161
ABBREVIATIONS: SCs, Schwann cells; DMEM, Dulbecco’s modified Eagle’s medium; FBS, fetal bovine serum; TBS, tris-buffered saline;
ERK, extracellular signal-regulated kinase; IL-6, interleukin-6; JAK, janus kinase; STAT, signal transducer and activator of transcription;
PBS, phosphate-buffered saline; NRG, neuregulin; MEK, mitogen- activated protein kinase/extracellular signal-regulated kinase kinase.
Received April 2, 2009, Revised May 13, 2009, Accepted May 26, 2009
Corresponding to: Hwan Tae Park, Department of Physiology, Medical Science Research Institute, College of Medicine, Dong-A University, 3-1, Dongdaesin-dong, Seo-gu, Busan 602-714, Korea.
(Tel) 82-51-240-2636, (Fax) 82-51-246-6481, (E-mail) phwantae@dau.
ac.kr
Extracellular Signal-regulated Kinase Activation Is Required for Serine 727 Phosphorylation of STAT3 in Schwann Cells in vitro and in vivo
Hyun Kyoung Lee1, Junyang Jung1, Sang Hwa Lee2, Su-Yeong Seo2, Duk Joon Suh1, and Hwan Tae Park1 Departments of 1Physiologyand 2Microbiology, Medical Science Research Institute, College of Medicine, Dong-A University, Busan 602-714, Korea
In the peripheral nerves, injury-induced cytokines and growth factors perform critical functions in the activation of both the MEK/ERK and JAK/STAT3 pathways. In this study, we determined that nerve injury-induced ERK activation was temporally correlated with STAT3 phosphorylation at the serine 727 residue. In cultured Schwann cells, we noted that ERK activation is required for the serine phosphorylation of STAT3 by neuropoietic cytokine interleukin-6 (IL-6). Serine phosphorylated STAT3 by IL-6 was transported into Schwann cell nuclei, thereby indicating that ERK may regulate the transcriptional activity of STAT3 via the induction of serine phosphorylation of STAT3. Neuregulin-1 (NRG) also induced the serine phosphorylation of STAT3 in an ERK-dependent fashion. In contrast with the IL-6 response, serine phosphorylated STAT3 induced by NRG was not detected in the nucleus, thus indicating the non-nuclear function of serine phosphorylated STAT3 in response to NRG. Finally, we determined that the inhibition of ERK prevented injury-induced serine phosphorylation of STAT3 in an ex-vivo explants culture of the sciatic nerves. Collectively, the results of this study show that ERK may be an upstream kinase for the serine phosphorylation of STAT3 induced by multiple stimuli in Schwann cells after peripheral nerve injury.
Key Words: STAT3, ERK, Schwann cells, Interleukin-6, Neuregulins-1, Nerve injury
INTRODUCTION
In the peripheral nervous system, the differentiation of Schwann cells (SCs) during postnatal development involves the loss of immature phenotypes and axonal myelination (Jessen and Mirsky, 2005). In the peripheral nerves of adult animals, myelinating SCs are a type of regenerative cells, which undergo a dramatic alteration after nerve injury (Jessen and Mirsky, 2008). They lose their myelinating phe- notypes via downregulation of myelin gene expression and acquire the properties of immature SCs, which can be iden- tified by the upregulation of several proteins including glial fibrillary acidic protein and the p75 neurotrophin receptor.
This dedifferentiation of SCs in response to nerve damage is a crucial component of Wallerian degeneration, and is an essential reaction of SCs to myelin removal and axonal regeneration (Fu and Gordon, 1997). Although a great many factors are capable of inducing the phenotype changes of SCs in vitro, the intracellular signaling associated with SC dedifferentiation has yet to be thoroughly elucidated. It was reported recently that the Ras/extracellular signal-regu- lated kinase (ERK) pathway might drive the dediffer- entiation of SCs in vitro (Harrisingh et al., 2004). A forceful
activation of ERK led to the downregulation of markers of myelinating SCs, including as P0 and periaxin. ERK activa- tion in the injured nerves in vivo further supports the no- tion that ERK signaling plays an important role in the phe- notype changes occurring in myelinating SCs after periph- eral nerve injury (Sheu et al., 2000; Harrisingh et al., 2004).
Signal transducer and activator of transcription 3 (STAT3) is a critical regulator of gene expression in response to a variety of cytokines (Battle and Frank, 2002). The exposure of cells to interleukin-6 (IL-6) or ciliary neurotrophic factors results in STAT3 activation via STAT3 phosphorylation at the tyrosine 705 and serine 727 residues (Kamimura et al., 2003). Phosphorylated STAT3 dimerizes and translocates to the nucleus, where it regulates the transcription of sev- eral apoptosis- and cell-cycle-associated genes (Reich and Liu, 2006). The tyrosine phosphorylation of STAT3 has been shown to be increased in the peripheral nerves follow- ing injury, and the activation of STAT3 in the dorsal root ganglion neurons performs a significant function in re- generative axonal growth (Cafferty et al., 2001; Lee et al., 2004; Qui et al., 2005). STAT3 activation has also been de- tected in SCs following injury (Lee et al., 2004), and we recently reported that STAT3 activation in SCs might be involved in the induction of a marker for dedifferentiated
SCs, glial fibrillary acidic protein (Lee et al., 2009).
Many factors can activate the Ras/ERK pathway in SCs after nerve injury. For example, neuregulins (NRG) may activate ERK for demyelination (Harrisingh et al., 2004).
We recently reported that IL-6 activated ERK in SC-de- rived RT4 cells and primary SCs (Lee et al., 2009), even though the role of IL-6-induced ERK activation currently remains unknown. In this study, we placed our focus on the possible role of the Ras/ERK pathway in STAT3 phos- phorylation in primary SCs and RT4 cells (Hai et al., 2002), because it has already been established that the Ras/ERK pathway is either positively or negatively involved in STAT3 activation. We determined that ERK is an upstream kinase for the serine 727 phosphorylation of STAT3 in SCs in vitro and in vivo.
METHODS Chemicals
Media and sera for cell cultures were obtained from Life Technologies (Grand Island, NY). All of the phospho-specific antibodies and recombinant IL-6 utilized herein were pur- chased from Cell Signaling Technology (Beverly, MA). STAT3, ERK and beta-tubulin antibodies were acquired from Santa Cruz Biotechnology (Santa Cruz, CA). An antibody against S100 was purchased from Chemicon (Temecula, CA). Elec- trophoresis and Western blot reagents were obtained from BioRad (Richmond, VA). Neuregulin-1 (NRG) was purchased from R&D systems (Minneapolis, MN). U0126 and PD98059 were purchased from Calbiochem (San Diego, CA), and all other reagents were acquired from Sigma (St. Louis, MO).
Sciatic nerve injury and immunohistochemistry In accordance with the protocols approved by the Dong-A University Committee on animal research that followed the guide of animal experiments established by The Korean Academy of Medical Sciences, adult male Sprague-Dawley rats (150∼300 gm) were anesthetized with an intraperitoneal injection of 10% ketamine hydrochloride (Sanofi-Ceva, Düsseldorf, Germany; 0.1 ml/100 g body weight) and 0.1 ml of Rompun (Bayer, Leverkusen, Germany). For sciatic nerve lesions, the right sciatic nerve was exposed at the mid-thigh level and was crushed twice in succession using a fine forceps. After recovery for the indicated time, the rats were sacrificed with a high dose of a mixture of 10%
ketamine and Rompun, and then the control and crushed nerves were removed and frozen with dryice. Frozen nerve sections (12 μm) were prepared using a cryostat (Leica CM3050), air-dried, and fixed for 20 min with methanol at 4oC. The immunostaining of phosphorylated STAT3 was conducted in accordance with the previously described pro- tocols (Lee et al., 2009). In brief, the sections were blocked with 4% goat serum in phosphate-buffered saline (PBS) containing 0.3% Triton X-100 for 15 min at room temper- ature, and then incubated overnight with a rabbit anti- phospho-STAT3 antibody (1:200) in PBS containing 0.2%
Triton X-100 at 4oC followed by 3 washings with PBS. Next, the sections were incubated for 2 h with Alexa 488-conjugated anti-rabbit IgG (1:800, Jackson Immunoresearch Labora- tories) at room temperature, and visualized with a laser confocal microscope (LSM510, Carl Zeiss, Germany). The sections were labeled with DAPI nuclear stain to visualize
nuclei in the sections.
Western blot analysis
In order to prepare protein lysates from the injured nerves, the proximal and distal stumps (10 mm length) were removed and homogenized with a polytron homoge- nizer in modified radioimmune precipitation assay (RIPA) lysis buffer (150 mM NaCl, 1% Nonidet P-40, 1 mM EDTA, 0.5% deoxycholic acid, 2 μg/ml aprotinin, 1 mM phenyl- methylsulfonyl fluoride, 5 mM benzamidine, 1 mM sodium orthovanadate). The lysates were centrifuged for 15 min at 9,000 g at 4oC, and then the supernatant was employed for Western blot analysis. The proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred onto a nitrocellulose membrane (Amer- sham Biosciences, Piscataway, NJ). After 1 h of blocking with 0.1% Tween 20 and 5% nonfat dry milk in Tris-buf- fered saline (TBS, 25 mM Tris-HCl pH 7.5, 140 mM NaCl) at room temperature, the membrane was incubated with primary antibodies (1:500∼1,000) at 4oC overnight. After three 15 min washes with TBS containing 0.1% Tween 20 (TBST), the membranes were incubated with a a horse- radish peroxidase-conjugated secondary antibody (1:3,000) for 1 h at room temperature. The signals were detected with an Enhanced Chemiluminescence System (ECL Advance kit, Amersham Biosciences).
For quantification, the X-ray films were then subsequently scanned with an HP scanner and analyzed using an LAS image analysis system (Fujifilm, Japan). The intensities of the bands were normalized to those of beta-tubulin or non- phosphorylated STAT3 in three independent experiments.
Primary Schwann cell cultures and RT4 cells For the experiment using SCs, primary SCs from adult sciatic nerves and RT4 cells were employed. Primary SCs were obtained from adult rat sciatic nerves as previously reported (Lee et al., 2007). Briefly, the sciatic nerves of adult Sprague-Dawley rats were axotomized to enhance the SC population. Sciatic nerves were sectioned 5 mm proximal to the tibioperoneal bifurcation with a fine iris scissor (FST Inc, Foster City, CA, USA) and the animals were housed in plastic cages for 3∼4 days after injury. The sciatic nerves were removed and subjected to chemical digestion in 0.2% collagenase for 2 h in calcium/magnesium-free Hank’s buffered solution at 37oC. The nerves were then dis- sociated via 2 min of gentle shaking followed by two or three triturations performed with a flame-polished Pasteur pipette. The cell pellets obtained after centrifugation were then resuspended in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% (vol/vol) heat-inactivated fetal bo- vine serum (FBS), plated at a density of 20,000 cells/cm2 and grown for 2 days at 37oC in a humidified atmosphere containing 5% CO2. After two or three subcultures using the coldjet method (Jirsova et al., 1997), the identity and purity of SCs were evaluated via immunostaining with an antibody raised against S100 (data not shown).
The rat schwannoma cell line (RT4, CRL-2768) was obtained from the American Type Culture Collection (Rockville, MD) and maintained as previously described (Lee et al., 2009).
Immunofluorescent staining
The cells were starved for 4 h or overnight in serum-free
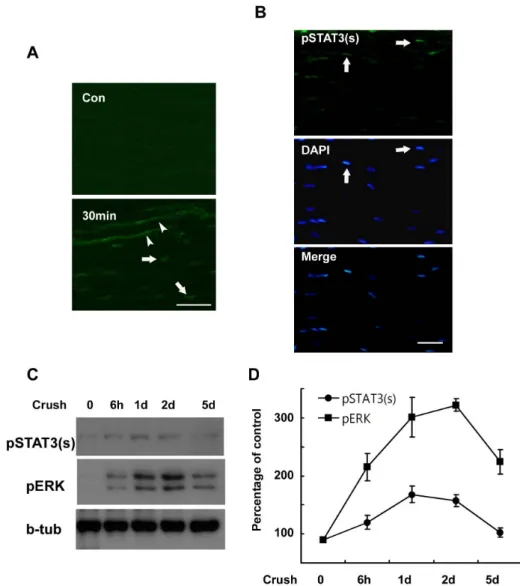
Fig. 1. Serine phosphorylation of STAT3 in the sciatic nerve following crush injury. (A, B) Frozen sections from control and injured sciatic nerves were immunostained with an anti- body against serine phosphorylated STAT3 (pSTAT3(s)). Axons (arrow- heads) and nuclei of SC-like cells (arrows) display a rapid induction in the serine phosphorylation of STAT3 at 30 min (A) and 60 min (B) after lesion. Scale bar; 30 μm. (C) Protein lysates were prepared from the injured sciatic nerves and analyzed with Western blot analysis for detec- tion of serine phosphorylated STAT3 and pERK. (D) A quantitative analy- sis for result (C).
DMEM supplemented with 0.5% FBS prior to the addition of cytokines or trophic factors at the concentrations in- dicated in the Results section. For ERK inhibition, the cells were pretreated with U0126 (10 μM) or PD98059 (20 μM) for 30 min before the addition of IL-6 or growth factors.
After stimulation with IL-6 or NRG, the cells were fixed for 15 min with ice-cold methanol at 4oC for pSTAT3 im- munostaining and then washed three times with PBS. The cells were blocked with PBS containing 0.2% Triton X-100 and 2% bovine serum albumin for 1 h. Primary SCs were double immunostained with a mouse anti-S100 antibody (1:200) and a rabbit anti-phospho-STAT3 antibody (1:200) for 16 h at 4oC. Next, the cells were incubated for 2 h with Alexa 488-conjugated anti-rabbit IgG and Texas Red-la- beled donkey anti-mouse IgG at room temperature, then viewed under a laser confocal microscope.
Ex-vivo explant culture
Sciatic nerves from adult rats were removed and the con- nective tissues surrounding the nerves were carefully re- moved in calcium/magnesium-free Hank’s buffered solution under a stereomicroscope. The sciatic nerves were longi-
tudinally cut into 2 or 3 nerve explants and were then cut into small explants of 3 mm length. The explants were maintained for 6 h or 24 h in DMEM containing 10% FBS at 37oC in a humidified atmosphere containing 5% CO2.
RESULTS
Serine phosphorylation of STAT3 in the injured sciatic nerves
The results of previous studies have demonstrated STAT3 activation in injured peripheral nerves by detecting phos- phorylation of STAT3 at tyrosine residue 705 (pSTAT3(y), Sheu et al., 2000; Lee et al., 2004). However, phosphor- ylation of STAT3 at serine residue 727 (pSTAT3(s)) has yet to be investigated in the sciatic nerve after injury. We ini- tially assessed the serine phosphorylation of STAT3 in in- jured sciatic nerves using immunofluorescent staining.
pSTAT3(s) was not detected in the control sciatic nerves, but was clearly observed in the nuclei of SC-like cells and damaged axoplasm within 30 min at lesion sites after sci- atic nerve injury (Fig. 1A). Increased pSTAT3(s) staining
Fig. 2. IL-6 induces serine phos- phorylation of STAT3 in Schwann cells. RT4 schwannoma cells (A∼D) and primary SCs (E, F) were serum starved overnight and then treated with IL-6 (50 ng/ml) for the indicated times. Total cell lysates were sub- jected to Western blot analysis using antibodies specific for phosphory- lated proteins. (A, B) RT4 cells were treated with IL-6 for 15 min, and the cellular lysates were analyzed using the indicated antibodies. (B) A quan- titative analysis of tyrosine phospho- rylated STAT3 in result (A). (C, D) RT4 cells were treated with IL-6 for the indicated time in the presence or absence of U0126. (D) A quantitative analysis of serine phosphorylated STAT3 in result (C). (E, F) Primary SCs were treated with IL-6 for 15 min, and the total lysates were analyzed by Western blot analysis using the indicated antibodies. (F) A quantita- tive analysis of phosphorylated STAT3 in result (E).
was also noted in the nuclei of SC-like cells in the proximal and distal stump of the injured nerves within 60 min of injury (Fig. 1B), thereby indicating that serine phosphor- ylation of STAT3 occurs in damaged axons and SC-like cells of injured nerves in vivo. In order to know determine the temporal profile of serine phosphorylation of STAT3 in the sciatic nerve after injury, we conducted Western blot analy- sis (Fig. 1C). This experiment demonstrated that the serine phosphorylation of STAT3 peaked at 1 day and persisted until 2 days after the induced crush injury. Because ERK is known to be activated in injured sciatic nerves (Harri- singh et al., 2004), we assessed the activation profile of
ERK after the crush injury. Interestingly, the temporal pro- file of ERK activation observed after nerve injury was sim- ilar to that of STAT3 serine phosphorylation, particularly during the early post-injury period (Fig. 1D), thereby sug- gesting that ERK may be associated with the serine phos- phorylation of STAT3 following nerve injury.
Interleukin-6 induces serine phosphorylation of STAT3 in an ERK-dependent manner
In order to characterize the relationship between ERK and the serine phosphorylation of STAT3 in SCs, we em-
Fig. 3. NRG-1 induces serine phos- phorylation of STAT3 in Schwann cells. (A, B) Primary SCs were serum starved overnight and then treated with NRG-1 (200 ng/ml) for the indi- cated times. Total cell lysates were subjected to Western blot analysis using antibodies specific for phos- phorylated proteins. (B) A quantita- tive analysis of serine phosphory- lated STAT3 in result (A). (C, D) Primary SCs were treated with NRG-1 for 15 min in the presence or absence of U0126 or DMSO, and cellular lysates were analyzed with Western blot analysis. (D) A quanti- tative analysis of serine phosphory- lated STAT3 in result (C).
ployed RT4 schwannoma cells and primary SCs cultured from adult rat sciatic nerves. Many stimuli can be involved in the activation of ERK in SCs, and we previously reported that IL-6 activated ERK in RT4 cells and primary SCs (Lee et al., 2009). These findings led us to evaluate the role of ERK in phosphorylation of STAT3 in response to IL-6.
Consistent with the previous result (Lee et al., 2009), we found an activation of ERK by IL-6 within 5 min in the RT4 cells (Fig. 2A, C). When the RT4 cells were pretreated with the mitogen-activated protein kinase/extracellular signal- regulated kinase kinase (MEK) inhibitors, U0126 (10 μM) or PD980519 (20 μM, data not shown), before the stimulation with IL-6, a complete reduction in the phosphorylation of ERK was noted (Fig. 2A, C). However, the tyrosine phos- phorylation of STAT3 by IL-6 was not altered by the inhibition of MEK (Fig. 2A, B), suggesting that ERK activation is not involved in the IL-6-induced tyrosine phosphorylation of STAT3. In the RT4 cells, IL-6 induced an increase in the level of serine phosphorylation of STAT3 even though this serine phosphorylation was not as robust as the tyrosine phosphorylation (Fig. 2C). The temporal profile of STAT3 serine phosphorylation was similar to that of ERK activation, and treatment with the specific MEK inhibitors resulted in the blockage of IL-6-induced STAT3 serine phosphorylation (Fig. 2C, D). The involvement of ERK in the serine phos- phorylation, but not the tyrosine phosphorylation, of STAT3 was also detected in primary SCs cultured from adult sciatic nerves (Fig. 2E, F). This finding shows that IL-6-mediated ERK activation is involved in STAT3 phosphorylation at the serine 727 residue in RT4 cells and primary SCs.
Neuregulin-1 induces serine phosphorylation of STAT3 in an ERK-dependent manner
It was previously reported that NRG is an important acti- vator of the MEK/ERK pathway in SCs (Harrisingh et al., 2004). However, the role of NRG-induced ERK activation in the serine phosphorylation of STAT3 in SCs had, until present, not yet been thoroughly investigated. Thus, we at- tempted to determine whether NRG-1 induces serine phos- phorylation of STAT3 through ERK activation using primary SCs. NRG-1 (200 ng/ml) induced not only ERK activation but also serine phosphorylation of STAT3 in a time-dependent manner (Fig. 3A, B). However, the tyrosine phosphorylation of STAT3 was not changed after NRG-1 treatment as pre- viously reported (data not shown, Lee et al., 2009). The serine phosphorylation appeared within 5 min and peaked 15 min after NRG treatment. MEK inhibition with U0126 completely inhibited serine phosphorylation of STAT3 induced by NRG-1 (Fig. 3C, D). These findings show that the NRG-1- induced activation of the MEK/ERK pathway is involved in the serine phosphorylation of STAT3 in primary SCs.
Differential localization of serine phosphorylated STAT3 by IL-6 and neuregulin-1
We then attempted to determine whether IL-6 induces the nuclear translocation of serine phosphorylated STAT3, and did indeed detect serine-phosphorylated STAT3 in the nucleus within 20 min of IL-6 treatment (Fig. 4A). Treat-
Fig. 5. ERK inhibition reduces the serine phosphorylation of STAT3 in explant cultures of sciatic nerves.
(A, B) Sciatic nerve explants were cultured for 6 or 24 h in the presence or absence of U0126, and then the protein lysates were analyzed with Western blot analysis using an anti- body against serine phosphorylated STAT3 and anti-pERK antibody.
(B) A quantitative analysis of serine phosphorylated STAT3 in result (A).
Fig. 4. Differential localization of phosphorylated STAT3 by IL-6 and NRG-1. (A) After 20 min of stimulation with IL-6, RT4 cells were fixed and immunostained for serine phosphorylated STAT3.
Con; untreated control. (B) RT4 cells were pretreated with U0126 for 30 min and then stimulated with IL-6 for 20 min. Cells were immunostained for tyrosine or serine phosphorylated STAT3. Scale bar; 30 μm. (C) Primary SCs were treated with IL-6 or NRG for 20 min, and the cells were fixed and double immunostained for a SC marker (S100) and phosphorylated STAT3. Scale bar; 30 μm.
ment with the MEK inhibitor U0126 did not affect a marked increase in pSTAT3(y) by IL-6 in the nucleus, whereas it prevented the appearance of serine phosphorylated STAT3 (Fig. 4B), in a finding consistent with the biochemical results (Fig. 2C). Additionally, the identical results were observed with the cultured primary SCs (Fig. 4C).
Although the tyrosine phosphorylation of STAT3 is a pre- requisite for the nuclear translocation of STAT3 in many situations, the nuclear translocation of serine phosphory- lated STAT3 (without tyrosine phosphorylation) has also been reported in previous studies (Ng et al., 2006b; Reich and Liu, 2006). Thus, we examined the possibility of that the nuclear translocation of serine phosphorylated STAT3 by NRG-1 might occur in primary SCs (Fig. 4C). Double immunofluorescent staining with an antibody against a SC marker (S100) and an antibody against pSTAT3(y) or pSTAT3(s) showed obvious nuclear translocation of pSTAT3(y) and pSTAT3(s) by IL-6 in S100-positive cells. However, NRG-1 treatment was not shown to induce an increase in the nuclear staining of either pSTAT3(y) or pSTAT3(s).
Rather, NRG generally increased the immunolabeling of pSTAT3(s) in the cytoplasm without distinct nuclear staining.
Thus, our data suggest that NRG-1 cannot induce the nuclear translocation of pSTAT3(s).
ERK inhibition reduces serine phosphorylation of STAT3 in an ex-vivo sciatic nerve injury model We have determined that ERK is a crucial upstream kin- ase for the IL-6 and NRG-induced serine phosphorylation of STAT3 in cultured SCs. In order to know the function of ERK in the serine phosphorylation of STAT3 in the in- jured nerve itself, we utilized an ex-vivo sciatic nerve injury model. When sciatic nerves are removed from animals and cultured as explants, the nerve explants are known to un- dergo Wallerian degeneration in a manner very similar to that seen under in vivo conditions (Thomson et al., 1993;
Bleuel and Monard, 1995). We cultured sciatic nerve ex- plants in the presence or absence of the MEK inhibitor, U0126 (20 μM), for 6 h and 24 h, after which the protein lysates from the explants were assessed with Western blot analysis (Fig. 5A). This experiment showed that ERK and STAT3 were activated during explant culture, and that the inhibition of ERK significantly reduced the serine phos- phorylation of STAT3 (Fig. 5), thus suggesting that ERK performs an important function in the serine phosphor- ylation of STAT3 after nerve injury.
DISCUSSION
It was previously reported that STAT3 in axons and SC-like cells is activated by peripheral nerve lesion (Sheu et al., 2000; Lee et al., 2004; Lee et al., 2009). The activation of STAT3 at lesion sites was demonstrated by the observed tyrosine phosphorylation of STAT3 and occurred within 30 min of injury. The present data demonstrate, for the first time, that the serine phosphorylation of STAT3 also oc- curred in axons and SC-like cells in the peripheral nerves following lesion. According to previous studies that demon- strated STAT3 activation in the peripheral nerves after the development of a lesion, the temporal and spatial profiles of tyrosine phosphorylation of STAT3 in the injured nerves were similar to that associated with the serine phosphor- ylation of STAT3. Thus, it appears that STAT3 can be phos- phorylated simultaneously at tyrosine 705 and serine 727 residues in the injured peripheral nerves. It has been pre- viously reported that the tyrosine phosphorylation of STAT3 is an essential step for the dimerization of STAT3.
Because dimerization is required for the binding of STAT3 to DNA promoter regions, tyrosine phosphorylation is a pre- requisite for STAT3-dependent transcriptional activation (Battle and Frank, 2002). In addition to the tyrosine phos- phorylation of STAT3, phosphorylation of STAT3 at the ser- ine 727 residue has been suggested to play an important role in the regulation of transcriptional activity of STAT3.
For example, it was reported that the serine phosphor- ylation of STAT3 results in the maximal transcriptional ac- tivation of tyrosine phosphorylated STAT3 (Wen et al., 1995; Plaza-Menacho et al., 2007). On the contrary, several papers have demonstrated that the serine phosphorylation of STAT3 exerts a negative effect on the transcriptional ac- tivity and tyrosine phosphorylation of STAT3 (Chung et al., 1997; Jain et al., 1998). Thus, the functional significance of serine phosphorylation of STAT3 may differ from cell to cell. Considering the role of serine phosphorylation of STAT3 in its transcriptional activity, we think that the ser- ine phosphorylation of STAT3 in SCs may play a significant role in STAT3 function in the injured sciatic nerves.
Because the role of STAT3 in SCs after nerve injury re- mains largely unknown at present, further studies into the function of STAT3 in SCs are required for a great under- standing of the functional significance of serine phosphor- ylation of STAT3 in SCs and in peripheral nerve regene- ration.
After nerve injury, there is an initial degradation of mye- lin sheath and subsequent phenotype changes of SCs into immature states. This process is called dedifferentiation (Jessen and Mirsky, 2008). This regenerative change of SCs is essential for successful nerve regeneration. The dediffer- entiation of SCs after injury involves the activation of sev- eral signaling pathways to alter gene expression profiles.
It was previously reported that ERK is activated within 30 min of injury and persisted for more than one week (Sheu et al., 2000; Harrisingh et al., 2004). Even though numer- ous ligands might be involved in the induction of this long lasting ERK activation, the initial activation of ERK after injury might be caused by the release of certain factors that are already present in the uninjured nerve. For example, NRG, which is derived from damaged axons and SCs, may be a promising candidate because NRG signaling is acutely activated after nerve injury, and has been implicated in the dedifferentiation of SCs via ERK activation (Harrisingh et al., 2004; Guertin et al., 2005). Later on, ERK activation may
be induced by injury-induced growth factors and cytokines.
For example, cytokines such as IL-6 could activate ERK in SCs as we have demonstrated in other studeies. The ERK activation by multiple ligands after nerve injury has been suggested to be associated with the initiation of de- myelination and proliferation (Harrisingh et al., 2004;
Monje et al., 2006), two important characteristics of dedif- ferentiation, of SCs. However, the molecular mechanism underlying the function of ERK in dedifferentiation of SCs remains largely uncharacterized. In this study, we have found that ERK activation induced by NRG and IL-6 is in- volved in the serine phosphorylation of the transcription factor STAT3 in SCs, thereby suggesting that ERK might play a role in the dedifferentiation of SCs, in part through the modulation of STAT3 activity.
Interestingly, our results suggested that the serine phos- phorylation of STAT3 induced by IL-6 may have a different meaning than that induced by NRG. IL-6 induced a dra- matic increase in the nuclear transport of double-phos- phorylated STAT3, whereas NRG induced only the serine phosphorylation of STAT3. Furthermore, the nuclear trans- port of serine phosphorylated STAT3 was not observed fol- lowing NRG treatment. It may be possible that serine phos- phorylated STAT3 by NRG may play a cytoplasmic func- tion, as was reported previously (Ng et al., 2006a) or NRG may induce genes with a very low level of serine phosphory- lated STAT3 in the nucleus (undetectable level with the immunostaining method). Recently, Ng et al (2006b) re- ported that nerve growth factor stimulates the expression of STAT3 target genes such as JunB and egr-1 by inducing solely via the induction of serine phosphorylation of STAT3 in PC12 pheochromocytoma cells. Further studies will be required to more clearly elucidate the functional significance of serine phosphorylated STAT3 in response to NRG in SCs.
On the other hand, it has been reported that the kinases that phosphorylate STAT3 on the serine 727 residue are diverse, and are dependent on both extracellular stimuli and cell types (Gartsbein et al., 2006; Ng et al., 2006b; Shi et al., 2006). However, we determined that the inhibition of MEK nearly completely inhibited the induction of serine phosphorylation of STAT3 in ex-vivo explant cultures. This finding demonstrates that ERK may constitute the princi- pal component of the serine/threonine kinase pathways that result in the phosphorylation of the serine 727 residue of STAT3 in SCs. Further studies will be required in order to elucidate the serine kinases that directly phosphorylate STAT3 in Schwann cells.
In conclusion, we have determined in this study that ERK is important in modulating the activity of STAT3 in cultured SCs and in injured nerves. Because ERK plays an important role in the dedifferentiation of SCs after nerve injury, our findings have important implications for our un- derstanding of the molecular mechanism of degenerative and regenerative processes occurring after nerve injury.
ACKNOWLEDGEMENTS
This work was supported by Research fund from Dong-A University.
REFERENCES
Battle TE, Frank DA. The role of STATs in apoptosis. Curr Mol
Med 2: 381−392, 2002.
Bleuel A, Monard D. Regulation of protease nexin-1 and angio- tensin II receptor subtype 1 expression: inverse relationship in experimental models of nerve injury. J Neurosci Res 42: 562−
570, 1995.
Cafferty WB, Gardiner NJ, Gavazzi I, Powell J, McMahon SB, Heath JK, Munson J, Cohen J, Thompson SW. Leukemia inhibitory factor determines the growth status of injured adult sensory neurons. J Neurosci 21: 7161−7170, 2001.
Chung J, Uchida E, Grammer TC, Blenis J. STAT3 serine phosphory- lation by ERK-dependent and -independent pathways negatively modulates its tyrosine phosphorylation. Mol Cell Biol 17: 6508−
6516, 1997.
Fu SY, Gordon T. The cellular and molecular basis of peripheral nerve regeneration. Mol Neurobiol 14: 67−116, 1997.
Gartsbein M, Alt A, Hashimoto K, Nakajima K, Kuroki T, Tennenbaum T. The role of protein kinase C delta activation and STAT3 Ser727 phosphorylation in insulin-induced kerati- nocyte proliferation. J Cell Sci 119: 470−481, 2006.
Guertin AD, Zhang DP, Mak KS, Alberta JA, Kim HA. Microa- natomy of axon/glial signaling during Wallerian degeneration.
J Neurosci 25: 3478−3487, 2005.
Hai M, Muja N, DeVries GH, Quarles RH, Patel PI. Comparative analysis of Schwann cell lines as model systems for myelin gene transcription studies. J Neurosci Res 69: 497−508, 2002.
Harrisingh MC, Perez-Nadales E, Parkinson DB, Malcolm DS, Mudge AW, Lloyd AC. The Ras/Raf/ERK signalling pathway drives Schwann cell dedifferentiation. EMBO J 23: 3061−3071, 2004.
Jain N, Zhang T, Fong SL, Lim CP, Cao X. Repression of Stat3 activity by activation of mitogen-activated protein kinase (MAPK).
Oncogene 17: 3157−3167, 1998.
Jessen KR, Mirsky R. Negative regulation of myelination: relevance for development, injury, and demyelinating disease. Glia 56:
1552−1565, 2008.
Jessen KR, Mirsky R. The origin and development of glial cells in peripheral nerves. Nat Rev Neurosci 6: 671−682, 2005.
Jirsova K, Sodaar P, Mandys V, Bar PR. Cold jet: a method to obtain pure Schwann cell cultures without the need for cytotoxic, apoptosis-inducing drug treatment. J Neurosci Methods 78: 133
−137, 1997.
Kamimura D, Ishihara K, Hirano T. IL-6 signal transduction and its physiological roles: the signal orchestration model. Rev Physiol Biochem Pharmacol 149: 1−38, 2003.
Lee HK, Seo IA, Park HK, Park YM, Ahn KJ, Yoo YH, Park HT.
Nidogen is a prosurvival and promigratory factor for adult
Schwann cells. J Neurochem 102: 686−698, 2007.
Lee HK, Seo IA, Suh DK, Hong JI, Yoo YH, Park HT. Interleukin-6 is required for the early induction of glial fibrillary acidic protein in Schwann cells during Wallerian degeneration. J Neurochem 108: 776−786, 2009.
Lee N, Neitzel KL, Devlin BK, MacLennan AJ. STAT3 phosphory- lation in injured axons before sensory and motor neuron nuclei:
potential role for STAT3 as a retrograde signaling transcription factor. J Comp Neurol 474: 535−545, 2004.
Monje PV, Bartlett Bunge M, Wood PM. Cyclic AMP synergistically enhances neuregulin-dependent ERK and Akt activation and cell cycle progression in Schwann cells. Glia 53: 649−659, 2006.
Ng DC, Lin BH, Lim CP, Huang G, Zhang T, Poli V, Cao X. Stat3 regulates microtubules by antagonizing the depolymerization activity of stathmin. J Cell Biol 172: 245−257, 2006a.
Ng YP, Cheung ZH, Ip NY. STAT3 as a downstream mediator of Trk signaling and functions. J Biol Chem 281: 15636−15644, 2006b.
Plaza-Menacho I, van der Sluis T, Hollema H, Gimm O, Buys CH, Magee AI, Isacke CM, Hofstra RM, Eggen BJ. Ras/ERK1/2- mediated STAT3 Ser727 phosphorylation by familial medullary thyroid carcinoma-associated RET mutants induces full activa- tion of STAT3 and is required for c-fos promoter activation, cell mitogenicity, and transformation. J Biol Chem 282: 6415−6424, 2007.
Qiu J, Cafferty WB, McMahon SB, Thompson SW. Conditioning injury-induced spinal axon regeneration requires signal trans- ducer and activator of transcription 3 activation. J Neurosci 25:
1645−1653, 2005.
Reich NC, Liu L. Tracking STAT nuclear traffic. Nat Rev Immunol 6: 602−612, 2006.
Sheu JY, Kulhanek DJ, Eckenstein FP. Differential patterns of ERK and STAT3 phosphorylation after sciatic nerve transection in the rat. Exp Neurol 166: 392−402, 2000.
Shi X, Zhang H, Paddon H, Lee G, Cao X, Pelech S. Phosphorylation of STAT3 serine-727 by cyclin-dependent kinase 1 is critical for nocodazole-induced mitotic arrest. Biochemistry 45: 5857−5867, 2006.
Thomson CE, Griffiths IR, McCulloch MC, Kyriakides E, Barrie JA, Montague P. In vitro studies of axonally-regulated Schwann cell genes during Wallerian degeneration. J Neurocytol 22, 590−602, 1993.
Wen Z, Zhong Z, Darnell JE Jr. Maximal activation of transcription by Stat1 and Stat3 requires both tyrosine and serine phosphory- lation. Cell 82: 241−250, 1995.