에 영향을 주어 저온 민감성을 보이는 경우이므로 glycine betaine 에 의한 생체물질 안정화로 저온에서의 생장 지속효과가 가능 할 것으로 생각된다. 다른 비교대상으로 glycine betaine의 전 구물질로 작용하는 choline으로 실험을 진행 해보았다. 이 역 시 실험결과는 glycine betaine과 같은 결과를 보였다. 야생형 균주 JH642에서는 choline에 의해 저온 생장이 지속되나, bkdR 과 des의 결손은 choline 유무에 관계없이 저온에서 생장이 억 제되었다. 이는 choline이 세포 안으로 흡수되고 glycine betaine 으로 바뀌어 저온 생장에 관여하고 있음을 확인 할 수 있었다. 세포막 운송과 관련이 있는 유전자 bkdR과 des 결손 균주에 세포막에 영향을 주는 세제의 효과를 저온에서 조사하였다.. 높은 염분 농도에서 glycine betaine은 Bacillus subtilis 안으 로 유입되어 세포 생장이 지속될 수 있게 한다. 뿐만 아니라 최 근 연구 결과에 따르면 저온에서도 glycine betaine이 세포 생 장을 지속시키는 것으로 알려져 있다. 저온에서 Bacillus subtilis 의 생장을 저해시키는 세포 대사 활동으로는 세포막 운송과 단백질 합성을 들 수 있다. 세포막 구조와 관련하여 저온에서 세포막 운송에 영향을 주는 유전자들로는 bkdR과 des가 있고, 단백질 합성 과정에서 RNA helicase 유전자인 ydbR과 yqfR들 은 저온 민감성을 보인다. 따라서 Bacillus subtilis 저온 민감성 유전자 결손 세포들에 대한 glycine betaine의 효과를 조사하 여 저온에서의 glycine betaine 생리적 기능에 대해 알아보고 자 하였다. 이 결과 glycine betaine의 존재 유무에 따라 야생형 Bacillus subtilis와 ydbR과 yqfR 결손 균주의 저온생장에 큰 차 이를 보였다(Td차이 190~686 min). 반면에 bkdR이나 des 결손 균주의 경우에는 glycine betaine 존재 유무에 따라 차이를 보 이지 않았다. Glycine betaine의 전구체인 choline으로 대치하 여도 저온에서의 생장은 같은 결과를 보였다. Glycine betaine 의 영향이 세포막 구조와 관련이 있는 유전자 bkdR과 des 결손 균주에 미치는 영향이 적은 것을 알아보기 위해 세포막에 영 향을 주는 세제의 효과를 조사하였다. Triton X-100과 N-lauryl sarcosine 세제에 의해 bkdR 결손 균주가 야생형에 비해 더 영 향 받는 것을 확인하였고 이는 bkdR 결손이 저온에서 막 구조 를 변형하여 glycine betaine의 투과에 영향을 미치는 것으로 보인다.. 세제인 triton X-100과 N-lauryl sarcosine에 의한 세포 사멸이 일어난 zone의 크기는 bkdR, des, 야생형 균주 순으로 관찰되 었으며, bkdR과 des의 이중 돌연변이도 des 결손 단일돌연변 이와 야생형에 비해 zone의 크기가 더 큰 것을 확인 할 수 있었 다. 이로써 bkdR과 des 결손은 야생형에 비하여 세포막이 세제 미생물학회지 제54권 제3호. 감사의 말 이 논문은 2018 학년도 배재대학교 교내학술연구비 지원에 의하여 수행되었습니다. 이에 감사함을 표합니다..
(8) Cold resistant glycine betaine effect ∙. References Ahyayauch H, Larijani B, Alonso A, and Goni FM. 2006. Detergent solubilization of phosphatidylcholine bilayers in the fluid state: influence of the acyl chain structure. Biochim. Biophys. Acta 1758, 190–196. Altabe SG, Aguilar P, Caballero GM, and de Mendoza D. 2003. The Bacillus subtilis acyl lipid desaturase is a Δ5 desaturase. J. Bacteriol. 185, 3228–3231. Beckering CL, Steil L, Weber MHW, Völker U, and Marahiel MA. 2002. Genomewide transcriptional analysis of the cold shock response in Bacillus subtilis. J. Bacteriol. 184, 6395–6402. Boch J, Kempf B, and Bremer E. 1994. Osmoregulation in Bacillus subtilis: synthesis of the osmoprotectant glycine betaine from exogenously provided choline. J. Bacteriol. 176, 5364–5371. Boch J, Kempf B, Schmid R, and Bremer E. 1996. Synthesis of the osmoprotectant glycine betaine in Bacillus subtilis: characterization of the gbsAB genes. J. Bacteriol. 178, 5121–5129. Bremer E. 2002. Adaptation to changing osmolarity, pp. 385–391. In Sonenshein AL, Hoch JA, and Losick R. (eds.), Bacillus subtilis and its closest relatives. ASM Press, Washington, DC, USA. Bremer E and Krämer R. 2000. Coping with osmotic challenges: osmoregulation through accumulation and release of compatible solutes in bacteria, pp. 79–97. In Storz G and Hengge-Aronis R. (eds.), Bacterial stress responses. ASM Press, Washington, DC, USA. Brigulla M, Hoffmann T, Krisp A, Völker A, Bremer E, and Völker U. 2003. Chill induction of the SigB-dependent general stress response in Bacillus subtilis and its contribution to low-temperature adaptation. J. Bacteriol. 185, 4305–4314. Brill J, Hoffmann T, Bleisteiner M, and Bremer E. 2011a. Osmotically controlled synthesis of the compatible solute proline is critical for cellular defense of Bacillus subtilis against high osmolarity. J. Bacteriol. 193, 5335–5346. Brill J, Hoffmann T, Putzer H, and Bremer E. 2011b. T-box-mediated control of the anabolic proline biosynthetic genes of Bacillus subtilis. Microbiology 157, 977–987. Budde I, Steil L, Scharf C, Volker U, and Bremer E. 2006. Adaptation of Bacillus subtilis to growth at low temperature: a combined transcriptomic and proteomic appraisal. Microbiology 152, 831– 853. Chen TH and Murata N. 2011. Glycinebetaine protects plants against abiotic stress: mechanisms and biotechnological applications. Plant Cell Environ. 34, 1–20. Cybulski LE, Albanesi D, Mansilla MC, Altabe S, Aguilar PS, and de Mendoza D. 2002. Mechanism of membrane fluidity optimization: isothermal control of the Bacillus subtilis acyl-lipid desaturase. Mol. Microbiol. 45, 379–388. Debarbouille M, Gardan R, Arnaud M, and Rapoport G. 1999. Role of BkdR, a transcriptional activator of the sigL-dependent isoleucine and valine degradation pathway in Bacillus subtilis. J. Bacteriol. 181, 2059–2066.. 207. Feller G and Gerday C. 2003. Psychrophilic enzymes: hot topics in cold adaptation. Nat. Rev. Microbiol. 1, 200–208. Hoffmann T and Bremer E. 2011. Protection of Bacillus subtilis against cold stress via compatible-solute acquisition. J. Bacteriol. 193, 1552–1562. Holtmann G, Bakker EP, Uozumi N, and Bremer E. 2003. KtrAB and + KtrCD: two K uptake systems in Bacillus subtilis and their role in adaptation to hypertonicity. J. Bacteriol. 185, 1289–1298. Holtmann G and Bremer E. 2004. Thermoprotection of Bacillus subtilis by exogenously provided glycine betaine and structurally related compatible solutes: involvement of Opu transporters. J. Bacteriol. 186, 1683–1693. Hunger K, Beckering CL, Wiegeshoff F, Graumann PL, and Marahiel MA. 2006. Cold-induced putative DEAD-box RNA helicase CshA and CshB are essential for cold adaptation and interact with cold shock protein B in Bacillus sutilis. J. Bacteriol. 188, 240–248. Kaan T, Homuth G, Mader U, Bandow J, and Schweder T. 2002. Genome-wide transcriptional profiling of the Bacillus subtilis cold-shock response. Microbiology 148, 3441–3455. Kappes RM, Kempf B, Kneip S, Boch J, Gade J, Meier-Wagner J, and Bremer E. 1999. Two evolutionarily closely related ABC transporters mediate the uptake of choline for synthesis of the osmoprotectant glycine betaine in Bacillus subtilis. Mol. Microbiol. 32, 203–216. Kempf B and Bremer E. 1998. Uptake and synthesis of compatible solutes as microbial stress responses to high osmolality environments. Arch. Microbiol. 170, 319–330. Kim DH and Lee SS. 2018. Cold shock sensitive growth of Bacillus subtilis mutants deleted for genes involved in fatty acid synthesis. Korean J. Microbiol. 54, 9–17. Nau-Wagner G, Opper D, Rolbetzki A, Boch J, Kempf B, Hoffmann T, and Bremer E. 2012. Genetic control of osmoadaptive glycine betaine synthesis in Bacillus subtilis through the choline-sensing and glycine betaineresponsive GbsR repressor. J. Bacteriol. 194, 2703–2714. Oh EH and Lee SS. 2010. Cold sensitive growth of deletion mutants of DEAD-box RNA helicase genes in Bacillus subtilis. Korean J. Microbiol. 46, 233–239. Rodrigues DF and Tiedje JM. 2008. Coping with our cold planet. Appl. Environ. Microbiol. 74, 1677–1686. Shivaji S and Prakash JS. 2010. How do bacteria sense and respond to low temperature? Arch. Microbiol. 192, 85–95. Strocchi M, Ferrer M, Timmis KN, and Golyshin PN. 2006. Low temperature-induced systems failure in Escherichia coli: insights from rescue by cold-adapted chaperones. Proteomics 6, 193– 206. Whatmore AM, Chudek JA, and Reed RH. 1990. The effects of osmotic up shock on the intracellular solute pools of Bacillus subtilis. J. Gen. Microbiol. 136, 2527–2535. Ziegler C, Bremer E, and Krämer R. 2010. The BCCT family of carriers: from physiology to crystal structure. Mol. Microbiol. 78, 13–34.. Korean Journal of Microbiology, Vol. 54, No. 3.
(9)
수치


관련 문서
Key words: Regulatory Impact Analysis (RIA), Cost-Benefit Analysis, Benefit Evaluation, Benefit Measurement, Benefit Overestimation.. 또한 이명박
The proposal of the cell theory as the birth of contemporary cell biology Microscopic studies of plant tissues by Schleiden and of animal tissues by Microscopic studies of
일본 Tokyo University of Marine Science and Technology 자매결연 2018. 태국 Chiang
[oragrid:+ASM1:/home/oragrid> opatch lsinventory Oracle Interim Patch Installer version 11.2.0.3.18.. Copyright (c)
It considers the energy use of the different components that are involved in the distribution and viewing of video content: data centres and content delivery networks
After first field tests, we expect electric passenger drones or eVTOL aircraft (short for electric vertical take-off and landing) to start providing commercial mobility
European journal of clinical microbiology & infectious diseases : official publication of the European Society of Clinical Microbiology.. Luchi M, Morrison DC, Opal S,
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
