(genus Coreoleuciscus)
12
0
0
전체 글
(2) 2. 송하윤·김재훈·서인영·방인철. 수산자원 증식 등의 목적으로 국내에 도입되었으며, 우리나라. 잡종 개체의 유전형은 핵 유전자 분석을 통해 확인할 수 있. 담수생태계 정착에 성공하여 기존의 하천생태계에 많은 악영. 다 (Wendel and Doyle, 1998). 이 중 핵 DNA의 recombination. 향을 미치고 있다 (Byeon and Oh, 2015). 이에 따라 위의 두 종 은 환경부에서 생태계 교란 야생 동·식물로 지정하였으며,. activating gene 1 (RAG1) 유전자는 염기서열이 보존적이고. 진화를 잘 반영하는 것으로 알려져 (Slechtova et al., 2007;. 국외 도입종에 대한 연구는 다방면으로 진행되어 왔다 (Lee. Cebrat et al., 2008) 담수어류의 종 간 잡종 판별과 계통분류. al., 2012). 우리나라 토착 어종의 생존에 심각한 피해를 주는. 2007; Lee et al., 2009; Yoon et al., 2009; Kim et al., 2015) 담수. 국외 도입종에 대한 연구에 관심을 집중하는 사이에 국내에. 어류를 포함한 많은 어류 종의 염기서열 정보가 알려져 있다.. and Kim, 1987; Byeon et al., 1997; Lee et al., 2008; Song et. 한 수계에 분포하는 어종이 다른 수계로 옮겨지는 담수어류의 수계이탈 사례가 빈번하게 일어나고 있다 (Kim et al., 2006a,. 2006b; NFRDI, 2007; Byeon and Oh, 2015; Chae et al., 2015).. 국내 도입종의 확산은 교통과 운송수단의 발달로 매우 용 이해졌으며, 양식, 관상어의 방류, 지역주민과 환경단체에서 서식 어종의 증대를 위한 무분별한 방류활동 등 다양한 목적 에 의해 전국적으로 확산되고 있는 추세이다 (NFRDI, 2007; Byeon and Oh, 2015).. 에 효과적인 분자 마커로 사용하고 있으며 (Sonnenberg et al.,. 특히 모계유전을 하는 미토콘드리아 DNA의 cytochrome c. oxidase subunit 1 (CO1) 유전자는 담수어를 포함하는 척추동. 물의 분류군에서 서로 다른 보전적인 염기서열의 변이를 가지 고 있어 이를 이용하여 종을 구분하는 DNA barcoding 연구에. 이용하고 있으며 (April et al., 2011), 본 연구 대상종인 쉬리속. 어류의 종 판별에 효과적으로 접목된 바 있다 (Song and Bang, 2015).. 따라서 본 연구는 낙동강 지류인 황지천에 서식하는 쉬리속. 국내 도입종의 경우 명확한 이입시기와 방류목적이 불분명. 어류 개체군을 대상으로 유전자 분석과 지느러미 반문 비교를. 한 경우가 많고 이입된 집단들의 정확한 출처를 알 수 없으며,. 통해 쉬리속 어류 개체군의 종 동정과 잡종 개체군 검증 및. 이입종과 동일한 종 또는 근연종이 서식하고 있는 경우 이입. 분포에 대한 연구를 수행하였다.. 집단과 원집단 (native population)과의 교배로 인해 유전자 소. 멸 (genetic extinction) 현상 또는 유전자오염 (genetic pollution). 등의 문제가 발생된다 (Epifanio and Philipp, 2001; Kwon et al., 2014).. 국내의 경우 연곡천 (Kim et al., 2006a), 양양남대천 (Kim et. al., 2006b), 강릉남대천 (Byeon and Oh, 2015) 등 강원도 독립 수계에 서한아 수계 어류가 이입된 이후 정착에 성공하여 이 입종들의 개체군이 증가되고 있는 것이 확인되고 있으며, 도수. 터널을 통해 동진강으로 이입된 줄종개 (Cobitis tetralineata) 개체군으로 인해 동진강에 분포하는 점줄종개 (C. lutheri)의. 유전자가 줄종개의 유전자로 대체되는 현상이 발생하여 점줄 종개 집단의 유전자 소멸 (genomic extinction) 가능성에 대해 언급한 바 있다 (Kwon et al., 2014).. 본 연구대상 종인 쉬리속 (genus Coreoleuciscus) 어류는. Mori (1935)에 의해 처음 기재되었으며, Song and Bang (2015). 에 의해 섬진강과 낙동강 수계에 분포하는 쉬리 (C. splendidus). 집단이 외부형태와 유전적 차이를 근거로 참쉬리 (C. aeruginos) 로 분류되었다. 두 종의 분포는 백두대간 서쪽으로 흐르는 서 한아 수계 (west korea subdistrict)에 쉬리가 분포하며, 남한아 수계 (south korea subdistrict)는 참쉬리가 분포하여 두 종 간의. 분포 구계가 명확하게 구분된다 (Song and Bang, 2015). 그러 나 최근 낙동강 지류인 황지천에 쉬리 (C. splendidus type) 개. 체가 다수 발견되어 (Chae et al., 2015) 기존에 알려져 있던 두 종 간의 분포 구계와 차이가 있어 이들 개체군을 대상으로 외 부 형질과 유전자 분석을 통해 쉬리 (C. splendidus type)의 종 구분과 잡종 개체군 존재 여부에 대한 검증이 필요하였다.. 재료 및 방법 1. 실험어의 확보 및 표본제작. 본 연구를 위해 2012년 12월과 2015년 2월에 황지천 본류. 7개점을 대상으로 족대 (망목 4 × 4 mm)를 사용하여 쉬리속 어. 류를 채집하였다 (Table 1). 각 조사지점에서 채집된 개체들은. Table 1. Distribution on survey sites of the Hwang-ji stream in Nakdong River from 2012 to 2015 Site No. and Sampling locality. GPS coordinate. Site 1. Munbaek bridge Moongok-dong, Taebaek-city, Gangwon-do Site 2. Gaesan bridge Jangseong-dong, Taebaek-city, Gangwon-do Site 3, Taebaek palezoic museum Dongjum-dong, Taebaek-city, Gangwon-do Site 4, Sasirang-e village Gumunso-dong, Taebaek-city, Gangwon-do Site 5. Seokpo bridge Seokpo-3ri, Seokpo-myeon, Bonghwa-gun, Gyeongsangnam-do Site 6. Seonghwang bridge Seokpo-3ri, Seokpo-myeon, Bonghwa-gun, Gyeongsangnam-do Site 7. Gulhyeon bridge Seokpo-ri, Seokpo-myeon, Bonghwa-gun, Gyeongsangnam-do. 37°01ʹ14.60ʺN, 129°00ʹ01.00ʺE 37°05ʹ38.88ʺN, 129°02ʹ19.88ʺE 37°05ʹ23.02ʺN, 129°02ʹ40.08ʺE 37°05ʹ22.32ʺN, 129°02ʹ40.29ʺE 37°04ʹ01.83ʺN, 129°02ʹ56.44ʺE 37°03ʹ25.08ʺN, 129°03ʹ07.02ʺE 37°02ʹ34.02ʺN, 129°03ʹ43.40ʺE.
(3) 낙동강 상류 황지천에 서식하는 쉬리속 어류 집단의 종 동정 및 잡종 판별. 3. Table 2. Names and sequences information of degenerated primers used in this study Primer name CO1. RAG1. Sequence (5ʹ → 3ʹ). Total bases. CO1-5236f CO1-7217r CO1-inner. CCTCGATCCTACAAACTCT GCGTCTTGGAATCCTARTTG TCCAGCTATTTCTCARTATC. 19 20 20. RAG1-1480f RAG1-3067r. TTCCTSAGCTGCGYAAGTA TTGTGAGCYTCCATRAACT. 19 19. Song and Bang (2015)을 참고하여 지느러미 반문의 형태를 기. 준으로 쉬리 (C. splendidus type)와 참쉬리 (C. aeruginos type). 로 구분하여 현장에서 개체 수를 기록하였다. 채집 개체 수가 가장 많은 4번 조사지점 (강원도 태백시 구 문소동)에서 채집된 개체는 실험실로 운반하여 유전자분석과 지느러미 반문형태분석에 이용하였다. 10% 포르말린으로 고정 하여 표본을 제작하였고 각 개체마다 고유의 voucher number 를 부여하여 개체를 구분하였다. 2. Genomic DNA 추출 및 PCR. 실험어의 genomic DNA는 배지느러미 일부를 절단하여. TNES-Urea buffer (10 mM Tris-HCL pH 8.0; 125 mM NaCl;. 3730XL DNA analyzer)로 염기서열을 결정하였다. 3. 분자계통 분석 및 종 동정. 분자계통 분석은 확보된 RAG1 유전자 (1,334 bp)와 CO1 유. 전자 (1,551 bp)를 이용하여 2012년에 4번 조사지점 (강원도 태. 백시 구문소동)에서 채집된 36개체를 대상으로 종 및 잡종 개 체의 동정을 실시하였다. 대조군으로 쉬리 (voucher number-. SUC 2179, 강원도 원주시 섬강)와 참쉬리 (voucher number-. SUC 2281, 경상북도 봉화군 낙동강)를 포함하여 분자계통 분. 석을 실시하였다. Outgroup은 모래무지아과 어류인 줄몰개 (Gnathopogon strigatus, voucher number-SUC0857)와 참중. 고기 (Sarcocheilichthys variegatus wakiyae, voucher number-. 10 mM EDTA ph 8.0; 1% SDS; 8M urea)를 이용하여 추출하. SUC7109)를 사용하였다.. proteinase K (Sigma, USA)를 100 mg/mL의 농도로 첨가하여. 1999) 프로그램을 이용하여 multiple alignment를 실시하였으. chloroform : iso-amylalcohol, 25 : 24 : 1)을 처리하여 단백질을. lsenbeck and Ronquist, 2001) 프로그램을 사용하였다. MrBayes. 였다 (Asahida et al., 1996). 상기 시료를 포함하는 완충용액에. 55℃에서 12시간 동안 반응시켰다. 이후 PCI solution (phenol : 제거하였으며, 2-propanol로 DNA를 침전시켰다. 준비한 시료. 는 spectrophotometer 측정과 0.8% agarose gel 전기영동을 통 해서 DNA의 양과 질을 확인한 후 실험에 이용하였다.. 핵 DNA의 RAG1 유전자를 증폭하기 위하여 NCBI의 Gen. Bank로부터 잉어목 어류들의 염기서열을 확보한 후 ClustalW. 확보된 염기서열을 대상으로 BioEdit version 7.0.9 (Hall,. 며 (Thompson et al., 1997), 계통도 작성은 MrBayes 3.1.2 (Hue. 프로그램을 이용한 Bayesian 분석은 1 × 107 generations를 수. 행하였으며, 각 1000세대마다 무작위로 sample하였다. 또한. 분석 영역에서 수집된 sample은 100 generations 분석에서 제. 외하였고, 50%가 넘지 않는 분지도의 지지도는 표시하지 않 았다.. 분석을 통해 총 3개의 degenerate primer를 제작하였으며, 제. 작된 primer를 이용하여 총 1,334 bp를 증폭하였다. 미토콘. 드리아 DNA의 CO1 유전자 역시 위와 같은 방법으로 2개의. primer를 제작하여 1,551 bp를 증폭하였다 (Table 2).. PCR 반응은 100 ng의 주형 DNA와 10 pmol의 primer를 사. 용하여 PCR premix 20 μL (Bioneer, Korea)로 반응시켰다. PCR. 조건은 95℃에서 4분간 initial activation시킨 후 95℃에서 1. 분간 denaturation, 53~55℃에서 1분간 annealing, 72℃ 1분. 간 extension 반응을 34회 반복하였다. 증폭된 산물은 1.5%. agarose gel 상에서 전기영동한 후에 ethidium-bromide (Et-Br) 용액으로 염색을 하여 band의 유무를 확인하였다. 효과적으. 로 증폭된 산물은 PCR Purification kit (Bioneer, Korea)로 정 제한 후, 동일한 PCR primer를 사용하여 염기서열분석기 (ABI. 4. 잡종 개체군 모계추정. 자연 개체군 내 잡종 개체의 비율과 모계추정은 4번 조사지. 점 (강원도 태백시 구문소동)에서 채집된 105마리를 대상으로. RAG1 유전자의 sequence chromatogram에 나타나는 double peaks의 유무를 통해 잡종 개체를 판별하였고 CO1 유전자의. 염기서열 분석을 통해 잡종 개체의 모계를 추정하였다. 5. 지느러미 반문비교. 4번 조사지점 (강원도 태백시 구문소동)에서 채집한 개체의 염기서열 분석결과를 기반으로 쉬리, 참쉬리 및 잡종 개체를 구분한 후 개체별로 순종표본인 쉬리 (SUC1313-1333, 경기도.
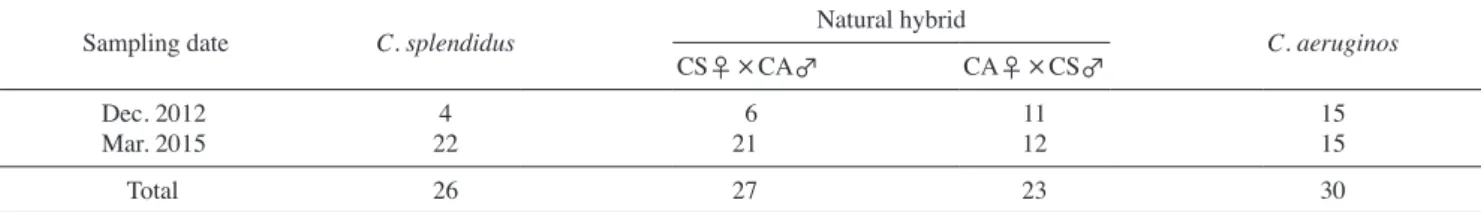
(4) 4. 송하윤·김재훈·서인영·방인철. Table 3. Number of individuals of Coreoleuciscus species collected from Hwang-ji Stream (2012~2015). This table is the result of morphological classification between C. splendidus and C. aeruginos, except for the genetic information Site numbers and sampling locality Site 1. Munbaek bridge Moongok-dong, Taebaek-City, Gangwon-do Site 2. Gaesan bridge Jangseong-dong, Taebaek-city, Gangwon-do Site 3, Taebaek palezoic museum Dongjum-dong, Taebaek-city, Gangwon-do Site 4, Sasirang-e village Gumunso-dong, Taebaek-city, Gangwon-do Site 5. Seokpo bridge Seokpo-3ri, Seokpo-myeon, Bonghwa-gun. Gyeongsangnam-do Site 6. Seonghwang bridge Seokpo-3ri, Seokpo-myeon, Bonghwa-gun. Gyeongsangnam-do Site 7. Gulhyeon bridge Seokpo-ri, Seokpo-myeon, Bonghwa-gun. Gyeongsangnam-do Total. 2012 Year C. splendidus type. 2015 Year. C. aeruginos type. C. splendidus type. C. aeruginos type. 2. 7. 3. 13. 2. 10. 3. 15. 13. 10. 11. 19. 5. 31. 32. 38. 12. 17. 12. 10. 0. 17. 27. 22. 0. 18. 0. 25. 34. 110. 88. 142. 가평군 조종천)와 참쉬리 (SUC 1290-1390, 경상북도 산청군 덕천강) 표본과 비교하였다. 쉬리와 참쉬리의 대표적인 형질과 잡종 개체의 지느러미 반문변이는 사진을 촬영하여 개체 간. 되어 총 17마리 (48%)의 잡종 개체를 확인하였다 (Fig. 1). 한편 순종 참쉬리는 15개체로 확인되었고 나머지 4개체 (SUC7841-. 7844)는 외형과 유전자가 순종 쉬리와 동일하였다 (Figs. 1, 2,. 4).. 비교하였다.. 두 종 간 잡종 개체의 유전형과 개체군 내 비율을 알아보고 자 가장 많은 개체가 채집된 4번 조사지점 (강원도 태백시 구. 결. 과. 문소동)에서 채집된 105마리를 대상으로 RAG1 유전자의 염 기서열을 분석한 결과 1,334 bp의 염기서열 중 13개 위치에서. 낙동강 상류 지류인 황지천의 7개 정점 (Table 1)을 대상으. 변이가 관찰되어 변이율은 약 0.95%였다. 또한 잡종 개체들은. 포현황을 조사하였다. 2012년 조사결과 쉬리 (C. splendidus. 위치에서 두 종의 염기서열이 함께 표현되는 double peaks가. 로 2012년 12월과 2015년 2월에 각 1회씩 쉬리속 어류의 분. type)는 5개 조사지역 (site 1~5)에서 서식이 확인되어 총 34개. 쉬리와 참쉬리의 두 종 간 염기서열 변이를 보인 9개의 변이. 나타났고 (Figs. 2, 3) 나머지 4개의 염기서열 변이는 개체 변이. 체를 채집하였고, 참쉬리 (C. aeruginos type)는 모든 조사지점. 에 의한 haplotype이었다.. 년 조사결과에서 쉬리 (C. splendidus type)는 6개 조사지점 (site. 쉬리와 참쉬리를 모계로 하는 비율에 큰 차이가 없어 (Table 4). 에서 서식이 확인되어 110개체가 채집되었다 (Table 3). 2015 1~6)에서 확인되어 서식 분포가 하류로 확산되었으며, 88개. 체가 채집되어 총 122개체의 쉬리 (C. splendidus type)가 채집. 되었다 (Table 3).. 확보된 염기서열을 이용하여 개체군의 분자계통과 종 동 정을 실시한 결과 RAG1 유전자와 (1,334 bp) CO1 유전자의. (1,551 bp) 계통도 상에서 쉬리와 참쉬리 두 종과 잡종 개체가 명확하게 구분되었다 (Fig. 1). 또한 분석에 이용한 36개체 중. 6개체는 (SUC7845, 7854, 7866 7852, 7856, 8107) 모계가 쉬리. 인 잡종 개체로 확인되었으며, 11개체 (SUC7846-7851, 7853,. 7867, 7868, 8103, 8106)는 모계가 참쉬리인 잡종 개체로 확인. CO1 유전자를 이용하여 잡종 개체의 모계를 추정한 결과. 특정 종에 대해 모계가 편중되지 않았으며, 순종의 정교배체 (C. splendidus ♀ × C. splendidus ♂, C. aeruginos ♀ × C. aeru. ginos ♂)와 잡종의 상반교배체 (C. splendidus ♀ × C. aeruginos ♂, C. aeruginos ♀ × C. splendidus ♂)가 계통도 (Fig. 1)와 염 기서열 분석결과 (Fig. 4)에서 명확히 구분되었다.. 염기서열 분석결과를 기준으로 지느러미 반문의 형태를 각. 종의 대조군과 비교한 결과 쉬리와 참쉬리 개체는 각 종의 대 조군과 반문 분포와 형태가 동일하였으나, 잡종 개체는 등지 느러미와 꼬리지느러미의 반문이 참쉬리의 반문 모양에 쉬리 의 반문 특징이 일부 반영된 중간 형질로 나타났다. 특히 잡종.
(5) 낙동강 상류 황지천에 서식하는 쉬리속 어류 집단의 종 동정 및 잡종 판별. 5. Fig. 1. Phylogenetic trees inferred from the nuclear recombination activating gene 1 (RAG1) gene, and mitochondrial cytochrome c oxidase subunit 1 (CO1) gene of a natural hybrid and two Coreolcuciscus species (collected at 2012). Numbers on the trees are posterior probability values from Bayesian inference (BI) analysis. Hybrid individuals in this study are in the blue and the red background font. (The blue background is C. splendidus × C. aeruginos hybrid, and red background is C. aeruginos × C. splendidus hybrid, respectively).. 개체는 뒷지느러미 기저부에 위치한 검은색 반문이 전혀 발현 되지 않아 순종 쉬리와 구분되었다 (Fig. 5).. korea subdistrict)에 속하고 있으며 (Kim et al., 2005), 얼룩새. 코미꾸리 (Koreocobitis naktongensis), 수수미꾸리 (Niwaella. multifasciata), 여울마자 (Microphysogobio rapidus), 꼬치동자 개 (Pseudobagrus brevicorpus)와 같이 낙동강에 제한적인 분. 고. 찰. 포를 가지는 우리나라 고유종의 서식 빈도가 높게 나타난다. 그러나 현재 낙동강에는 우리나라의 다른 하천들에 비해 외래. 낙동강 상류에 서식하는 어류는 14과 34속 42종이 (Chae et. 종 및 다른 수계로부터 어류의 도입이 빈번하게 일어나고 있. 류 하천에 폭 넓게 분포하고 있다 (Song and Bang, 2015). 낙. 한 연구자료가 부족하며 이로 인한 어류상 및 수생태계의 변. al., 2015) 분포하고 있으며, 이 중 참쉬리는 낙동강의 각 지. 동강은 담수어류의 지리적 분포 구계에서 남한아 지역 (south. 으나 (Chae, 2003) 이들 종들에 대한 유입 경로와 확산 등에 대 화에 대한 포괄적인 연구는 미흡한 실정이다 (Jang et al., 2001;.
(6) 6. 송하윤·김재훈·서인영·방인철. Fig. 2. Thirteen base positions in which sequences of recombination activating gene 1 (RAG1) gene showing natural hybrid individual and its supposed parent species, C. splendidus and C. aeruginos. The polymorphic base positions are indicated according to IUPAC codes.. Chae, 2003; Chae et al., 2015).. 현재 낙동강에는 배스, 블루길 등의 외래종뿐만 아니라 Chae. et al. (2015)의 연구결과에 따르면 어름치 (Hemibarbus mylo. don), 참종개 (Iksookimia koreensis), 새코미꾸리 (K. rotundicau. data) 등 9종의 어류가 한강수계에서 낙동강으로 이입되었음. 을 확인하였고, 쉬리 또한 2014년에 한강 type으로 여겨지는. 개체군이 낙동강 지류인 황지천과 경북 봉화군 석포면 낙동강 본류에서 채집되어 낙동강에 서식함을 보고하였다. 본 연구에서 2012년부터 2015년까지 쉬리속 어류의 분포를. 2회 조사한 결과 쉬리가 6개 조사지점에서 88개체가 채집되. 어 쉬리 개체군이 강원도 태백시에서 경상북도 봉화군 석포면 에 이르는 황지천 전 지역에 분포하고 있었다. 2012년 1차 조.
(7) 낙동강 상류 황지천에 서식하는 쉬리속 어류 집단의 종 동정 및 잡종 판별. 7. Fig. 3. Samples were obtained from pure species C. splendidus and C. aeruginos, as well as a suspected hybrid individual. The sequence traces show diagnostic positions for the two species and the double peaks at the respective positions for the apparent hybrid individuals.. 사에서 쉬리가 5번 조사지점까지 서식이 확인되어 분포가 황. 지천에 한정되었으나, 이후 2차 조사와 Chae et al. (2014)의 연 구결과를 통해 황지천뿐만 아니라 낙동강 본류 (경상북도 봉화. 군 석포면)까지 쉬리의 서식 분포가 확인되었다.. 소 등의 2차적인 피해를 야기하지 않을 것으로 판단되었으나 이입된 쉬리 개체군으로 인해 참쉬리 개체군과 교배를 통한 잡종 개체군이 발생됨에 따라 참쉬리 집단의 유전자 풀 (gene. pool)에 변화가 발생하고 있음을 확인하였다.. 따라서 강원도 태백시의 황지천 중류지역에서 경상북도 봉. 최근 근연종의 이입으로 인해 유전자 풀에 변화가 진행 중. 화군 석포면의 낙동강 상류 일부 지역에 치어부터 성어까지. 인 동진강에 서식하는 점줄종개 집단은 1931년과 1945년에. 모든 연령대의 쉬리 개체군이 채집됨에 따라 2012년 이전에 쉬리가 낙동강 상류지역에 이입되어 개체군이 낙동강 본류로 확산되며 안정적으로 서식하고 있음이 확인되었다.. 건설된 도수터널로 인해 섬진강에 분포하는 줄종개 개체군이 과거 80년 동안 지속적으로 이입됨에 따라 두 종 간의 잡종 개. 체군이 형성되었으며 (Kim and Lee, 1984; Kwon et al., 2014),. 황지천을 비롯한 낙동강 본류와 지류 하천의 참쉬리 집단은. 잡종 개체군의 세대가 진행될수록 잡종 개체군 유전자가 줄종. 신종으로 분류되기 이전부터 서한아 수계에 분포하는 쉬리 집. 개의 유전자로 변화되는 유전자 교체 (genomic replacement). 단과 지느러미 반문의 차이와 더불어 분자계통학적으로 두 종. 현상으로 인해 토착종인 점줄종개의 유전자가 사라질 가능성. 으로 뚜렷하게 구분됨이 보고되어 왔다 (Song and Park, 2006;. 이 높을 것으로 예측한 바 있다 (Kwan et al., 2014).. Song and Bang, 2009; Song et al., 2010). 낙동강에 이입된 쉬. 리의 경우 끄리 (Opsariichthys uncirostris)의 이입사례 (Chae, 2003)와 같이 강한 어식성으로 인해 토착 어종의 개체 수 감. 낙동강 상류 지역에 인위적으로 어류가 이입되는 요인으로 Chae et al. (2015)는 1990년 이후 여러 차례 석포 제련소에서 제조된 황산이 운반 도중 낙동강의 지류나 본류의 황산이 누.
(8) 8. 송하윤·김재훈·서인영·방인철. Fig. 4. Nucleotide sequence alignment of the cytochrome c oxidase subunit 1 gene (CO1) of a natural hybrid individual and its supposed parent species, C. splendidus and C. aeruginos. The polymorphic base positions are indicated according to IUPAC codes. *CS, C. splendidus; CA, C. aeruginos.. 출됨에 따라 대량의 어류 폐사 사고가 발생하였으며, 지역주. 한강에서 낙동강으로 이입되었음을 확인하였다.. 민들이 어족자원의 빠른 복원을 위해 한강수계 지역에서 어류. 따라서 도수터널을 통해 지속적으로 줄종개 개체군이 이입. 를 구매하여 방류함에 따라 쉬리를 포함한 5종 이상의 어류가. 된 점줄종개 개체군과는 달리 황지천에는 인위적인 영향으로.
(9) 낙동강 상류 황지천에 서식하는 쉬리속 어류 집단의 종 동정 및 잡종 판별. 9. Table 4. Species identification and number of individuals of two Coreoleuciscus species by mtDNA and nrDNA analysis (collected in the site no. 4, from 2012 to 2015) Sampling date. C. splendidus. Dec. 2012 Mar. 2015. 4 22. Total. 26. Natural hybrid. C. aeruginos. CS♀ × CA♂. CA♀ × CS♂ 11 12. 15 15. 27. 23. 30. 6 21. Fig. 5. Coreoleuciscus species and hybrid type from the Hwang-ji stream (Nakdong River) showing external morphological differences. They have differences in black array(s) on dorsal, caudal, and anal fin rays. Photographs show the morphological traits of the pure C. splendidus (A, B) and C. aeruginos (C, D), and hybrid individuals (Hybrid type 1~5). Red arrows are morphological variation points.. 비교적 소수의 쉬리 개체군이 이입되었을 것으로 추정되므로 동진강의 점줄종개 집단과 같은 급속적인 유전자 대체현상 및 낙동강에 분포하는 참쉬리 집단의 유전자 소멸까지는 나타나 지 않을 것으로 판단되었다. RAG1 유전자 염기서열과 CO1 유전자의 염기서열을 이용. 한 계통도 분석 결과 기존의 연구결과 (Song et al., 2010; Song. 류는 속 간에도 잡종 개체가 빈번하게 발생하고 있다 (Kuriiwa. et al., 2007).. 쉬리속 어류의 산란생태는 수심 10~20 cm, 유속 5~15 cm/. sec인 하상이 돌로 이루어진 여울에 집단적으로 산란하는 것. 으로 알려져 있으므로 (Song, 1976; Song and Kwon, 1993) 계 통도 상에서 잡종 개체의 모계가 특정 종에 편중되지 않는 결. and Bang, 2015)와 동일하게 쉬리와 참쉬리 두 종이 계통도. 과는 집단 산란하는 쉬리속 어류의 산란생태가 반영된 결과로. 상에서 잘 구분됨에 따라 잡종 개체 또한 명확하게 구분이 되. 판단된다.. 었으며 (Fig. 1), 잡종 개체는 CO1 유전자의 모든 염기서열이. RAG1 유전자의 염기서열 분석 결과에서 잡종 개체는 쉬리. 모계에 따라 100% 일치하였다 (Fig. 4). 이는 모계 유전하는 미. 와 참쉬리의 종 간 변이서열 위치에서 두 종의 염기서열이 함. 토콘드리아 DNA의 특성으로 (Hauswith and Clayton, 1985) 본. 께 표현되는 double peaks가 확인되어 (Figs. 2, 3) 모계와 부. 연구 결과에서도 잡종 개체는 모계와 동일한 미토콘드리아 유. 계의 변이 영역을 반영하고 있어 멘델의 유전법칙을 잘 반영. 전자 정보를 가지는 일반적인 특징을 보였다.. 하였다. 또한 sequence electropherogram에 나타난 sequence. 어류는 자연 상태에서도 산란시기 중복과 산란 장소의 부. 의 peaks가 잡종 F1 세대에서 나타나는 동일한 비율의 double. 족현상 (Smith et al., 2004; Kim et al., 2010, 2014), sneaker 행. peaks로 분석되어 (Sonnenberg et al., 2007; Kim et al., 2015). 2002)등으로 인해 종 간 잡종화 (hybridization)가 발생하며, 돌. 3).. 동 (Jansson et al., 1991; Kanoh, 2000; Candolin and Reynolds, 고기 (Puntungia herzi)와 참붕어 (Pseudorabora parva)의 속. 간 자연 잡종 개체가 발견된 경우와 같이 (Kim et al., 2015) 어. 본 연구에서 분석한 잡종 개체는 F1 세대로 추정되었다 (Fig. 쉬리와 참쉬리는 혼인색과 등지느러미, 뒷지느러미, 꼬리. 지느러미에 분포하는 검은색 반문이 종 간 차이를 보이며, 특.
(10) 10. 송하윤·김재훈·서인영·방인철. 히 뒷지느러미 기저부에 위치한 반점의 유무에 따라 두 종을. 를 이용한 분자계통 분석결과 황치천에 분포하는 쉬리속 어. 명확하게 구분할 수 있다 (Song and Bang, 2010; Song et al.,. 류는 쉬리, 참쉬리 두 종과 두 종 간의 잡종 개체군으로 구성. 2010).. 되어 있음을 확인하였으며, CO1 유전자의 염기서열 분석결과. 황지천에서 채집된 개체 또한 지느러미 반문을 기준으로 쉬. 순종인 정교배체와 잡종인 상반교배체가 잘 구분되었다. 또한. 리와 참쉬리로 분류가 가능하였고, 잡종 개체의 경우 등지느. RAG1 유전자 분석결과 13개의 염기서열 변이를 확인하였고,. 러미, 꼬리지느러미, 뒷지느러미의 반문 모양과 분포가 쉬리 와 참쉬리 두 종의 특징이 모두 반영되어 (Fig. 5) 어류에서 잡 종 자손은 지느러미 반문, 체색, 무늬 등 외부 형질에서 양친 의 중간 형질을 나타내는 결과와 (Chevassus, 1983; Kim et al,. 2014) 동일하였다. 특히 본 연구에서 분석한 잡종 개체는 뒷지. 잡종 개체는 9개의 염기서열에서 double peaks가 확인되었다. 유전학적 분석과 외부형태 변이 분석에 의해 쉬리와 참쉬리 사이에 잡종화가 발생한 것을 확인하였으나 잡종 F2세대와. 잡종 F1 세대의 생식적 격리 여부는 확인하지 못하였다.. 느러미 기저부에 위치한 반점이 전혀 표현되지 않아 잡종 개 체와 순종 쉬리를 구분하는 외형적인 특징으로 사용할 수 있 을 것으로 판단되었다. 따라서 본 연구에서 분석한 쉬리와 참쉬리 및 잡종 개체의 유전적 특성과 형태적 특징은 종 및 잡종 개체를 분석한 이 전의 연구결과들과 동일하여 (Sonnenberg et al., 2007; Lee et. al., 2009; Yoon et al., 2009; Kim et al., 2014; Song and Bang, 2015) 낙동강 지류인 황지천에는 인위적인 이입에 의해 쉬리. 와 참쉬리 순종이 함께 분포하며 두 종 간의 2차 접촉으로 잡 종 개체군이 발생하여 서식하고 있음을 확인하였다. 하지만 이 들 잡종 개체들이 번식에 참여하여 발생된 잡종 F2 세대는 본. 연구결과에서 명확하게 확인하지 못하여 추후 microsatellite. 마커를 이용하여 잡종 개체군에 (F1, F2 and backcross) 대한 유전형 분석과 함께 생식소 발달상태 분석을 통한 생식적 격 리 여부에 대한 후속연구가 필요하였다. 현재 낙동강에는 한강에서 이입된 참종개, 새코미꾸리 두 종. 과 근연종인 기름종개 (C. hankugensis) 얼룩새코미꾸리가 분포 하고 있으며, 실제 황지천에서 참종개와 기름종개의 잡종 개 체로 여겨지는 개체가 채집된 바 있고 (Chae et al., 2014) 미꾸 리과 어류에서는 종 간 잡종화 현상이 다수 보고되어 있기 때 문에 (Choi et al., 1995; Ko, 2009; Lee et al., 2009; Kwon et al.,. 2014) 쉬리속 어류뿐만 아니라 인위적 이입에 의한 종 간 잡 종 개체군의 존재 여부에 대한 후속 검증연구가 요구되었다.. 요. 약. 낙동강 상류 지류인 황지천에서 쉬리 (Coreoleuciscus splen. didus)와 참쉬리 (C. aeruginosa)의 종 간 자연잡종 개체를 채. 집하였다. 쉬리와 참쉬리의 종 간 잡종 개체는 외부형태 비교 와 함께 핵 DNA의 RAG1 유전자 (1,334 bp)와 미토콘드리아. DNA인 CO1 유전자 (1,551 bp)를 이용한 염기서열 분석을 실 시하였다. 외부형태 분석결과 잡종 개체는 등지느러미, 꼬리 지느러미 및 뒷지느러미 3곳에서 지느러미 반문의 형태가 쉬 리와 참쉬리의 중간 형태를 나타내었다. RAG1과 CO1 유전자. REFERENCES April, J., R.L. Mayden, R.H. Hanner and L. Bernatchez. 2011. Genetic calibration of species diversity among North america’s freshwater fishes. Nat. Acad. Sci., 108: 10602-10607. Asahida, T., T. Kobayashi, K. Saitoh and I. Nakayama. 1996. Tissue preservation and total DNA extraction from fish stored at ambient temperature using buffers containing high concentration of urea. Fish. Sci., 62: 727-730. Byeon, H.K. and J.K. Oh. 2015. Fluctuation of fish community and inhabiting status of introduced fish in Gangeumgnamdae stream, Korea. Korean J. Environ Ecol., 29: 718-728. (in Korean) Candolin, U. and J.D. Reynolds. 2002. Why do males tolerate sneakers? Tests with the European bitterling, Rhodeus sericeus. Behav. Ecol. Sociobiol., 51: 146-152. Cebrat, A., A. Cebula, A. Laszkiewicz, M. Kasztura, A. Miazek and P. Kisielow. 2008. Mechanism of lymphocyte-specific inactivation of RAG-2 intragenic promoter of NWC: Implications for epigenetic control of RAG locus. Mol. Immunol., 45: 2297-2306. Chae, B.S. 2003. Introduction of native fish species to the other river system. Case study. In: The introduced fishes of Korea. The Ichthyological Society, pp. 65-86. (in Korean) Chae, B.S., S.K. Kim, Y.H. Kang, N.S. Heo, J.M. Park, H.U. Ha and U.W. Hwang. 2015. Ichthyofauna and fish community structure in upper reach of the Nakdong River, Korea. Korean J. Ichthyol., 27: 116-132. (in Korean) Chevassus, B. 1983. Hybridization in fish. Aquaculture, 33: 245262. Choi, C.G., Y.J. Hwang, M.S. and M.S. Na. 1995. A natural hybrid between spinous laoch, Cobitis longicorpus and cyprinid loach Misgurunus anguilicaudatus (Pices, Cobitis). Korean J. Ichthyol., 7: 203-207. (in Korean) Epifanio, J. and D. Philipp. 2001. Simulating the extinction of parental lineages from introgressive hybridization: the effects of fitness, initial proportions of parental taxa, and mate choice. Rev. Fish Biol. Fisheries, 10: 339-354. Hall, T.A. 1999. BioEdit: A user-friendly biogical sequence alig-.
(11) 낙동강 상류 황지천에 서식하는 쉬리속 어류 집단의 종 동정 및 잡종 판별. ment editor and analysis program for windows 95/98/NT. Nucl. Acids. Symp. Set., 41: 95-98. Hauswith, W.W. and D.A. Clayton. 1985. Length heterogeneity of a conserved displacement-loop sequence in human mitochondrial DNA. Nucl. Acids Res., 13: 8093-8104. Huelsenbeck, J.P. and F.R. Ronquise. 2001. MRBYES: bayesian inference of phylogeny. Bio-informatics, 17: 754-755. Jang, M.H., G.I. Cho and G.J. Joo. 2001. Fish fauna of the main channel in the Nakdong river. Korean J. Limnol., 34: 223238. (in Korean) Jansson, H., U. Holmgrea, K. Wedin and T. Andersson. 1991. High frequency of natural hybrids between trout, S. trutta L., in a Swedish river. J. Fish Biol., 39: 343-348. Kanoh, Y. 2000. Reproductive success associated with territoriality, sneaking, and grouping in male rose bitterings, Rhodeus ocellatus (Pisces: Cyprinidae). Environ. Biol. Fishes, 57: 143-154. Kim, C.H., K.E. Hong, J.H. Kim and K.H. Kim. 2006a. Ichthyofauna in Yeongok stream, ganneung, Korea. Korean J. Ichthyol., 18: 244-250. (in Korean) Kim, C.H., W.O. Lee, K.E. Hong, C.H. Lee and J.H. Kim. 2006b. Icthyofauna and fish community structure in Namdae stream, Yangyang, Korea. Koran J. Ichthyol., 18: 112-118. (in Kor ean) Kim, C.H., W.O. Lee, Y.J. Kang and J.M. Baek. 2010. Occurrence of a natural intergeneric hybrid, Rhodeus uyekii × Acheilognathus signifer (Pisces: Cyprinidae) from Jojongcheon Bukhan River, Korea. Korean J. Ichthyol., 22: 225-229. (in Korean) Kim, H.S., S.W. Yun, J.G. Ko and J.Y. Park. 2014. Occurrence of a natural intergeneric hybrid between Rhodeus pseudosericeus and Acheilognathus signifer (Pisces: Cyprinidae) from the Namhangang (river), Korea. Koran J. Ichthyol., 26: 153-158. Kim, I.S. 1997. Illustrated encycolopedia of fauna & flora of Korea, Vol. 37, Freshwater fishes. Ministry of Education, Yeongi, pp. 216-271. (in Korean) Kim, I.S. and W.O. Lee. 1984. Effects of stream modification in the Seomjin river on the fish communities of the Dongjin river in Korea. Bull. Korean Fish. Soc., 17: 549-556. (in Korean) Kim, I.S., Y. Choi, C.L. Lee, Y.J. Lee, B.J. Kim and J.H. Kim. 2005. Illustrated book of Korean fishes. Kyohak Publishing Co. Ltd., Seoul, 615pp. (in Korean) Kim, K.S., H.R. Lee, A. Sade and I.C. Bang. 2014. Identification of hybrid between the tiger grouper Epinephelus fuscoguttatus and the giant grouper E. lanceolatus by analyzing COX I and RAG 2 sequences. Korean J. Ichthyol., 26: 70-73. (in Korean) Kim, K.Y., M.H. Ko, S.J. Cho, W.J. Kin, M.H. Son and I.C. Bang. 2015. A natural hybrid of intergeneric mating between a female Puntungia herzi and a male Pseudorasbora parva (Cypriniformes: Cyprinidae). Fish. Aquat. Sci., 18: 99-107. Kim, M.J. and C.B. Song. 2011. Origin of the Korean mandarin fish, Siniperca scherzeri and molecular phylogenetic rela-. 11. tionships to other Siniperca fishes. Koran J. Ichthyol., 23: 95-105. (in Korean) Ko, M.H. 2009. Reproductive mechanisms of the unisexual diploidtriploid hybrid complex of between the spine loach Cobitis hankugensis and Iksookimia longicorpa (Teleostei, Cobitis) in Korea. Doctors’s thesis, Chonbuk National University, 160pp. (in Korean) Kuriiwa, K., N. Hanzawa, T. Yoshino, S. Kimura and M. Nishida. 2007. Phylogenetic relationships and natural hybridization in rabbitfishes (Teleostei: Siganidae) inferred from mitochondrial and nuclear DNA analysis. Mol. Phyl. Ecol., 45: 69-80. Kwan, Y.S., M.H. Ko and Y.J. Won. 2014. Genomic replacement of native Cobitis lutheri with introduced C. tetralineata through a hybrid swarm following the artificial connection of ricer systems. Ecol. Evol., 4: 1451-1465. Lee, I.R., H. Yang, J.H. Kim, K.Y. Kim and I.C. Bang. 2009. Identification of a natural hybrid between the striped spine loach Cobitis tetralineata and the king spine loach Iksookimia longicorpa by analyzing mitochondrial COI and nuclear RAG1 sequence. Koran J. Ichthyol., 21: 287-290. (in Korean) Lee, T.Y. and S.Y. Kim. 1987. Experimental studies on the mechanism of reproductive cycle in the bluegill, Lepomis macrochirus. Korean J. Fish. Aquat. Sci., 20: 489-500. (in Korean) Lee, W.O., I.R. Lee, H.Y. Song and I.C. Bang. 2008. Genetic differentiation of the largemouth bass Micropterus salmoides from the major rivers and reservoirs in Korea assessed by AFLP. Korean J. Limnol., 41: 395-401. (in Korean) Mori, T. 1935. Studies on the geographical distribution of freshwater fish in Korea. Bull. Biogeogr. Soc. Jap., 7: 35-61. NFRDI. 2007. Ecological survey and management strategy of introduced freshwater fishes. Report of national fisheries research and development institute (inland fisheries research institute). Gapyeong, Korea, 81pp. (in Korean) Park, S.C., K.Y. Lee, T.B. Choi, W.M. Kim, M.J. Kim and J.S. Choi. 2013. The fish fauna and disturbance of geographical distributions in the eastern civilian control line of Korea. J. Environ. Impact Assess., 22: 27-37. (in Korean) Slechtova, V., J. Bohlen and H.H. Tan. 2007. Families of Cobitidea (Teleostei: Cypriniformes) as revealed from nuclear genetic data and the position of the mysterious genera Barbucca, Psilorhynchus, Swepenticobitis and vaillantella. Mol. Evol., 44: 1358-1365. Smith, C., M. Reichard, P. Jurajda and M. Przybylski. 2004. The reproductive ecology of the European bitterling (Rhodeus sericeus). J. Zool., Lond., 262: 107-124. Song, H.B. and O.K. Kwon. 1993. Ecology of Coreoleuciscus splendidus Mori (Cyprinidae) in Hongchon River. Korean J. Limnol., 3: 235-244. (in Korean) Song, H.B. and G.M. Park. 2006. A molecular genetic variation among intra-populations of Korean shiner, Coreoleucuscus splendidus Mori (Cyprinidae)., Koran J. Ichthyol., 18: 78-.
(12) 12. 송하윤·김재훈·서인영·방인철. 86. Song, H.B., M.S. Byeon, D.W. Kang, C.Y. Jang, J.S. Moon and H.K. Park. 2012. Population structure of bluegill, Lepomis macrochirus in lakes of the Han river system, Korea. Koran J. Ichthyol., 24: 278-286. (in Korean) Song, H.H. 1976. Study on the spawning and early development of Coreoleuciscus splendidus Mori. Master’s thesis, Kyunghee University, 23pp. (in Korean) Song, H.Y. and I.C. Bang. 2009. Genetic variation of Coreoleuciscus splendidus populations from four major river in Korea as assessed by RAPD PCR. Korean J. Ichrhyol., 21: 129133. (in Korean) Song, H.Y. and I.C. Bang. 2015. Coreoleuciscus aeruginos (Teleostei: Cypriniformes: Cyprinidae), a new species from the Seomjin and Nakdong rivers, Korea. Zootaxa, 3931: 140150. Song, H.Y., K.Y. Kim, M.G. Yoon, Y.K. Nam, D.S. Kim and I.C. Bang. 2010. Genetic variation of Coreoleuciscus splendidus population (Teleostei; Cypriniformes) from four major river drainage system in South Korea as assessed by AFLP markers. Genes & Genomics, 32: 199-205.. Sonnenberg, R., A.W. Nolte and D. Tautz. 2007. An evaluation of LSU rDNA D1-D2 sequences for their use in species identification. Frontiers in Zool., 4: 6. Thompson, J.D., T.J. Gibson, F. Plewniak, F. Jeanmougin and D.G. Higgins. 1997. The CLUSTAL X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nuclecis Acids Res., 24: 173-216. Wendel, J.F. and J.J. Doyle. 1998. Phylogenetic incongruence: window into genome history and molecular evolution. In: Soltis, D.E., Soltis, P.S., Doyle, J.J. (eds.), Molecular systematics of plants II: DNA sequencing. Kluwer academic publishers, Boston, pp. 265-296. Wu, G., J. Luo, K. Chen, C. Xian, G. Lin, Z. Wang, Z. Lin, J. Luo and K. Pan. 1990. Analysis and comparison of morphology and cytology between the F sub (1) hybrid catfish (Claris fuscus × C. lazera) and its parents. Acta Hydrobiologica Sinica, Wnhan., 14: 328-335. Yoon, Y.E., I.R. Lee, S.Y. Park, E.J. Kang, E.O. Kim, S.K. Yang, Y.K. Nam and I.C. Bang. 2009. Genetic identification of hybrids between Rhodeus uyekii and R. notatus by sequence analysis of RAG-1 gene. J. Aquacult., 22: 79-82. (in Korean).
(13)
수치


관련 문서
[14] OECD International Energy Agency, World Energy Outlook 2017, OECD Publishing, Paris (2017). [15] OECD International Energy Agency, World Energy Outlook 2014, OECD
L38-76 Air space Commission Implementing Decision (EU) 2017/259 of 13 February 2017 c oncerning certain revised performance targets and appropriate measures included in
L69-31 Food Commission Implementing Decision (EU) 2017/450 of 13 March 2017 aut horising the placing on the market of lactitol as a novel food ingredient un der Regulation
Commission Regulation (EU) 2017/492 of 21 March 2017 amending Regul ation (EC) No 883/2004 of the European Parliament and of the Council on the coordination of social
[r]
그림에서 왼쪽은 현재의 상태를 표현한 것으로 물 -에너지-식량 연관관계가 명확하게 분석되지 않아 (Unknown and unbalanced) 가용 수자원으로부터 생산을 위한 물로
여러 종류의 식당을 , 일반식당, 패밀리레스토랑, 고급식당, 계약급식업체로 나누어 보면 일반식당에서는 고객이 남기고 간 음식물쓰레기
[14] OECD International Energy Agency, World Energy Outlook 2017, OECD Publishing, Paris (2017). [15] OECD International Energy Agency, World Energy Outlook 2014, OECD
