저산소증에 의한 활막 섬유모세포에서 I CAM-1 발현의 증가
전북대학교 의과대학 내과학교실*, 정형외과학교실**, 임상연구소†
전현순*・백홍선*,†・황병연* *,†・류완희*,†
─A b s t r a c t─
Hypoxia Increases the Expression of ICAM-1 in Cultured Human Synovial Fibroblasts
Hyun Soon Jeon, M.D.*, Hong Sun Baek, M.D.*,†
Byung Yun Hwang, M.D.**,†, Wan Hee Yoo, M.D.*,†
Department of Internal Medicine* and Orthopedic Surgery**, Chonbuk National University Medical School and Research Institute of Clinical Medicine
†, Chonju, Korea
O b j e c t i v e : Hypoxic conditions are thought to be exist in inflamed arthritic synovium. Several in vitro and in vivo studies indicate that hypoxia can initiate events that lead to pro-adhesive changes. Therefore, this study was designed to examine the effects of hypoxia on the expression of ICAM-1 by cultured human synovial fibroblasts.
M e t h o d s : Synovial fibroblasts were isolated from patients with RA and cul- tured at hypoxic condition. To quantify the expression of ICAM-1 mRNA in synovial fibroblasts, RT-PCR was performed. The levels of cytokines in culture supernatants were measured by ELISA. The activation of NF-κB was analyzed by electrophoretic mobility shift assay. The assay of adhesive reaction between synovial fibroblasts and lymphocytes was performed by photographic evaluation with colloidin blue staining and measurement of fluorescent intensity of BCECF- AM in lymphocytes.
<접수일 : 2001년 8월 13일, 심사통과일 : 2001년 11월 1일>
※통신저자 : 류이완이희
전북 전주시 덕진구 금암동 634-18 전북대학교 의과대학 내과학교실
Tel : 063) 250-1672, Fax : 063) 254-1609, E-mail : [email protected] 이 논문은 전북대학교병원 임상연구소의 학술연구비 지원에 의하여 연구되었음.
Vol. 9, No. 1, March, 2002
서 론
류마티스 관절염은 관절의 파괴를 특징으로 하는 전신 질환이며, 활막 섬유모세포(synovial fibrob- l a s t )의 증식과 함께 염증세포의 침윤에 의해 활막의 과형성( h y p e r p l a s i a )이 일어나게 된다1 , 2 ). 염증세포 가 침윤되는 과정에는 여러 가지 c y t o k i n e s과 chemokines 및 유착분자(adhesion molecules)가 중요한 역할을 하는 것으로 보고되었다3 - 1 0 ). Intercel- lular adhesion molecule 1 (ICAM-1)과 같은 유착 분자들은 활막 섬유모세포와 염증세포간의 상호작용 을 매개함으로써 염증반응을 지속시키고 관절의 파괴 를 일으키는데 관련이 있는 것으로 알려져 있다7 - 1 0 ).
류마티스 관절염에서 염증성 활막은 활막 조직의 부종에 의해 모세혈관이 눌리게 되고, 표재성 모세 혈관이 부족하여 현저한 저산소증 상태에 있는 것으 로 보고되어 있다1 1 , 1 2 ). 또한 운동 유발성 저산소증- 재관류 과정(hypoxia-reperfusion cycle)에 의해 생성되는 반응성 산화물(reactive oxygen species) 에 의한 조직 손상도 이에 관여하는 것으로 알려져
있다1 3 ). 염증성 관절에서 반응성 산화물에 의한 저산
소증-재관류 손상이 활막의 염증을 지속시킨다는 실 험 결과들은1 4 , 1 5 )
, 저산소증이 류마티스 관절염에서도 중요한 역할을 할 것으로 추측할 수 있다. 이후 여러 연구들에서 저산소증은 내피세포 등에서 c y t o k i n e s 과 유착분자를 발현시킴으로써 염증 반응을 지속시 키고, 이러한 과정에 N F -κB와 같은 전사인자가 관 여하고 있음을 보고하였다1 6 - 1 8 ).
이에 본 저자들은 류마티스 관절염 환자의 활막 섬유모세포를 저산소증 상태에서 배양하면서 활막 섬유모세포에서 I C A M - 1과 같은 유착분자의 발현과 활막 섬유모세포와 염증세포 간의 유착반응에 저산 소증이 미치는 영향에 대해 알아보고자 하였다. 또 한 이러한 과정에 관련되리라 여겨지는 c y t o k i n e s (IL-1, TNF-α) 및 전사인자( N Fκ- B )의 발현을 조 사함으로써 저산소증에 의한 I C A M - 1의 발현이 일 어나는 기전에 대해서도 알아보고자 하였다.
대상 및 방법
1. 활막 섬유모세포의 분리와 배양
미국 류마티스 학회(American College of R h e u m a t o l o g y )의 진단기준1 9 )을 만족하는 류마티스 관절염 환자에서 무릎 관절의 활막 제거술이나 관절 치환술을 통해 얻은 조직으로부터 Matsuoka 등2 0 )이 기술한 방법을 적용하여 활막 섬유모세포를 분리하였 다. 활막 조직을 가위로 잘게 자른 후 c o l l a g e n a s e (Sigma Chemical Co, St. Louis, Missouri, U S A )로 3 7°C에서 4 5분간 처리한 후 세포 부유액을 나일론 망에 통과시키고 남은 세포를 RPMI 1640- 10% FCS이 담긴 petri dish에 담아 두었다. 비 부 착세포를 P B S로 3회 씻어 제거하고 남아 있는 부착 세포를 계대 배양하였다. 배양세포가 활막 섬유모세 포임을 확인하기 위하여 anti-CD3 단일클론 항체, anti-CD68 단일클론 항체, anti-CD20 단일클론 항 체, anti-vWF 항체 등을 사용한 flow cytometry (Becton-Dickinson, Mountain View, CA, R e s u l t s : The expression ICAM-1 mRNA was increased at 12 to 48 hours of hypoxic stimuli and peaked at 24 hours (p<0.01). The concentration of IL-1 and T N F -αwas increased as the duration of hypoxic stimuli was increased. Hypoxia also induced activatation of NF-κB, which was inhibited by antioxidant (pyrroli- dine dithiocarbamate). Fibroblast-lymphocyte adhesive interaction was increased significantly by hypoxic stimuli (p<0 . 0 1 ) .
C o n c l u s i o n s : Synovial hypoxia may have an important role in inflammatory synovitis, in part, as a cause of adhesion molecule (ICAM-1) expression in syn- ovial fibroblasts and recruitment of inflammatory cells.
Key Words : Hypoxia, ICAM-1, Synovial fibroblasts
U S A )를 이용하여 배양세포 내에 T 림프구, B 림프 구, 대식세포, 내피세포 등이 1% 미만임을 입증하였 다. 활막 섬유모세포는 uridine diphosphoglucose dehydrogenase 염색을 이용하여 확인하였다.
2. 저산소증 상태에서 활막 섬유모세포의 생존도 저산소증 상태의 조건을 만들기 위하여 n i t r o g e n g a s를 주입함으로써 i n c u b a t o r의 조건을 5% CO2,
>95% N2, <3% O2가 되도록 하였다. 37°C에서 배양하였으며, 최대 4 8시간 동안 저산소증에 노출시 키는 동안에는 O2 분석기를 이용하여 i n c u b a t o r내의 O2농도를 지속적으로 감시하였다. 저산소증 상태에서 활막 섬유모세포의 생존도는 trypan blue exclu- s i o n2 1 ) 방법과 세포배양 상층액에서 LDH (lactate dehydrogenase) 활성도를 통해 측정하였다.
3. 림프구의 분리와 배양
림프구는 단핵세포(mononuclear cells)를 P e r- coll gradient centrifugation을 이용하여 t h r e e step method로 분리하였다2 2 ). 항응고제가 포함되어 있는 주사기로 채혈하여 70% Percoll, 55% Per- coll gradient로 5 0 0×g으로 3 0분간 원심하여 단핵 세포 분획을 얻어 비 부착세포를 제거하였고, 적혈 구가 포함되어 있으면 적혈구 용해액을 처리하여 제 거하였다. 이런 과정을 통해 얻은 림프구는 1 0 % FBS-RPMI 1640에서 배양하였다.
4. Flow cytometric analysis of cell surface I C A M - 1
활막 섬유모세포에서 ICAM-1 발현을 관찰하기 위하여 F A C S c a n을 이용하였다. 배양 평판( c u l- ture plate)에 trypsin EDTA를 처리한 후 원심 분리하여 활막 섬유모세포를 수집하였다. 수집된 세 포를 1mg/ml BSA가 함유된 HBSS (Hanks’
balanced salt solution)로 2회 세척하고 세포 수 를 5×1 05 c e l l s / m l로 맞추어 staining buffer (0.01 M NaPO4, 0.15 M NaCl, pH 7.1, con- taining normal human serum, 0.02% NaN3) 에 넣고 세포 부유액에 F I T C가 표지된 h u m a n anti-ICAM-1 항체 1 0μg / m l를 첨가한 후에 얼음 위에서 3 0분간 부치시켰다. 이런 과정 후 H B S S로 2회 세척하고 1% paraformaldehyde 용액(300 μ
l )으로 고정한 후에 flow cytometer로 분석하였다.
5. RNA의 분리와 I C A M - 1의 R T - P C R
배양 평판에 trypsin EDTA를 처리한 후 원심 분 리하여 활막 섬유모세포만을 수집하고, Trizol reagent (Gibco-BRL, Gaithersburg, MD, U S A )를 이용하여 R N A를 추출한 후 분광광도계 ( s p e c t r o p h o t o m e t e r )를 이용하여 m R N A를 정량하 였다. 50 ng의 m R N A로부터 M-MLV RNase H- reverse transcriptase (Gibco-BRL, Gaithers- burg, MD, USA)와 oligo-p(dT)15 primer을 이 용하여 c D N A을 합성한 후 이를 주형으로 하고 ICAM-1 염기서열로부터 유추한 o l i g o n u c l e o t i d e s , HindIII-ICAM-1 (5’ CAC GCC TCC CTG AAC CTA 3’)과 ICAM1-XbaI (3’ AGT CCC AGC CCC ATT TGA 5’)를 p r i m e r로 이용하여 P C R을 실시하였다. 증폭된 PCR 산물을 1% agarose gel을 이용한 전기영동을 통하여 정량분석 비교하였다.
6. Electrophoretic mobility shift assay (EMSA) 활막 섬유모세포 만을 수집한 후에 핵 추출물 (nuclear extract)을 얻어 분광광도계를 이용하여 단백질을 정량하였다. NF-κB의 활성화를 조사하기 위해서 핵 추출물( 1 0μg )과 N F -κB의 c o n s e n s u s DNA binding site를 end label한 P3 2labeled B probe (Ig-사슬결합부위 B, 5’CCG GTT AAC AGA GGG GGC TTT CCG AG3’, 10,000 c p m )를 2 5°C에서 3 0분간 반응시켰다. Probe 결합 의 특이성은 5 0배 농도의 unlabeled probe를 첨가 하여 확인하였고, 단백질-DNA 복합체는 4% non- denaturating polyacrylamide gel에서 전기영동 을 한 후에 N F -κB의 활성도를 측정하였다.
7. Cytokines의 측정
배양 상층액에서 pro-inflammatory cytokines 의 변화를 조사하기 위해서 I L - 1와 T N F -α를 sandwitch ELISA kit (BioSource, Sunnyvale, CA, USA)를 이용하여 측정하였다.
8. 활막 섬유모세포와 림프구의 유착반응 조사 활막 섬유모세포와 림프구간의 유착정도를 조사하 기 위하여 먼저 정상인의 혈액에서 림프구를 분리한
후 phorbol 12-myristate 13-acetate (PMA, Cal- biochem, La Jolla, CA, USA) 5ng/ml로 처리하 여 림프구를 활성화시켰다. 활성화된 림프구를 2 , 7 - bis (carboxyethyl)-5(6)-carboxyfluorescein-ace- toxymethyl ester (BCECF-AM, 5μM final con- centration, Calbiochem, La Jolla, CA, USA)로 3 7°C에서 3 0분간 반응시켜 표지한 후에 H B S S로 3 회 세척하였다. 활막 섬유모세포에 대한 림프구의 유 착정도를 보기 위하여 6-well 배양 평판에 활막 섬유 모세포를 배양한 다음 B C E C F - A M으로 표식된 림프 구( 2×1 06)를 첨가하여 활막 섬유모세포와 림프구가 서로 유착되도록 5% CO2 i n c u b a t o r에서 3 7°C에서 1 0분간 반응시킨 후 유착되지 않은 림프구를 제거하 기 위해 H B S S로 3회 세척하였다. 활막 섬유모세포 와 이에 유착된 림프구를 trypsin EDTA으로 처리한 후 원심 분리하여 수집하여 fluorescent plate read- e r를 이용하여 형광강도(485-nm excitation, 530-
nm emission)를 측정하였다. 활막 섬유모세포에 유 착된 림프구의 수는 일정한 비율로 희석한 림프구를 이용하여 B C E C F - A M를 처리한 후 형광강도를 측정 하여 standard curve를 얻고, 이를 이용하여 유착된 림프구의 비율을 계산하였다.
활막 섬유모세포와 이에 유착된 림프구를 관찰하 기 위하여 배양 평판에 cover glass를 두어 이 곳에 활막 섬유모세포를 배양하였다. 활막 섬유모세포와 림프구가 서로 유착되도록 P M A로 자극한 림프구를 첨가하여 1 0분간 배양한 후에 유착되지 않은 림프구 를 P B S로 3회 세척하여 제거하고, 남은 세포를 10% formaldehyde로 고정하고 colloidin blue로 염색하여 광학현미경으로 관찰하였다.
9. 항산화제
세포내 전사인자인 N F -κB의 억제제로는 항산화 제인 PDTC (pyrrolidine dithiocarbamate)를 사
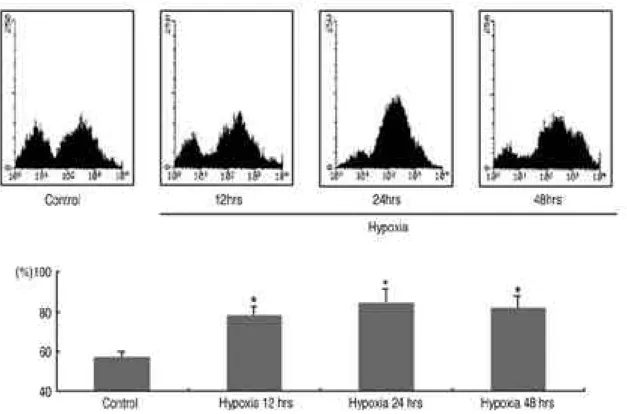
Fig. 1. ICAM-1 expression in synovial fibroblasts cultured in hypoxic condition. Cell surface ICAM-1 expression was analyzed after 12, 24, 48 hours of hypoxic stimuli, and compared with control sample (normoxic culture). Samples were analyzed using FACScan and the results were expressed as percentage of positive cell. *P<0.05 compared with control of normoxic culture condition.
용하였다. PDTC는 IκB의 인산화 ( p h o s p h o r y l a- t i o n )를 억제함으로써 N F -κB로부터 IκB의 분리를 막아 phorbol esters, IL-1과 T N F -α등의 효과를 억제하는 것으로 알려져 있다2 3 ).
10. 통계처리
모든 실험 결과는 평균 standard deviation (SD) 으로 표기하였고, analysis of variance (ANOVA) 를 이용하여 통계처리 하였다. P values<0 . 0 5를 유 의 수준으로 정하였으며, Student’s paired t-test를 이용하여 각 군간의 차이를 비교하였다.
결 과
1. 저산소증이 활막 섬유모세포의 생존도에 미치는 영향
Trypan blue exclusion상 저산소증에 노출된 4 8시간까지 활막 섬유모세포의 생존도는 98% 이상 을 유지하였다. 저산소증 자극 후 각 시간별로 배양 상층액에서 측정한 LDH 활성도는 저산소증에 노출 된 시간 경과에 따라 증가하는 경향을 보였으나 대 조군(normoxic condition)과 유의한 차이를 보이
지 않았다. 그리고, 저산소증 자극을 주기 전에 항 산화제인 P D T C를 첨가한 경우에도 유의한 차이를 보이지 않았다(data not shown).
2. 저산소증이 활막 섬유모세포에서 I C A M - 1의 발 현에 미치는 영향
1) Flow cytometric analysis of cell surface I C A M - 1
활막 섬유모세포에 저산소증에 의한 자극을 주면 I C A M - 1의 발현이 대조군 (normoxic condition) 보다 증가하는 것을 관찰할 수 있었다(그림 1 ) . I C A M - 1을 발현하는 활막 섬유모세포의 비율이 대조 군에서 5 7±2 . 8 %이었던 것에 비해, 저산소증 자극을 준 후 12, 24, 48시간에는 7 8±4.8%, 84±7 . 5 % , 8 1±6 . 8 %로 유의하게 증가하였다 ( p<0 . 0 5 ) .
2) RT-PCR for ICAM-1 mRNA
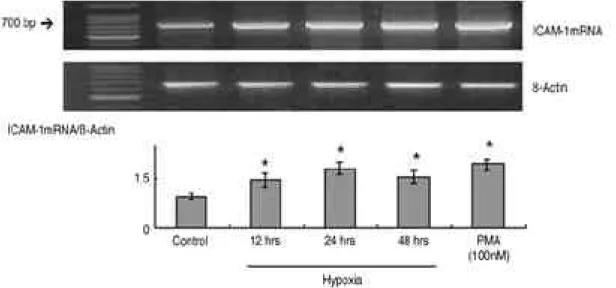
위의 결과를 R T - P C R을 통해 확인해 보면 대조군 에 비해 저산소증 시에 ICAM-1 mRNA의 발현이 증가하는 것을 관찰할 수 있었다( p<0.01). ICAM- 1 mRNA/β-actin ratio 역시 대조군에 비해 증가 하였으며, 24시간에 최대치를 보였다(그림 2 ) .
Fig. 2. ICAM-1 mRNA expression in synovial fibroblasts exposed to hypoxic condition (<3% O2) for 12, 24, 48 hours, as determined by RT-PCR analysis. The ICAM-1 mRNA/β-actin ratio were pre- sented as the amount of the induction of ICAM-1 mRNA transcription, and the results shown above are representative of three separate experiments. *P<0.01 compared with control of nor- moxic culture condition.
3. 배양 상층액에서 I L - 1과 T N F -α농도
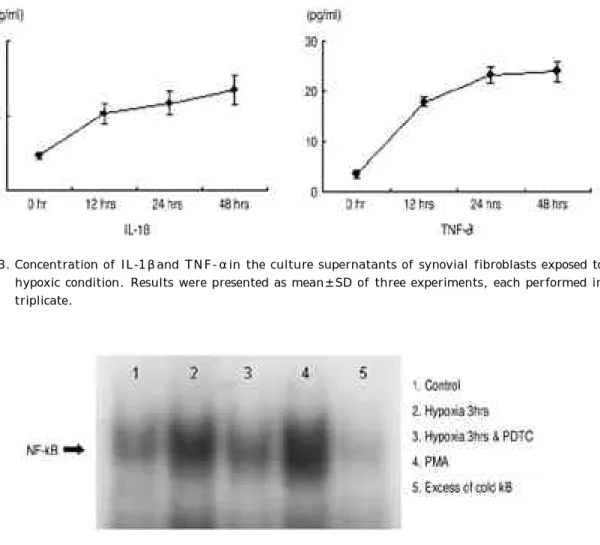
Normoxic culture condition에 비해 저산소증에 의한 자극을 주었을 때, 배양 상층액의 I L - 1과 T N F -α 가 증가하는 소견을 보였고, 저산소증에 노출된 시간이 경과함에 따라 I L - 1과 T N F -α가 증가하였다(그림 3 ) .
4. NF-kB의 활성화
저산소증에 의한 자극에 따른 N F -κB의 활성화 여 부를 알아보기 위해서 저산소증 상태로 3시간 동안 배양한 후에 N F -κB의 활성도를 측정하였다. Nor-
moxic culture condition에 비해 저산소증에 의한 자극을 준 활막 섬유모세포에서 P M A를 처리한 것 처럼 N F -κB의 활성도가 증가됨을 확인할 수 있었으 며, 항 산화제인 PDTC 200 μM / m l를 전처치를 한 경우에는 저산소증에 의한 N F -κB의 활성도가 억제 되었다(그림 4 ) .
5. 활막 섬유모세포와 림프구와의 유착반응 림프구와의 유착반응을 colloidin blue로 염색을 한 후에 광학현미경으로 관찰하여 본 결과, nor- moxic condition에서 배양한 활막 섬유모세포 비하 Fig. 3. Concentration of IL-1βand TNF-αin the culture supernatants of synovial fibroblasts exposed to hypoxic condition. Results were presented as mean±SD of three experiments, each performed in t r i p l i c a t e .
Fig. 4. N F -κB activation in rheumatoid synovial fibroblasts exposed to hypoxic cul- ture condition with/without antioxidant PDTC (200μM) and phorbol 12- myristate 13-acetate (PMA, 100 nM as a positive control) as determined by electrophoretic mobility shift assay using P3 2-labeled oligonucleotides. A 50- fold of excess of cold κB was added as competitors.
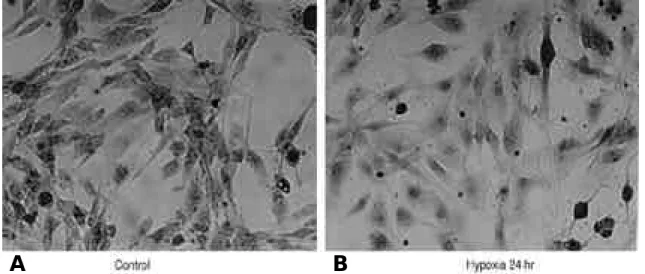
여 저산소증 자극시에 활막 섬유모세포에 보다 많은 수의 림프구가 결합하고 있는 것이 관찰되었다(그림 5). 또한, BCECF-AM을 이용하여 유착반응을 조 사한 결과에서도 저산소증에 의한 자극을 받은 활막 섬유모세포에 보다 많은 수의 림프구가 결합하고 있 음을 관찰할 수 있었다(p<0.01) (그림 6 ) .
고 찰
류마티스 관절염은 조직학적으로 활막세포의 증식 에 의한 활막의 과형성과 염증세포의 광범위한 침윤 을 가장 중요한 특징으로 한다1 , 2 ). 아직까지 활막염 Fig. 5. Lymphocyte adhesion assay. Synovial fibroblasts were cultured at normoxic (A) and hypoxic cul- ture condition for 24 hours (B). PMA-stimulated and BCECF-AM labeled lymphocytes enriched mononuclear cells were co-cultured with synovial fibroblast monolayer in culture plates for 10 minutes. Culture plates were washed three times with PBS for removal of non-adherent cells and fixed with 10% formaldehyde and stained with colloidin blue, ×2 0 0 ) .
A B
Fig. 6. Lymphocyte adhesion assay. The fluorescent intensity of adherent lymphocytes was measured by fluorescent plate reader. Results were % of bounded lymphocytes and presented as mean±SD of 3 experiments, each performed in triplicate. *P<0.01 compared with control of normoxic culture c o n d i t i o n .
의 원인과 염증이 지속적으로 진행하는 기전에 대해 서는 명확히 밝혀져 있지 않지만 염증세포의 침착이 중요하며, 여러 가지 cytokines 및 유착분자들이 중 요한 역할을 하는 것으로 알려져 있다3 - 1 0 ). 염증성 활 막에서 활막 섬유모세포는 I L - 1와 T N F -α와 같은 염증성 c y t o k i n e s과 유착 분자들을 발현함으로써 염 증세포의 침윤 및 활성화와 분화를 도와 염증반응을 유지하고 관절의 파괴에 관여하는 것으로 여겨진다.
여러 생체 외 및 생체 내 실험을 통해서 활막 섬유모 세포에는 ICAM-1, VCAM-1, E-selectin 등의 발 현이 증가되는 것으로 보고되었으며8 - 1 0 , 2 4 - 2 5 )
, 유착분 자의 발현이 없는 knockout mice와 유착분자에 대 한 단일 클론항체는 염증반응에 의한 조직 손상을 막을 수 있다는 보고가 있다1 0 ). 따라서, 이러한 유착 분자들의 발현이 세포간의 상호작용에 영향을 미쳐 류마티스 관절염의 병인에 중요한 역할을 할 수 있 을 것으로 추측을 할 수 있다.
염증성 관절이 어떠한 기전에 의해 저산소증 상태 에 있는지에 대해서는 여러 가지 견해가 있다. 염증 성 활막에서 대사 요구량의 증가에 의한 산소 소비량 의 증가가 저산소증에 기여할 것으로 여겨지며, 혐기 성 당대사 과정의 증가에 의한 젖산( l a c t a t e )의 증가 가 보고된 바 있다2 6 , 2 7 ). 염증성 활막에서 관류가 증가 된다는 이전의 보고가 있기는 하였지만, 동정맥 단락 ( s h u n t )으로 인해 활액 내 산소 분압이 감소됨으로 써 산소 공급이 부적절할 수 있을 것으로 여겨진다2 8 ). 염증성 활막 조직의 부종에 의해 모세혈관이 눌리게 되고, 표재성 모세혈관이 부족함으로 인해서 현저한 저산소증 상태가 초래된다는 보고도 있다1 1 , 1 2 )
. 또한 운동 유발성 저산소증-재관류 과정에 의해 생성되는 반응성 산화물에 의한 조직 손상도 이에 관여할 수 있다는 보고도 있다1 3 ).
정상 활막강(synovial cavity)은 음압 상태에 있 고, 움직일 때에도 혈관의 개통( p a t e n c y )이 유지됨 으로 해서 혈관이 없는 연골에 영양을 공급하게 된 다. 그러나 류마티스 관절염에서 활막강내의 압력은 상승되어 있고, 움직이게 되면 모세 혈관 관류압 이 상으로 상승되어 혈관을 허탈( c o l l a p s e )시킨다. 이러 한 운동 유발성 저산소증-재관류 과정에서 저산소증- 재관류 손상에 의해 반응성 산화물은 생성되게 된다.
이렇게 생성된 반응성 산화물은 I g G를 산화시켜 류 마티스 인자를 생성하게 하고, hyaluronan을 분절
화시키고 지질로부터 알데하이드를 생성하여 면역기 능에 영향을 주며, 지단백( l i p o p r o t e i n )으로부터 monocyte chemotactic peptide를 생성함으로써 염증반응에 중요한 역할을 한다고 알려져 있다2 9 , 3 0 ). 또한 N F -κB 등과 같은 전사인자의 o x i d a t i v e i n d u c t i o n을 통해 여러 가지 c y t o k i n e s과 유착분자 의 발현을 증가시킴으로써 염증반응을 지속시키는 것 으로 보고되고 있다1 6 - 1 8 , 3 1 - 3 7 ).
따라서, 저자들은 활막의 저산소증 상태의 자극에 대한 영향을 조사하기 위하여 활막 섬유모세포를 hypoxic condition에서 배양하면서 I C A M - 1의 발 현을 조사하였다. 또한 유착분자의 발현에 관련되리 라 여겨지는 cytokines (IL-1, TNF-α) 및 N F -κB 의 변화도 조사하였다.
Chin 등3 8 )이 류마티스 관절염 환자의 활막 섬유모 세포를 normoxic condition에서 배양하여 시간에 따 른 I C A M - 1의 발현의 변화를 관찰하였던 것을 고려 하면, 저자들이 시행한 저산소증 상태에서 배양한 활 막 섬유모세포에서는 I C A M - 1의 발현이 증가되고 발 현시간이 연장되는 것을 확인할 수 있었다. ICAM-1 과 같은 유착분자는 단핵구-대식세포, 림파구, 혈관 내피세포 등 여러 세포에서 발현되는 세포 표면의 당 단백( g l y c o p r o t e i n )으로서 세포간의 상호작용에 중요 한 역할을 한다. 이러한 I C A M - 1은 류마티스 관절염 의 활막 조직에서도 발현이 증가되고8 , 1 0 , 2 4 , 2 5 )
, 이에 대 한 차단항체를 실험동물에게 투여한 경우 관절염의 발생이 늦어지며1 0 ), 또한 활막 조직에서 I C A M - 1의 발현 정도가 활막염의 정도와도 관련이 있다는 보고 가 있어 유착분자는 류마티스 관절염의 병인에 관련 되는 것으로 알려져 있다2 4 , 2 5 ). 따라서, 본 연구 결과 는 활막의 저산소증은 류마티스 관절염에서 I C A M - 1 의 발현을 증가시켜 활막염의 발생 및 병인에 중요한 역할을 할 수 있을 것으로 여겨진다.
저산소증에 의해 자극된 활막 섬유모세포에서 I C A M - 1의 발현 증가는 림프구 등과 같은 염증세포와 의 유착반응을 증가시킴으로써 활막에 염증세포의 침 윤을 일으킬 수 있을 것으로 여겨진다. 저자들은 광학 현미경과 B C E C F - A M을 이용한 유착반응을 조사한 결과에서 모두 림프구와 활막 섬유모세포간의 유착반 응이 증가되는 것을 확인할 수 있었다. 이러한 결과는 류마티스 관절염에서 저산소증이 활막 섬유모세포에서 의 유착분자의 발현을 증가시켜 염증세포의 이동과 침
윤을 통해서 만성 염증을 지속시키고 관절 파괴를 일 으킬 수 있음을 시사하는 소견이라 할 수 있다.
본 연구에서는 유착분자의 발현에 중요한 역할을 하 는 것으로 알려진 I L - 1과 T N F -α가 저산소증에 의해 활막 섬유모세포에서 분비되는지 여부를 알아보았으 며, 저산소증에 의한 자극에 의해 활막 섬유모세포의 배양 상층액에서 IL-1, TNF-α이 증가하는 것을 확인 할 수 있었다. 이러한 결과는 저산소증이 활막 섬유모 세포를 자극하여 I L - 1과 T N F -α와 같은 c y t o k i n e s을 분비시키고, 이러한 c y t o k i n e s이 활막 섬유모세포에 서 I C A M - 1의 발현에 영향을 줄 수 있을 것으로 생각 해 볼 수 있었다. 이는 저산소증에 의해 증가된 I L - 1 , T N F -α등의 c y t o k i n e s이 autocrine stimulation을 통하여 유착분자의 발현에 영향을 줄 수 있는 것으로 알려진 이전의 보고들1 6 - 1 8 , 3 1 - 3 7 )
과 일치하는 소견이다.
저산소증 자극에 의해 생성되는 reactive oxygen s p e c i e s는 세포내 전사인자인 N F -κB를 활성화시키 고, 이는 여러 가지 c y t o k i n e s과 유착분자 등의 발현 을 증가시킬 수 있는 것으로 알려져 있다3 1 ). 본 연구 에서는 활막 섬유모세포를 3시간 동안 저산소증 상태 에 노출시킨 결과 N F -κB가 활성화됨을 관찰할 수 있 었고, 활막 섬유모세포의 배양 상층액에 항산화제인 P D T C를 전처치 한 경우에서는 저산소증에 의한 N F -κB의 활성화가 억제되는 것을 알 수 있었다. 이 는 저산소증이 N F -κB의 활성화를 통해서 I C A M - 1의 발현을 직접 활성화 할 수 있음을 보여주는 것이다.
I L - 1과 T N F -α는 N F -κB의 natural inducer로 서 잘 알려져 있으며, 이러한 c y t o k i n e s의 발현에는 N F -κB가 관여하는 것으로 알려져 있으므로 저산소 증에 의한 세포내 전사인자인 N F -κB의 활성화와 저 산소증에 의한 I L - 1β, TNF-α의 증가는 활막 섬유 모세포에서 I C A M - 1의 발현에 복합적으로 관련될 수 있을 것으로 추정할 수 있다. 이에 대한 정확한 기전에 대해서는 I L - 1β, TNF-α등에 대한 단일클 론 항체를 이용한 실험을 통해 c y t o k i n e s이 저산소 증에 의한 I C A M - 1의 발현에 미치는 영향을 알아볼 필요가 있을 것으로 보인다.
결론적으로, 저산소증은 활막 섬유모세포에서 I C A M - 1의 발현을 유도할 수 있으며, 여기에는 세포 내의 전사인자인 N F -κB의 활성화가 관련되는 것으 로 보인다. 저산소증에 의해 자극된 활막 섬유모세포 가 림프구와 유착반응에 의해 결합이 증가하는 것은
저산소증이 활막에 염증세포의 침착에 관여하는 것으 로 생각된다. 이와 같이 저산소증은 류마티스 관절염 에서 면역반응에 영향을 주어 병인에 중요한 역할을 하는 것으로 보이며, 항 산화제와 같은 저산소증과 관련된 일련의 과정을 조절할 수 있는 방법이 류마티 스 질환의 치료에 도움을 줄 수 있을 것으로 여겨지
고 있고3 9 , 4 0 ), 이를 류마티스 관절염의 치료에 이용하
려는 연구들이 진행 중이므로 새로운 형태의 치료방 법이 될 수 있을 것으로 여겨지지만 이에 대해서는 보다 많은 연구가 필요할 것으로 사료된다.
결 론
류마티스 관절염은 유전적 요인, 환경적 요인과 면역세포의 이상 등이 복합적으로 작용하여 발생한 다고 생각하고 있으나, 아직도 활막염의 원인과 염 증이 지속적으로 진행하는 기전에 대해서는 아직 명 확히 밝혀져 있지 않다. 활막 조직에의 염증세포의 지속적인 침윤이 류마티스 관절염의 병인에 중요한 역할을 할 것으로 생각되며, 여기에는 염증세포와 혈관의 내피세포 및 활막 섬유모세포 간의 상호반응 과 cytokine, 유착분자 등이 중요한 역할을 하는 것 으로 보고되고 있다.
본 연구에서는 류마티스 관절염 환자의 활막 섬유 모세포를 저산소 상태에서 배양하였을 때, ICAM-1 의 발현 및 N F -κB의 활성화와 림프구와의 유착반응 이 증가하는 것을 보여 주고 있다. 이는 활막의 저산 소증이 염증반응에 관련된 단백질의 유전자 발현에 영향을 줄 수 있고, 활막조직에의 염증세포 침윤 및 활막 섬유모세포와 염증세포간의 상호반응에 중요한 역할을 할 수 있음을 보여주는 결과이다.
결론적으로, 저산소증은 류마티스 관절염에서 염 증반응에 영향을 주는 것으로 보아 병인에 중요한 역할을 하는 것으로 보이며, 저산소증과 관련된 과 정을 조절할 수 있는 방법을 치료법의 하나로서 이 용할 수 있는가에 대해서는 보다 많은 연구가 필요 할 것으로 사료된다.
R E F E R E N C E S
11 ) Keyszer GM, Heer AH, Gay S. Cytokines and oncogenes in cellular interactions of rheuma-
toid arthritis. Stem Cells 1994;12:75-86.
12 ) Gay S, Gay RE, Koopman WJ. Molecular and cellular mechanisms of joint destruction in rheumatoid arthritis: two cellular mechanisms explain joint destruction? Ann Rheum Dis 1 9 9 3 ; 5 2 : S 3 9 - 4 7 .
13 ) Harris ED Jr. Rheumatoid arthritis. Pathophysi- ology and implications for therapy. N Engl J Med 1990;322:1277-89.
14 ) Sewell KL. Immunotherapy and other novel therapies, including biologic response modifiers, apheresis, and dietary modifications. Curr Opin Rheumatol 1993;5:293-8.
15 ) Kunkel SL, Lukacs N, Kasama T, Strieter RM. The role of chemokines in inflammatory joint disease. J Leukoc Biol 1996;59:6-12.
16 ) Brennan FM, Feldmann M. Cytokines in autoimmunity. Curr Opin Immunol. 1992;4:
7 5 4 - 9 .
17 ) Cavender D, Saegusa Y, Ziff M. Stimulation of endothelial cell binding of lymphocytes by tumor necrosis factor. J Immunol 1987;139:1855-60.
18 ) Hale LP, Martin ME, McCollum DE, Nunley JA, Springer TA, Singer KH, et.al. Immuno- histologic analysis of the distribution of cell adhesion molecules within the inflammatory synovial microenvironment. Arthritis Rheum 1 9 8 9 ; 3 2 : 2 2 - 3 0 .
19 ) Marlor CW, Webb DL, Bombara MP, Greve JM, Blue ML. Expression of vascular cell adhe- sion molecule-1 in fibroblastlike synoviocytes after stimulation with tumor necrosis factor.
Am J Pathol 1992;140:1055-60.
1 0 ) Panes J, Perry MA, Anderson DC, Manning A, Leone B, Cepinskas G, et al. Regional differences in constitutive and induced ICAM-1 expression in vivo. Am J Physiol 1995;269: H1955-64.
1 1 ) Paleolog EM, Young S, Stark AC, McCloskey RV, Feldmann M, Maini RN. Modulation of angiogenic vascular endothelial growth factor by tumor necrosis factor alpha and interleukin-1 in rheumatoid arthritis. Arthritis Rheum 1998;41:
1 2 5 8 - 6 5 .
1 2 ) Stevens CR, Blake DR, Merry P, Revell PA, Levick JR. A comparative study by morphome- try of the microvasculature in normal and rheumatoid synovium. Arthritis Rheum 1992;34:
1 5 0 8 - 1 3 .
1 3 ) Allen RE, Blake DR, Nazhat NB, Jones P.
Superoxide radical generation by inflamed human synovium after hypoxia. Lancet 1 9 8 9 ; 2 9 ; 2 : 2 8 2 - 3 .
1 4 ) Unsworth J, Outhwaite J, Blake DR, Morris CJ, Freeman J. Dynamic studies of the rela- tionship between intraarticular, synovial fluid oxygen tension, and lipid peroxidation in the inflamed knee: an example of reperfusion injury. Ann Clin Biochem 1988;25:S8-11.
1 5 ) Blake DR, Merry P, Unsworth J, Kidd BL, Outhwaite JM, Ballard R, et. al. Hypoxic - reperfusion injury in the inflamed human joint.
Lancet 1989;11;1:289-93.
1 6 ) Wood JG, Johnson JS, Mattioli LF, Gonzalez NC. Systemic hypoxia promotes leukocyte- endothelial adherence via reactive oxidant generation. J Appl Physiol 1999;87;5:1734-40.
1 7 ) Cepinskas G, Lush CW, Kvietys PR. Anoxia/
reoxygenation-induced tolerance with respect to polymorphonuclear leukocyte adhesion to cul- tured endothelial cells. A nuclear factor-kap- paB-mediated phenomenon. Circ Res 1999;84:
1 0 3 - 1 2 .
1 8 ) Arnould T, Michiels C, Remacle J. Increased PMN adherence on endothelial cells after hypoxia: involvement of PAF, CD18/CD11b, and ICAM-1. Am J Physiol 1993;264:C1102-10.
1 9 ) Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF, Cooper NS, et. al. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis.
Arthritis Rheum 1988;31:315-24.
2 0 ) Matsuoka N, Eguchi K, Kawakami A, Ida H, Nakashima M, Sakai M, et. al. Phenotypic characteristics of T cells interacted with synovial cells. J Rheumatol 1991;18:1137-42.
2 1 ) Rosengard BR. and Cochrane DE. Comple- ment-mediated cytolysis: A quick, simple method for determining levels of immunoglob- ulin E bound to mast cells. J. Histochem.
Cytochem 1983;131:441-4.
2 2 ) Oppenheimer-Marks N, Davis LS, Bogue DT, Ramberg J, Lipsky PE. Differential utilization of ICAM-1 and VCAM-1 during the adhesion and transendothelial migration of human T lymphocytes. J Immunol 1991;147:2913-21.
2 3 ) Schreck R, Meier B, Mannel DN, Droge W, Baeuerle PA. Dithiocarbamates as potent inhibitors of nuclear factor kappa B activation
in intact cells. J Exp Med 1992;175:1181-94.
2 4 ) Grober JS, Bowen BL, Ebling H, Athey B, Thompson CB, Fox DA, et al. Monocyte - endothelial adhesion in chronic rheumatoid arthritis. In situ detection of selectin and inte- grin-dependent interactions. J Clin Invest 1993;
9 1 : 2 6 0 9 - 1 9 .
2 5 ) Ishikawa H, Nishibayashi Y, Kita K, Ohno O, Imura S, Hirata S. Adhesion molecules in the lymphoid cell distribution in rheumatoid syn- ovial membrane. Bull Hosp Jt Dis 1993;53:23-8.
2 6 ) Dingle JTM, Page-Thomas DP. In vitro stud- ies in human synovial membrane. A metabolic comparison of normal and rheumatoid disease.
Br J Exp pathol 1956;37: 318-36.
2 7 ) Roberts JE, McLee BD, Kerby GP. Pathways of glucose metabolism in rheumatoid and non- rheumatoid synovium. J Lab Clin Med 1967;70:
5 0 3 - 1 1 .
2 8 ) Falchuk KH, Goetzl EJ, Kulka JP. Respiratory gases of synovial fluids. An approach to syn- ovial tissue circulatory-metabolic imbalance in rheumatoid arthritis. Am J Med 1970;49:223- 3 1 .
2 9 ) McNeil JD, Wiebkin OW, Betts WH, Cleland LG. Depolymerisation products of hyaluronic acid after exposure to oxygen-derived free radicals. Ann Rheum Dis 1985; 44:780-9.
3 0 ) Mapp PI, Grootveld MC, Blake DR. Hypoxia, oxidative stress and rheumatoid arthritis. Br Med Bull 1995;51:419.
3 1 ) Baeuerle PA, Baltimore D. The physiology of N F -κB transcription factor. In molecular aspects of cellular regulation. Vol. 6. The hormonal con- trol of gene transcription. P. Cohen & JG Foulkes, eds. p. 423-46, 1991.
3 2 ) Koong AC, Chen EY, Giaccia AJ. Hypoxia causes the activation of nuclear factor kappa B through the phosphorylation of I kappa B alpha
on tyrosine residues. Cancer Res 1994;54:1425- 3 0 .
3 3 ) Karin M. Signal transduction and gene con- trol. Curr Opin Cell Biol. 1991;3:467-73.
3 4 ) Shreeniwas R, Koga S, Karakurum M, Pinsky D, Kaiser E, Brett J, et al. Hypoxia- mediated induction of endothelial cell interleukin-1 alpha.
An autocrine mechanism promoting expression of leukocyte adhesion molecules on the vessel sur- face. J Clin Invest 1992;90:2333-9.
3 5 ) Ala Y, Palluy O, Favero J, Bonne C, Modat G, Dornand J. Hypoxia/reoxygenation stimulates endothelial cells to promote interleukin-1 and interleukin-6 production. Effects of free radical scavengers. Agents Actions 1992;37:134-9.
3 6 ) Lindsley HB, Smith DD, Davis LS, Koch AE, Lipsky PE. Regulation of the expression of adhesion molecules by human synoviocytes.
Semin Arthritis Rheum 1992;21:330-4.
3 7 ) Bombara MP, Webb DL, Conrad P, Marlor CW, Sarr T, Ranges GE, et al. Cell contact between T cells and synovial fibroblasts causes induction of adhesion molecules and cytokines.
J Leukoc Biol 1993;54:399-406.
3 8 ) Chin JE, Winterrowd GE, Krzesicki RF, Sanders ME. Role of cytokines in inflammatory synovitis. The coordinate regulation of intercel- lular adhesion molecule 1 and HLA class I and class II antigens in rheumatoid synovial fibrob- lasts. Arthritis Rheum. 1990;33:1776-86.
3 9 ) Darlington LG, Stone TW. Antioxidants and fatty acids in the amelioration of rheumatoid arthritis and related disorders. Br J Nutr 2001;
8 5 : 2 5 1 - 6 9 .
4 0 ) Kerimova AA, Atalay M, Yusifov EY, Kuprin SP, Kerimov TM. Antioxidant enzymes; possible mechanism of gold compound treatment in rheumatoid arthritis. Pathophysiology 2000;7:
2 0 9 - 2 1 3 .