Nitrosopumilus spindle-shaped viruses (NSVs) infect ammonia- oxidizing archaea (AOA). In this study, we detected the presence of NSVs in different ecosystems using semi-nested PCR of the NSV-specific protein-primed DNA polymerase B (pPolB) gene.
From most samples (75%), NSV-like pPolB genes were amplified, indicating NSVs’ ubiquity in oceans, coastal waters, and terrestrial environments. Library analysis of the PCR amplicons showed low diversity of NSV-like pPolB genes in the ecosystems. Only 20 OTUs could be recovered, with 97% nucleotide similarity cutoffs and some dominant OTUs shared over different environments.
Collectively, our data indicate that NSVs might be widespread in environments where AOA inhabit.
Keywords: ammonia-oxidizing-archaea, distribution, nitrification, Nitrosopumilus spindle-shaped virus, protein-primed- DNA polymerase
Members of Thaumarchaeota have been reported to be widespread and are key ammonia-oxidizing organisms in the oceans, coastal waters, and terrestrial environments (Massana et al., 2000; Karner et al., 2001; Herndl et al., 2005; Wuchter et al., 2006; Hatzenpichler, 2012). In addition, because of the capability of autotrophic ammonia oxidation (Könneke et al., 2005, 2014; Walker et al., 2010), ammonia-oxidizing archaea
(AOA) play a key role in nitrogen and carbon cycles in different ecosystems. Thus, grasping the factors that regulate community composition and abundance of AOA is essential to under- standing nitrogen and carbon cycling in various ecosystems (Sintes et al., 2013; Santoro et al., 2017).
So far, more than 100 archaeal viruses have been identified.
However, slightly more than half of the archaeal viruses were studied in detail (Snyder et al., 2015), falling far behind their bacteriophage counterpart, which served for an extended period as a model for prokaryotic viral studies (Dennehy, 2009;
Pietilä et al., 2014; Turgeon et al., 2014; Emmoth et al., 2017).
The relative lack of archaeal virus studies was due to the complications accompanying culturing their archaeal hosts (Snyder et al., 2015) and viruses (Oren et al., 1997; Santoro et al., 2017). Hyperthermophilic viruses took the lead in the studies of archaeal viruses (Zillig et al., 1993, 1998; Prangishvili et al., 1999), followed by haloviruses from hypersaline habitat (Santos et al., 2012). Thus far, viruses that infect marine Thau- marchaeota have been recently reported throughout metagenomic and single-cell genome studies (Stahl and de la Torre, 2012;
Labonte et al., 2015; Roux et al., 2016; Ahlgren et al., 2017a, 2017b, 2019). A previous metagenomic-based study (López- Pérez et al., 2019) suggested that Thaumarchaeota viruses are diverse.
Isolation of Nitrosopumilus spindle-shaped viruses (NSVs)
Prevalence of Nitrosopumilus spindle-shaped viruses in different ecogeographical locations
Khaled S. Gazi 1 , Jong-Geol Kim 2 , Olorunleke Y. Olufowora 1 , Ui-Ju Lee 1 , and Sung-Keun Rhee 1 *
1
Department of Biological Sciences and Biotechnology, Chungbuk National University, Cheongju 28644, Republic of Korea
2
Department of Biological Science, Wonkwang University, Iksan 54538, Republic of Korea
Nitrosopumilus 감염 방추사 형태 바이러스의 생태계 분포
칼레드 사이드 가지 1 ・ 김종걸 2 ・ 올로론레케 잉카 올루포워라 1 ・ 이의주 1 ・ 이성근 1 *
1
충북대학교 생명시스템학과,
2원광대학교 생명과학과
(Received March 29, 2021; Revised April 21, 2021; Accepted April 22, 2021)
*For correspondence. E-mail: [email protected];
Tel.: +82-43-261-2300; Fax: +82-43-264-9600
that infect AOA members were succeeded for the first time (Kim et al., 2019), indicating viral predation could be a key factor affecting the activity and community composition of AOA in marine environments. NSVs comprise linear double- stranded (ds) DNA genomes of about 28 kb and represent a new virus family, Thaspiviridae. For replication of linear dsDNA chromosomes, viruses often employ a protein-primed DNA replication strategy in which protein-primed DNA polymerase functions. Kim et al. (2019) detected NSV-like protein primed
DNA polymerase B (pPolB) gene in marine sediment and seawaters using PCR primers targeting NSV-specific pPolB gene.
In this study, the biogeographical distribution and composition of NSV-like viral communities were investigated using PCR analysis of pPolB gene sequences of NSVs with degenerate primers newly designed.
To determine the biogeographical distribution of NSV-like viruses, 53 environmental samples from 16 different locations (Table 1) were collected: 1) seawater of west and east coast of
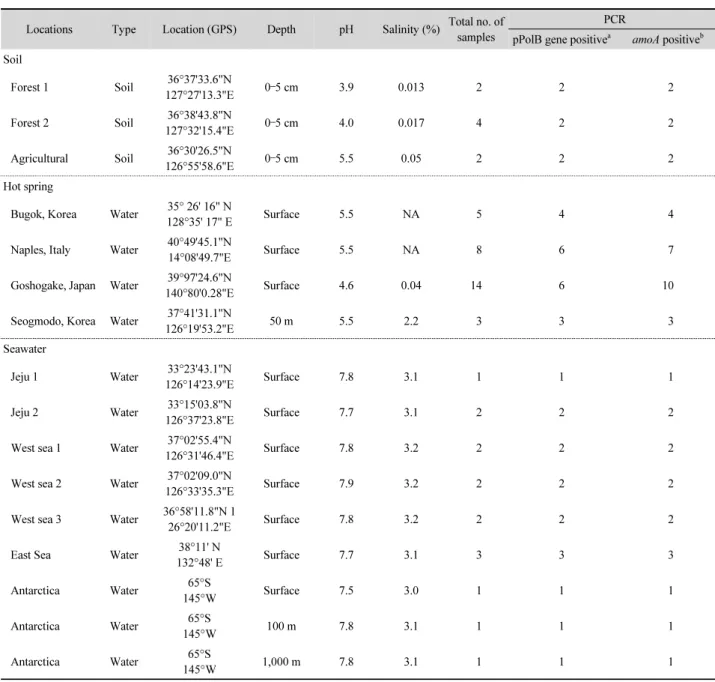
Table 1. Sample properties used in this study
Locations Type Location (GPS) Depth pH Salinity (%) Total no. of samples
PCR
pPolB gene positive
aamoA positive
bSoil
Forest 1 Soil 36°37'33.6"N
127°27'13.3"E 0–5 cm 3.9 0.013 2 2 2
Forest 2 Soil 36°38'43.8"N
127°32'15.4"E 0–5 cm 4.0 0.017 4 2 2
Agricultural Soil 36°30'26.5"N
126°55'58.6"E 0–5 cm 5.5 0.05 2 2 2
Hot spring
Bugok, Korea Water 35° 26' 16" N
128°35' 17" E Surface 5.5 NA 5 4 4
Naples, Italy Water 40°49'45.1"N
14°08'49.7"E Surface 5.5 NA 8 6 7
Goshogake, Japan Water 39°97'24.6"N
140°80'0.28"E Surface 4.6 0.04 14 6 10
Seogmodo, Korea Water 37°41'31.1"N
126°19'53.2"E 50 m 5.5 2.2 3 3 3
Seawater
Jeju 1 Water 33°23'43.1"N
126°14'23.9"E Surface 7.8 3.1 1 1 1
Jeju 2 Water 33°15'03.8"N
126°37'23.8"E Surface 7.7 3.1 2 2 2
West sea 1 Water 37°02'55.4"N
126°31'46.4"E Surface 7.8 3.2 2 2 2
West sea 2 Water 37°02'09.0"N
126°33'35.3"E Surface 7.9 3.2 2 2 2
West sea 3 Water 36°58'11.8"N 1
26°20'11.2"E Surface 7.8 3.2 2 2 2
East Sea Water 38°11' N
132°48' E Surface 7.7 3.1 3 3 3
Antarctica Water 65°S
145°W Surface 7.5 3.0 1 1 1
Antarctica Water 65°S
145°W 100 m 7.8 3.1 1 1 1
Antarctica Water 65°S
145°W 1,000 m 7.8 3.1 1 1 1
a
Number of samples with successful amplification of pPolB gene.
b
Number of samples with successful amplification of archaeal amoA gene.
South Korea, the coast of Jeju island, and Antarctic ocean, 2) terrestrial soils, and 3) hot spring waters from Seogmodu and Bugok in Korea, Naples in Italy, and Goshogake in Japan. Water and soil samples were kept in 4°C and -80°C respectively in the dark until DNA extraction. The pH of soil (soil/Milli-Q water ratio of 1:2.5) and water were measured with a pH analyzer (Mettler Toledo GmbH). Water samples were filtered using 0.2 μm pore size filters (45 mm diameter, Supor polyethersulfone;
Pall Life Sciences), and the FeCl 3 flocculation method was used for the concentration of viral particles from the filtrates (John et al., 2011). DNA was obtained using a DNeasy Blood and Tissue Kit (Qiagen). Soil samples were collected from the top 0-5 cm, then DNA was extracted from 0.5 g soil, using the Exgene Soil DNA mini kit (GeneAll), following the manu- facturer’s instructions.
DNA samples were checked for the presence of NSV-like virus using degenerated primers designed for the conserved regions in pPolB gene of NSVs (Kim et al., 2019). The primers were newly designed using pPolB genes from the three genomes of NSV isolates and two partial scaffolds (Fig. 2) while the previous primers of Kim et al. (2019) were designed using only viral genomes of NSV isolates. For the first step of the semi- nested PCR, primers, 620F (5'-GAAAATAYCARGAYAAR AAATTTCC-3') and 1880R (5'-TTTATGATCWCCATTRA GATCWATT-3'), were used. The PCR protocol is as follows:
95°C (5 min), then 25 cycles at 95°C (30 sec), 55°C (30 sec), and 72°C (1.5 min). Since the PCR amplification did not give any positive amplification for pPolB gene, another PCR primer, 1470F, was designed for a semi-nested PCR approach.
Semi-nested PCR approach was widely used in different scientific fields and showed high sensitivity toward bacteria, fungi, virus, and bacteriophage detection (Kembou Tsofack et al., 2017; Yang et al., 2017; John et al., 2019; Porter et al., 2020). Thus, for the second step of the semi-nested PCR, primers, 1470F (5'-ATGCWCYAAAGATAAMAAAGAAG AT-3') and 1880R (5'-TTTATGATCWCCATTRAGATCW ATT-3') were used. The PCR protocol was set up as follows:
95°C (5 min), then 25 cycles at 95°C (30 sec), 55°C (30 sec), and 72°C (1.5 min). Using the semi-nested PCR, most of the samples were PCR-positive for the NSV-like pPolB gene. In addition to the detection of NSV-like viruses in different coastal sea sediments around the Korean peninsula and Viet Nam (Kim
et al., 2019), the NSV-like pPolB gene has been successfully amplified from seawaters of the Antarctic ocean (100 m depth) in this study. Considering the widespread presence of AOA in terrestrial environments (Stopnišek et al., 2010; Gubry-Rangin et al., 2011) and hot springs (de la Torre et al., 2008; Lehtovirta- Morley et al., 2011, 2016; Hatzenpichler, 2012; Gubry-Rangin et al., 2017), amplification of NSV-like pPolB gene in these samples is not unexpected. However, NSVs are known to be specific to AOA group I.1a (Kim et al., 2019) although hot springs mostly contain thermophilic AOA groups (Abby et al., 2018).
For analyzing the community structure using pPolB gene libraries, paired-end Miseq Illumina platform reads (2 × 301 bp) were analyzed through QIIME2 (Bolyen et al., 2019).
pPolB gene sequences were clustered with a cutoff value of 97% identity at the nucleotide level to generate operational taxonomic units (OTUs) (Li et al., 2018), and chimeras were removed using de novo method to obtain final OTUs. pPolB gene OTUs were aligned and clustered in BioEdit software version 7.0.5.3 (Hall, 1999). Then, MEGA version 7 (Kumar et al., 2016) was used to construct the phylogenetic tree using the maximum-likelihood method and Kimura-2-parameter model with 1,000 bootstraps assessed for evolutionary distance. The maximum likelihood phylogenetic tree was created using PhyML pipeline (Guindon et al., 2010) with automatic selection of the best-fit substitution model for the provided alignment. The best model found by PhyML was VT + G + F. Midpoint view of the tree was rooted for better visualization. The branch support was calculated with aBayes applied in PhyML.
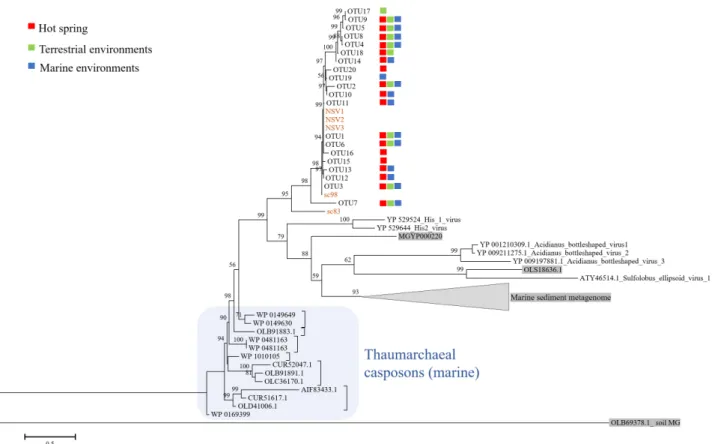
In this study, 20 OTUs were successfully obtained, and the OTU sequences were similar to each other and closely related to these previously obtained NSVs (Fig. 1). OTU1 was the most dominant (98.5% of the total reads) and recovered from all the samples (Fig. 1). Nine of the 20 OTUs were recovered from all three groups of samples from marine, terrestrial, and hot springs.
OTU19 and OTU17 were specific to the marine and terrestrial samples, respectively. Moreover, three OTUs (OTU15, OTU16, and OTU17) were obtained only from hot spring samples (Fig.
1). Recovering different OTUs from different habitats with
different salinity and pH indicates possible niche differentiation
of these viruses. Gaining a phenotype in favor of environmental
adaptation is common in viruses. Trade-offs were shown
previously with Φ6 phage, which gets adapted to heat shock but resulted in a low reproduction (Dessau et al., 2012), and foot- and-mouth disease virus resulted in smaller plaques; to withstand the low pH (Vázquez-Calvo et al., 2014). Besides, the limited diversity obtained in this study might also be attributable to the limited coverage of primers which were designed using only a few NSV isolates and scaffolds (Kim et al., 2019).
For comparison, host of NSV, AOA, was quantified using primers for archaeal amoA gene with a previous protocol (Park et al., 2008). Archaeal amoA gene could be detected in all the samples as pPolB gene. pPolB and amoA gene copies were quantified as described previously (Kim et al., 2019), using real-time PCR with CFX Connect Real-Time system (Bio-Rad Laboratories) using iQ SYBRGreen Supermix (Bio-Rad Labo- ratories) and built-in CFX manager software v3.0 (Bio-Rad Laboratories). AamoAF and AamoAR primers (Francis et al., 2005) were used with following PCR condition: 95°C (5 min), then 40 cycles at 95°C (30 sec), 55°C (30 sec), and 72°C (1.5
min). For quantification of viral pPolB gene, semi-nested real- time PCR was performed: the first PCR using primers 620F and 1880R with following PCR condition: first, 95°C (10 min), then 20 cycles at 95°C (30 sec), 55°C (30 sec), and 72°C (1.5 min); the second PCR using primers 620F and primer 950R (5'- CATGGCRTAWGGATAWGCWGAATT-3') with the same thermal program as the first step. In the semi-nested real-time PCR, all the PCR amplicons were purified and diluted (1:50) in 1× TE. Largely, the correlation of amoA and pPolB gene copies was not high. This might be caused by the different relative abundance of AOA clades in samples. It is not clear if NSVs infect only specific clades of AOA, such as Nitrosopumilus-like AOA. In that case, the abundance of NSV may not be correlated with the abundance of all AOA. In most of the samples, amoA gene copies were more abundant than NSV-like pPolB gene copies, while archaeal amoA and NSV-like pPolB gene copy numbers were similar in Jeju and the agricultural samples. In the case of large mismatches between pPolB and amoA copies,
Fig. 1. Phylogenetic tree of pPolB gene sequences. The OTUs obtained from different locations were indicated as follows red, green, and blue squares for hot
spring, terrestrial, and marine samples, respectively. Sequences from metagenomic datasets are indicated in gray. pPolB genes from isolated NSVs and
related scaffolds obtained in Kim et al. (2019) are indicated in brown. OTU sequences obtained in this study were deposited under the accession numbers
MW810454 to MW810473.
only limited members of AOA can be infected by NSVs since NSVs are known to have a very narrow host range (Kim et al., 2019). In hot spring samples of Japan and Italy, NSV-like pPolB gene copy numbers were close to the detection limit of the qPCR. In these cases, most AOA of thermophilic clades might have different cell surfaces and thus resistant to NSV- type viral infection. Interestingly, pPolB gene copies in the hot spring samples of Bugok were higher than amoA gene (Fig. 2).
Although NSV’s primary host was reported from the marine environment, NSV-like pPolB genes are distributed in different habitats around the globe. Detection of NSV-like pPolB genes in terrestrial environments can be explained by the presence of AOA group I.1a clades in terrestrial environments (Song et al., 2019). Besides, NSV-like viruses may often not be entirely marine AOA (group I.1a)-specific, and AOA of different taxonomic clades can be attacked by NSV-like viruses. Previous reports showed that bacteriophage could adapt to new hosts, which may play a critical role in supporting the distribution of a type of bacteriophage in different environments (Koskella and Meaden, 2013; Czajkowski et al., 2019). For example, bacteriophages (SN-1, SN-2, SN-X, and SN-T) were demonstrated to have the ability to infect S. natans and P. aeruginosa (Jensen et al., 1998).
DNA polymerase sequences of a lytic bacteriophage, T7-like Podophage, infecting E. coli, appear in aquatic, freshwater, terrestrial, and extreme environments (Breitbart et al., 2004).
Nearly identical cyanophages were also reported to be widely distributed in different environments (Short and Suttle, 2005).
Further, detection of NSV-like pPolB gene in hot springs might also be explained by the presence of NSV-like pPolB gene in other viruses with entirely different hosts such as non-AOA Thaumarchaeota or a relative of AOA, Aigarchaeota in hot springs.
In this study, NSV-like pPolB gene sequences were found to be prevalent in different environments where archaeal amoA was detected. The presence of identical NSV-like pPolB gene sequences in different ecosystems implies possible conservation of replication strategies of NSV-like viruses infecting AOA hosts of different clades. Cultivation of diverse NSV using novel AOA hosts is required to improve primers for NSV studies of diversity and abundance which could reveal virus and AOA interactions in ecosystems.
적 요
Thaspiviridae 과에 포함된 Nitrosopumilus 감염 방추사 형 태 바이러스(NSV)는 암모니아 산화 고균(ammonia oxidizing- archaea, AOA)을 감염시킨다. 본 연구에서는 NSV의 단백질 프라이머-의존 DNA 중합 효소 B (pPolB) 유전자에 특이적인 primer를 이용한 semi-nested-PCR 기법을 개발하였다. 대부분 의 환경(75%) 시료에서 NSV-like pPolB 유전자가 증폭됨을 통하여 NSV가 해양, 토양, 온천 생태계에 존재한다는 것을 확 인하였다. PCR 증폭된 유전자 라이브러리의 분석 결과 pPolB 유전적 다양성이 낮았으며, 97% 유사성 기준으로 오직 20개 의 OTU만이 존재하는 것을 보여주었다. 또한, 우점하는 몇 개 의 OTU는 상이한 환경에 공존하는 것을 알 수 있었다. 이 결과 는, AOA가 존재하는 다양한 환경에 NSV가 널리 퍼져 있음을 보여주고 있다.
Acknowledgments
This research was supported by Chungbuk National University Korea National University Development Project (2020).
Conflict of Interest
The authors have no conflict of interest related to this study.
Fig. 2. Quantification of copy numbers of archaeal amoA (empty bars) and
NSV pPolB (solid bars) gene measured by qPCR. Error bars represent
standard deviations for three replicates.
References