PREVENTION RESEARCH □ORIGINAL ARTICLE □
246 책임저자:김기영, 690-756, 제주특별자치도 제주시 제주대학로 66
제주대학교 해양과학대학 해양과학부 면역학실험실 Tel: 064-754-3427, Fax: 064-756-3493
E-mail: [email protected]
접수일:2007년 10월 10일, 게재승인일:2007년 12월 7일
Correspondence to:Gi-Young Kim
Faculty of Applied Marine Science, Cheju National University, 66, Jejudaehakno, Jeju 690-756, Jeju Special Self-Governing Province, Korea Tel: +82-64-754-3427, Fax: +82-64-756-3493
E-mail: [email protected]
Sulforaphane-induced Apoptosis is Regulated by Bcl-2 and Caspase-3 in Human Leukemia U937 Cells through
Modulation of the ERK and Akt Pathways
Cheng-Yun Jin1, Dong-Oh Moon2, Yung Hyun Choi3, Cheol Park3, Mun-Ock Kim3, Jae-Dong Lee1, Tae-Ho Lee1 and Gi-Young Kim2
1Department of Microbiology, College of Natural Sciences, Pusan National University, Busan 609-735,
2Faculty of Applied Marine Science, Cheju National University, Jeju Special Self-Governing Province, Jeju 690-756,
3Department of Biochemistry, Dongeui University College of Oriental Medicine, Busan 614-714, Korea
Sulforaphane (SFN) is a major isothiocyanate compound shown to induce anti-proliferation and apop- tosis in solid human cancer cell lines. However, the mechanisms responsible for SFN-induced apoptosis are poorly understood in leukemia cells. In the present study, we first investigated the role of the anti-apoptotic protein, Bcl-2, in response to SFN-mediated apoptosis in human leukemia U937 cells. We also investigated whether SFN-induced apoptosis involved mitogen-activated protein kinases (MAPKs) and the Akt pathway. Treatment with 4μM SFN for 48 h resulted in a significant induction of apoptosis in U937 cells. This effect was characterized by accumulation of cells with sub-G1 phase DNA content, caspase activation, and poly (ADP-ribose) polymerase (PARP) degradation. In contrast, U937/Bcl-2 exposed to the same SFN treatment only exhibited a slight induction of apoptotic features. A specific Bcl-2 inhibitor, HA14-1, significantly increased SFN-induced apoptosis with caspase-3 activation. SFN treatment also increased extracellular-signal regulating kinase (ERK) phosphorylation in U937 cells, and combined treatment with a specific ERK inhibitor (PD98059) and SFN appeared to significantly inhibit SFN-induced apoptosis by interfering with caspase-3 activation. In contrast, treatment with SFN decreased the phosphorylation of Akt, which resulted in significant induction of apoptosis and combined treatment with Akt upstream inhibitor LY294002 induced more potent apoptosis. These results indicate that SFN-induced apoptosis is regulated by Bcl-2 and caspase in human leukemic U937 cells through modulation of the ERK and Akt pathways. (Cancer Prev Res 12, 246-255, 2007)
Key Words: Sulforaphane, Bcl-2, Caspase-3, Mitogen-activated protein kinase
INTRODUCTION
Many anti-tumor agents kill chemosensitive leukemic cells via an apoptosis through a variety of molecular targets.1,2) Apoptosis is a genetically encoded cell death program characterized by a distinct set of morphological and biochemical changes.3∼6) Apoptosis is affected by a number of factors, including tumor cell type, differentiation status, and the
presence of growth factors or oncogenes.4) Several genes have been identified that can either induce or repress apoptosis.
Bcl-2 was initially identified as the product of the gene encoded at the breakpoint of the chromosomal translocation that occurs in the majority of non-Hodgkin’s B-cell lymphomas.7,8) Bcl-2 can protect cells from apoptosis in response to such diverse stimuli as viral infection, hypoxia, ionizing radiation, or chemotherapeutic agents.9∼13) Bcl-2 preserves the integrity of the outer mitochondrial membrane and thereby prevents the
release of pro-apoptotic factors into the cytoplasm.14∼16) Although the ability of Bcl-2 to prevent the mitochondrial release of pro-apoptotic factors has been well documented, other functions of Bcl-2 in the upstream and downstream regulation of apoptosis remain poorly understood. Evidence suggests that overexpression of Bcl-2 can inhibit apoptosis in response to a variety of stimuli. This inhibition of apoptosis may involve modulation of caspase-3, PARP degradation,17,18) mitogen-activated protein kinase (MAPK), and Akt path- ways.19)
Sulforaphane (SFN; 1-isothiocyanato-4-(methylsulfinyl)-butane) is a naturally occurring member of the isothiocyanate-family of chemopreventive agents that has received particular attention because of its anticancer effects. SFN was discovered more than a decade ago and is a potent inducer of protective enzymes that provide defense against cancer-causing chemicals.20) Both intrinsic and extrinsic caspase cascades were shown to contribute to cell death in response to this highly promising cancer chemopreventive agent in solid tumor cells.21) SFN was also proven to be effective at inducing apoptosis in HepG2 cells through activation of the caspase-3 cascade.22) SFN was also reported to inhibit tubulin polymerization in breast cancer cells23) and to attenuate histone deacetylase activity in prostate epithelial cells.24) However, little is known about the molecular and the role of Bcl-2 of in SFN-induced apoptosis.
In this study, we investigated the response of human leukemia U937 cells to SFN-mediated apoptosis. Our results demonstrated that SFN-induced apoptosis was associated with Bcl-2 expression, caspase activation, PARP degradation, and signaling through the ERK and Akt pathways. We further investigated the ability of Bcl-2 overexpression to prevent SFN-induced apoptosis in U937 cells by a mechanism that interferes with the activity of caspase-3.
MATRIALS AND METHODS 1. Reagents
Fetal bovine serum (FBS), RPMI-1640, penicillin-strepto- mycin, and trypsine-EDTA were purchased from Gibco BRL (Gaithersburg, MD). SFN, propidium iodide (PI), paraform- aldehyde, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT), RNase A, HA-14-1, and proteinase K were purchased from Sigma (St. Louis, MO). Caspase activity assay kits were obtained from R&D systems (Minneapolis, MN). An
enhanced chemiluminescence (ECL) kit was purchased from Amersham (Arlington Heights, IL). Caspase-3 inhibitor 1 (z-DEVD-fmk), phosphoinositide 3-kinase (PI3K) inhibitor LY294002, extracellular signal-regulated kinase (ERK) inhi- bitor PD98059, c-Jun N-terminal kinase (JNK) inhibitor SP600125, and p38 MAPK inhibitor SB203580 were obtained from Calbiochem (San Diego, CA). Any other chemicals not specifically cited here were purchased from Sigma.
2. Antibodies
Anti-Bcl-2, anti-Bax, anti-p38, anti-pp38, anti-JNK, anti- phospho (p)-JNK, anti-ERK, anti-p-ERK, anti-Akt, anti-p-Akt, anti-caspase-3, anti-caspase-8, and anti-caspase-9 were purchased form Santa Cruz Biotechnology (Santa Cruz, CA). Anti-PARP was purchased from PharMingen (San Diego, CA) and anti-β- actin from Sigma. Peroxidase-labeled anti-rabbit and anti- mouse polyclonal immunoglobulins were purchased from Ame- rsham.
3. Cell line and cell culture
Human myeloid leukemia U937 cells were purchased from the ATCC (Rockville, MD) and cultured in RPMI-1640 medium supplemented with 10% FBS and 1% penicillin/
streptomycin in a humidified atmosphere with 5% CO2 incu- bator at 37oC. Bcl-2 overexpressing U937 cells (U937/Bcl-2) were generated using a pMAX vector containing the human Bcl-2 gene (provided by T.K. Kwon, Keimyung, Daegu, South Korea). Cells were cultured in the absence and presence of variable concentrations of SFN for the indicated times.
4. Cell viability
The cells were seeded at a density of 2×105 cells/ml and then incubated under various conditions as indicated. MTT (0.5 mg/ml) was subsequently added to each well. After 4 h of additional incubation, 100μl of a solution containing 10% SDS (pH 4.8) plus 0.01 N HCl was added to dissolve the crystals.
The absorption values at 570 nm were determined with a ELISA plate reader (Multiskan EX; Labsystems, MA).
5. Flow cytometric analysis
Cells were washed with PBS, pelleted by low speed centrifugation, resuspended in citrate buffer, and treated with RNase A. Nuclei were stained with a PI solution. The DNA content in each cell nucleus was determined by a FACSCalibur
flow cytometer (Becton-Dickinson, San Jose, CA), acquisition software Cell Quest (Becton Dickinson), and the DNA curve fitting program Modfit LT (Becton-Dickinson).
6. Gel electrophoresis and immunoblotting
The cells were harvested and lysed, and protein concen- trations were quantified using the BioRad protein assay (BioRad Lab., Hercules, CA) following the procedure described by the manufacturer. Immunoblotting was performed. Briefly, an equal amount of protein was subjected to electrophoresis on SDS-polyacrylamide gels and transferred to nitrocellulose membranes by electroblotting. Blots were probed with the desired antibodies for 1 h, incubated with diluted enzyme- linked secondary antibody and then visualized by the ECL according to the recommended procedure (Amersham).
7. Caspase activity assay
Caspase activities were determined with colorimetric assays using a caspase-3, -8, and -9 activity kits according to the manufacturer's protocol. Briefly, the cells were lysed in a lysis buffer for 30 min on an ice bath. The lysed cells were centrifuged at 12,000 g for 10 min, and 100μg of the protein was incubated with 50μl of a reaction buffer and 5μl of the colorimetric tetrapeptides, Asp-Glu-Val-Asp (DEVD)-p-nitroaniline (pNA) for caspase-3, Ile-Glu-Thr-Asp (IETD)-pNA for caspase-8 and Leu-Glu-His-Asp (LEHD)-pNA for caspase-9, respectively, at 37oC for 2 h. The optical density of the reaction mixture was quantified spectrophotometrically at a wavelength of 405 nm using a VERSAmax tunable microplate reader.
8. Statistical analysis
All experiments were performed in triplicate. The results of multiple observations were presented as the means±SD of at least three separate experiments. Statistical significance was determined by Student's t-test. A value of p<0.05 was considered to be significant.
RESULTS
1. Bcl-2 overexpression attenuates SFN-induced apoptosis in U937 cells
To determine the role of Bcl-2 in SFN-induced apoptosis, U937 cells were stably transfected with either the human Bcl-2 cDNA (U937/Bcl-2) or vector alone (U937). G418-resistant
clones found to overexpress Bcl-2 were selected and used for subsequent experiments. Immunoblot analysis revealed that control U937 and U937/Bcl-2 expressed comparable levels of the loading control β-actin, however, expression of Bcl-2 was approximately 4-fold greater in U937/Bcl-2 than in U937 cells (Fig. 1A).
U937 and U937/Bcl-2 cells were then incubated for 48 h in the presence of increasing concentrations of SFN (0∼4μM).
Following SFN incubation, the extent of proliferation inhibition and apoptosis was analyzed by performing MTT reduction assays and determining the percentage of cells with sub-G1
DNA content by flow cytometry, respectively. As shown in Fig. 1B, treatment with SFN for 48 h resulted in a dose- dependent decrease in U937 cell viability (69±5% at 4μM SFN). In contrast, U937/Bcl-2 cells did not exhibit a dramatic decrease in cell viability following incubation with SFN (28±5% at 4μM SFN). Furthermore, treatment with SFN for 48 h resulted in a dose-dependent accumulation of U937 cells with sub-G1 phase DNA content (Fig. 1C). Treatment of U937 cells with 4μM SFN resulted in significant increase in percentage of cells with sub-G1 phase DNA content (30±6%), however U937/Bcl-2 cells did not exhibit such a dramatic accumulation (12±3%). These findings indicate that SFN- induced apoptosis in U937 cells can be attenuated by the overexpression of the Bcl-2 protein.
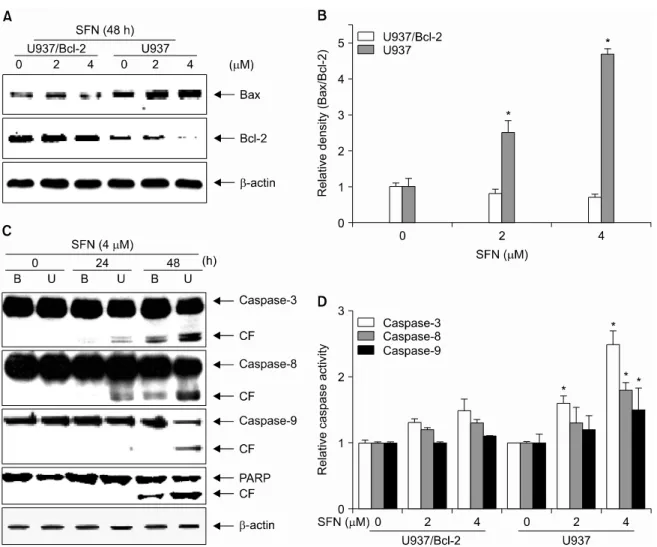
2. Bax/Bcl-2 ratio and caspase activation are major targets in SFN-induced apoptosis
The ratio between Bcl-2 protein family members, especially Bax and Bcl-2, plays a pivotal role in apoptotic signaling. The relative levels of these pro- and anti-apoptotic proteins integrate a wide array of diverse upstream signals that determine cell fate. Therefore, we used immunoblot analysis to investigate the effect SFN treatment on the levels of pro-apoptotic Bax and anti-apoptotic Bcl-2. As shown in Fig.
2A, treatment for 48 h with 2 or 4μM SFN resulted in decreased levels of Bcl-2 in U937 cells, but increased the levels of Bax U937/Bcl-2 cells. A densitometric analysis revealed that SFN treatment increases the Bax/Bcl-2 ratio to favor pro- apoptosis in a dose-dependent manner (Fig. 2B). These results indicated that the expression levels of Bcl-2 family members are involved in SFN-induced apoptosis in U937 cells.
Caspases are also known to be very important regulators of apoptosis in response to a variety of apoptotic stimuli.
Fig. 1. Bcl-2 overexpression inhibits SFN-induced apoptosis in U937 cells. (A) Immunoblot analysis of cell lysate from control (U937) or Bcl-2 transfected (U937/Bcl-2) cells with anti-Bcl-2 antibody. β-Actin was used as an internal control. (B) The cells were seeded at 2×105 cells/ml and treated with different concentrations of SFN for 48 h. Cell viability was determined by MTT assay. (C) The percentage of cells with sub-G1 phase DNA content was analyzed by flow cytometry. Each point represents the mean±SD of three independent experiments. The significance was determined by Student's t-test (*p<0.05 vs. untreated control).
Therefore, we additionally examined the status of various caspases during SFN-induced apoptosis. As shown in Fig. 2C and 2D, exposure to SFN increased levels of the cleavage and activation of caspases-3, -8, and -9 in U937 cells. In contrast, the overexpression of Bcl-2 attenuated SFN-induced activation of caspases and cleavage of PARP. These data clearly indicate that SFN-induced apoptosis is associated with caspase acti- vation and that this activation is attenuated by Bcl-2 over- expression.
3. Bcl-2 inhibitor HA14-1 increases SFN-induced apoptosis in U937 cells
To further elucidate the role of the Bcl-2 protein in SFN-mediated apoptosis, we next investigated the effect of SFN in the absence and presence of Bcl-2 inhibitor HA14-1.
We then attempted to determine whether HA14-1 could increase apoptotic signaling in our experimental model.
HA14-1 (10μM) treatment alone did not significantly alter the
proportion of cells with sub-G1 phase DNA content, cell viability, or caspase-3 activity (Fig. 3A∼C). However, co- administration of SFN and HA14-1 significantly increased accumulation of U937 cells with sub-G1 phase DNA content (79±3%), increased caspase-3 activity, and decreased cell viability (21±3%) compared to cells treated with SFN alone.
Moreover, we also analyzed expression of Bcl-2 and cleavage of PARP and caspase-3 following combined treatment with HA14-1 and SFN. As shown in Fig. 3D, HA14-1 alone did not alter expression levels of Bcl-2 or the cleavage of PARP and caspase-3. However, combined treatment with SFN and HA14-1 resulted in a significant decrease in Bcl-2 expression and a marked increase the cleavage of PARP and caspase-3 relative to cells treated with SFN alone. Taken together, these data indicate that endogenous Bcl-2 expression suppresses caspase-3 activation and cleavage of PARP, which are required for induction of apoptosis.
Fig. 2. SFN increases the Bax/Bcl-2 ratio and upregulates caspase activity. Cells were treated with SFN (4μM) for the indicated times. Equal amounts of cell lysates (30μg of total protein) were resolved by SDS-PAGE, transferred to nitrocellulose, and probed with specific antibodies [(A) anti-Bax and anti-Bcl-2; (C) anti-caspase-3, anti-caspase-8, anti-caspase-9, and anti-PARP]. (B) The data from the immunoblot analysis of Bax and Bcl-2 were used to evaluate the effect of SFN on the Bax/Bcl-2 ratio. (D) After 48 h treatment with SFN, caspase activity was determined using a caspase assay kit. Data are expressed as mean±SD of three independent experiments. The significance was determined by Student's t-test (*p<0.05 vs. control). B, U937/Bcl-2; U, U937;
and CF, cleaved form.
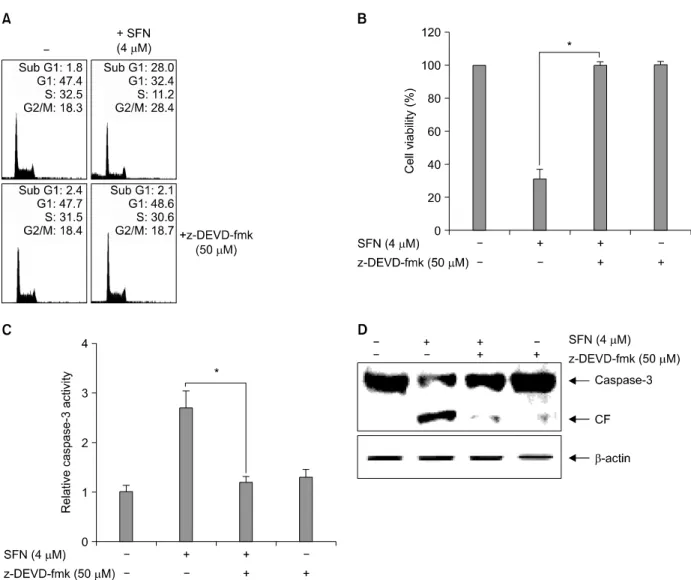
4. SFN-induced apoptosis appears through caspase-3 activation in U937 cells
We next addressed the role of caspase-3 in SFN-induced apoptosis by investigating whether the specific caspase-3 inhibitor z-DEVD-fmk could block apoptosis in U937 cells.
SFN treatment resulted in substantial accumulation of U937 cells with sub-G1 phase DNA content (Fig. 4A), decreased cell viability (Fig. 4B), and activation of caspase-3 (Fig. 4C).
However, z-DEVD-fmk pre-treated U937 cells exhibited significantly lower levels of accumulation with sub-G1 phase DNA content (3±2%). Pre-treated cells also had lower
caspase-3 activity and increased cell viability (96±4%) follow- ing SFN-treatment. As shown in Fig. 4D, the inhibitor signifi- cantly attenuated cleavage of caspase-3 in the presence of SFN.
These results clearly suggest that caspase-3 closely regulates SFN-mediated apoptosis in U937 cells.
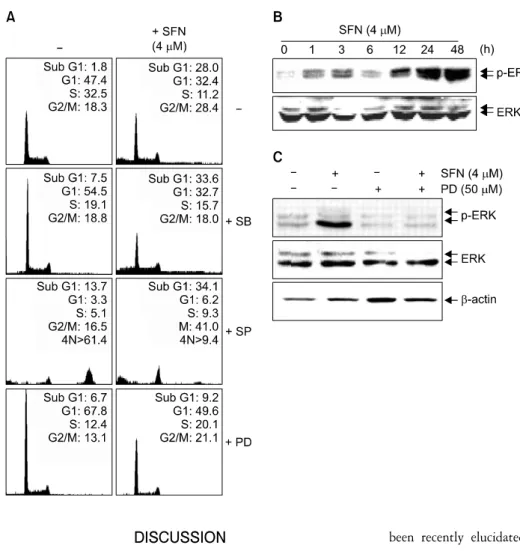
5. The ERK pathway is important for the induction of apoptosis by SFN
We next investigated the effect of inhibitors for MAPK pathways on SFN-induced apoptosis in order to determine whether these signaling pathways are involved in the observed apoptotic response. As shown in Fig. 5A, pre-treatment with
Fig. 3. Bcl-2 inhibitor HA14-1 significantly increases SFN-induced apoptosis in U937 cells. Cells were incubated with SFN for 48 h after 1 h pretreatment with HA14-1 (10μM). (A) DNA content was analyzed by flow cytometry and (B) cell viability was determined by MTT reduction. (C) Caspase-3 activity was determined using a commercially available kit. (D) Equal amounts of cell lysate (30μg) were resolved by SDS-PAGE, transferred to nitrocellulose, and probed with specific antibodies (anti-caspase-3 and anti-PARP). β-Actin was used as an internal loading control. Each point represents the mean±SD of three independent experiments. The significance was determined by Student's t-test (*p<0.05 vs. SFN treatment). CF, cleaved form.
PD98059 (a potent inhibitor of ERK) significantly decreased the number of cells with sub-G1 DNA content from 28±5%
to 10±3%. However, pre-treatment with SP600125 (a potent inhibitor of JNK) or SB203589 (a specific inhibitor of p38 MAPK) did not exert a statistically significant effect on accumulation of cells with sub-G1 phase DNA content follow- ing SFN treatment.
We next performed time-dependent experiments in order to determine how expression and phosphorylation levels of ERK were affected. As shown in Fig. 5B, the levels of phospho- rylated ERK increased significantly following 12 h treatment
with SFN while total ERK protein levels remained constant throughout the course of the experiment. We also analyzed levels of phosphorylated and total ERK following combined treatment with SFN and PD98059. As shown in Fig. 5C, immunoblot analysis revealed that treatment with SFN alone increased the levels of phosphorylated ERK, however co-admi- nistration of PD98059 and SFN significantly decreased the levels of phosphorylated ERK. These results indicate that SFN treatment is likely to induce ERK activation, suggesting that this pathway is involved in promoting apoptosis.
Fig. 4. Caspase-3 inhibitor z-DEVD-fmk alleviates SFN-induced apoptosis in U937 cells. Cells were incubated with SFN for 48 h after 1 h pretreatment with z-DEVD-fmk (50μM). (A) DNA content was analyzed by flow cytometry and (B) cell viability was determined by MTT reduction. (C) Caspase-3 activity was determined using a commercially available kit. (D) Equal amounts of cell lysate (30μg of total protein) were resolved by SDS-PAGE, transferred to nitrocellulose, and probed with specific antibodies (anti-Bcl-2 and anti-caspase-3). β-Actin was used as an internal loading control. Each point represents the mean±SD of three independent experiments. The significance was determined by Student's t-test (*p<0.05 vs. SFN treatment). CF, cleaved form.
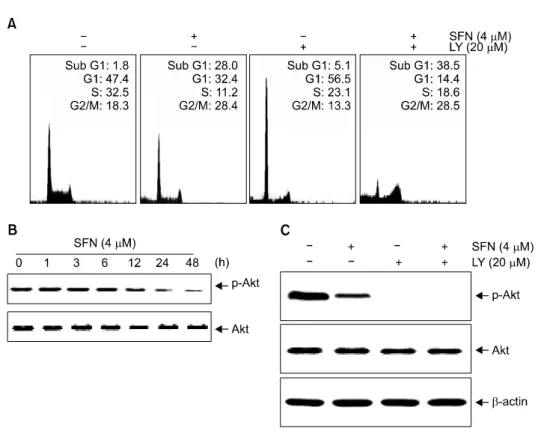
6. SFN induces Akt-mediated apoptosis in U937 cells
Akt is a downstream effector of PI3K that has been shown to mediate survival signaling. Therefore, we investigated whether the inhibition of the Akt pathway was necessary for SFN-induced apoptosis. As shown in Fig. 6A, treatment with LY294002 alone resulted in increased G1 arrest without evidence of apoptosis. Combined treatment with the inhibitor and SFN resulted in an increase of apoptosis, as determined by the percentage of cells with sub-G1 DNA content (41±6%).
We next performed SFN time-dependent experiments in order to determine how the expression and phosphorylation levels of Akt were affected. As shown in Fig. 6B, the levels of phospho- rylated Akt decreased significantly following exposure to SFN while total Akt protein levels remained constant throughout the course of the experiment. However, co-administration of LY294002 and SFN resulted in a significant decrease and phosphorylated levels of Akt relative to cells treated with LY294002 alone (Fig. 6C). These results also indicate a role for Akt downregulation in response to SFN treatment in U937 cells since LY294002 alone has significant cytotoxicity.
Fig. 5. SFN-induced apoptosis is mediated through ERK activation in U937 cells. (A) Cells were treated with PD98059 (50μM), SB203580 (20μM), or SP600125 (15μM) for 1 h prior to SFN treatment for 48 h. The cells were stained with PI (50μg/ml) and analyzed by flow cytometry. (B) The cells were treated with SFN (4μM) for varying time intervals. (C) Cells were incubated with SFN (4μM) for 48 h after a 1 h pretreatment with PD98059 (50μM). Equal amounts of cell lysates (30μg total protein) were resolved by SDS-PAGE, transferred to nitrocellulose, and probed with specific antibodies (anti-ERK and anti-p-ERK). β-Actin was used as an internal control. β-Actin was used as an internal control. The results are from one representa- tive experiment of three performed and show similar patterns.
DISCUSSION
Since the discovery of Bcl-2, several theories have been proposed to unravel the antiapoptotic properties of this protein.25∼28) The potent anti-apoptotic activity of Bcl-2 is likely ascribed to its ability to control several key steps of death signaling. Bcl-2 can form ion channels in biological mem- branes, which may affect apoptosis by influencing the perme- ability in intracellular membranes and release of mitochondrial contents into the cytoplasm, causing activation of caspase-3 and PARP cleavage.29,30) However, overexpression of Bcl-2 protein may rescue cells from apoptosis by blocking the release of mitochondrial contents, maintaining membrane integrity, or by an as yet unrecognized function. Our data demonstrated that Bcl-2 and caspase-3 are major mediators of SFN-induced apoptosis in U937 cells. Overexpression of Bcl-2 protein may rescue cells from apoptosis through the inactivation of caspase-3.
The caspase family of aspartate-specific cysteine proteases plays a critical role in regulating apoptosis. Many of the key biochemical components required for caspase activation have
been recently elucidated.31) Caspase signaling is initiated and propagated by proteolytic autocatalysis and the cleavage of downstream caspases and substrates, such as PARP and lamin A.32) Caspase-3 is an especially critical executioner of apoptosis that is either partially or totally responsible for the proteolytic cleavage of many key proteins, including PARP. PARP is important for cell viability and the cleavage of PARP facilitates cellular disassembly and serves as a marker of cells undergoing apoptosis.30) In this study, we demonstrated that SFN induces apoptosis through the activation and cleavage of caspase-3, -8, and -9. Treatment with SFN in the presence z-DEVD-fmk inhibited cell death and apoptotic body formation, which suggest that SFN-induced apoptosis was caused by caspase-3- dependent cell death.
The ERK and Akt pathways play critical roles in cell survival in many physiological and pathological settings.33,34) Our observation suggests an unexpected role for increased activation of ERK in SFN-induced apoptosis. In support of our results, some studies recently reported that ERK pathway induces apoptosis in human colon cancer cells as well as other cell types,35,36) suggesting that the ERK pathway may play a variety
Fig. 6. SFN-induced apoptosis is mediated through downregulation of the Akt pathways in U937 cells.
Cells were treated with LY294002 for 1 h before SFN treatment. (A) After 48 h-incubation, the cells were stained with PI (50μg/ml) and analyzed by flow cytometry. (B) The cells were treated with SFN (4 μM) for varying time intervals. (C) Cells were incubated with SFN (4 μM) for 48 h after a 1 h pre- treatment with LY294002 (20μM).
Equal amounts of cell lysate (30μg of total protein) were resolved by SDS-PAGE, transferred to nitro- cellulose, and probed with spe- cific antibodies (anti-Akt and anti- p-Akt). β-Actin was used as an internal loading control.
of roles, depending on the cell type and toxic stimulus. In addition, we investigated that SFN induces a downregulation of Akt, and pretreatment of LY294002 significantly increased SFN-induced apoptosis indicating that Akt might have a survival role in SFN-induced U937 cell apoptosis. These data strongly suggest that SFN-induced apoptosis is associated with the ERK and Akt pathways.
In conclusion, we have demonstrated that SFN promotes a significant induction of apoptosis via regulation of Bcl-2 family proteins and caspase activity. SFN-induced apoptosis was associated with upregulation of the ERK pathway and down- regulation of the Akt pathway. Therefore, SFN is likely to represent a promising molecule for cancer chemoprevention and chemotherapy in leukemic cells.
ACKNOWLEDGMENTS
We thank Prof. TK KWON for providing Bcl-2 over- expressing U937 cells. This work was supported by the research grant of the Cheju National University in 2007.
REFERENCES
1) Fisher DE. Apoptosis in cancer therapy: crossing the threshold.
Cell 78, 539-542, 1994.
2) Hickman JA. Apoptosis induced by anticancer drugs. Cancer Metastasis Rev 11, 121-239, 1992.
3) Cohen JJ. Apoptosis. Immunol Today 14, 126-130, 1993.
4) White E. Life, death, and the pursuit of apoptosis. Genes Dev 10, 1-15, 1996.
5) Williams GT, Smith CA. Molecular regulation of apoptosis:
genetic controls on cell death. Cell 74, 777-779, 1993.
6) Wyllie AH, Kerr JF, Currie AR. Cell death: the significance of apoptosis. Int Rev Cytol 68, 251-306, 1980.
7) Tsujimoto Y, Faffe E, Cossman J, Gorham J, Novwell PC, Croce CM. Clustering of break points on chromosome II in human B-cell neoplasms with the t(11;14) chromosome trans- location. Nature 315, 340-334, 1985.
8) Tsujimoto Y, Croce CM. Analysis of the structure, transcripts and protein products of bcl-2, the gene involved in human follicular lymphoma. Proc Natl Acad Sci USA 83, 5214-5218, 1986.
9) Lawrence MS, Ho DY, Sun GH, Steinberg GK, Sapolsky RM.
Overexpression of bcl-2 with herpes simplex virus vectors protects CNS neurons against neurological insults in vitro and in vivo. J Neurosci 16, 486-496, 1996.
10) Shimizu G, Eguchi Y, Kosaka H, Kamiike W, Matsuda H, Tsujimoto Y. Prevention of hypoxia-induced cell death by Bcl-2 and Bcl-XL. Nature 374, 811-813, 1995.
11) Tudor G, Aguilera A, Halverson DO, Laing ND, Sausville EA. Susceptibility to drug-induced apoptosis correlates with differential modulation of Bad, Bcl-2 and Bcl-XL protein levels. Cell Death Differ 7, 574-586, 2000.
12) Gillardon F, Moll I, Meyer M, Michaelidis TM. Alterations in cell death and cell cycle progression in the UV-irradiated epidermis of bcl-2-deficient mice. Cell Death Differ 6, 55-60, 1999.
13) Kyprianou N, King ED, Bradbury D, Rhee JG. Bcl-2 overexpression delays radiation-induced apoptosis without affecting the clonogenic survival of human prostate cancer cells. Int J Cancer 70, 341-348, 1997.
14) Dlugosz PJ, Billen LP, Annis MG, Zhu W, Zhang Z, Lin J, Leber B, Andrews DW. Bcl-2 changes conformation to inhibit Bax oligomerization. EMBO J 25, 2287-2296, 2006.
15) Qiao X, Chen X, Wu D, Ding R, Wang J, Hong Q, Shi S, Li J, Xie Y, Lu Y, Wang Z. Mitochondrial pathway is responsible for aging-related increase of tubular cell apoptosis in renal ischemia/reperfusion injury. J Gerontol A Biol Sci Med Sci 60, 830-839, 2005.
16) Thees S, Hubbard GB, Winckler J, Schultz C, Rami A.
Specific alteration of the Bax/Bcl2 ratio and cytochrome c without execution of apoptosis in the hippocampus of aged baboons. Restor Neurol Neurosci 23, 1-9, 2005.
17) Kim YH, Park JW, Lee JY, Surh YJ, Kwon TK. Bcl-2 overexpression prevents daunorubicin-induced apoptosis through inhibition of XIAP and Akt degradation. Biochem Pharmacol 66, 1779-1786, 2003.
18) Park JW, Choi YJ, Suh SI, Baek WK, Suh MH, Jin IN, Min DS, Woo JH, Chang JS, Passaniti A, Lee YH, Kwon TK.
Bcl-2 overexpression attenuates resveratrol-induced apoptosis in U937 cells by inhibition of caspase-3 activity. Carcinogenesis 22, 1633-1639, 2001.
19) Yu C, Mao X, Li WX. Inhibition of the PI3K pathway sensitizes fludarabine-induced apoptosis in human leukemic cells through an inactivation of MAPK-dependent pathway.
Biochem Biophys Res Commun 331, 391-397, 2005.
20) Fahey JW, Haristoy X, Dolan PM, Kensler TW, Scholtus I, Stephenson KK, Talalay P, Lozniewski A. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene- induced stomach tumors. Proc Natl Acad Sci USA 99, 7610- 7615, 2002.
21) Singh SV, Srivastava SK, Choi S, Lew KL, Antosiewicz J, Xiao D, Zeng Y, Watkins CS, Trump DL, Lee YJ, Xiao H, Herman-Antosiewicz A. Sulforaphane-induced cell death in human prostate cancer cells is initiated by reactive oxygen species. J Biol Chem 20, 19911-19924, 2005.
22) Chi-Tai Y, Gow-Chin Y. Effect of sulforaphane on meta- llothionein expression and induction of apoptosis in human hepatoma HepG2 cells. Carcinogenesis 26, 2138-2148, 2005.
23) Jackson SJ, Singletary KW. Sulforaphane: a naturally occur- ring mammary carcinoma mitotic inhibitor, which disrupts
tubulin polymerization. Carcinogenesis 25, 219-227, 2004.
24) Myzak MC, Hardin K, Wang R, Dashwood RH, Ho E.
Sulforaphane inhibits histone deacetylase activity in BPH-1, LNCap and PC-3 prostate epithelial cells. Carcinogenesis 27, 811-819, 2006.
25) Mow BM, Blajeski AL, Chandra J, Kaufmann SH. Apoptosis and the response to anticancer therapy. Curr Opin Oncol 13, 453-462, 2001.
26) Murph AN, Bredesen DN, Cortopassi G, Wang E, Fiskum G. Bcl-2 potentiates the maximal calcium uptake capacity of neural cell mitochondria. Proc Natl Acad Sci USA 93, 9893- 9898, 1996.
27) Yang J, Liu X, Bhalla K, Kim CN, Ibrado AM, Cai J, Peng TI, Jones DP, Wang X. Prevention of apoptosis by Bcl-2:
release of cytochrome c from mitochondria blocked. Science 275, 1129-32, 1997.
28) Kluck RM, Bossy-Wetzel E, Green DR, Newmeyer DD. The release of cytochrome c from mitochondria: a primary site for Bcl-2 regulation of apoptosis. Science 275, 1132-1136, 1997.
29) Fernandes-Alnemri T, Litwack G, Alnemri ES. CPP32, a novel human apoptotic protein with homology to Caenorhabditis elegans cell death protein Ced-3 and mammalian interleukin- 1beta-converting enzyme. J Biol Chem 269, 30761-30764, 1994.
30) Oliver FJ, de la Rubia G, Rolli V, Ruiz-Ruiz MC, de Murcia G, Murcia JM. Importance of poly (ADP-ribose) polymerase and its cleavage in apoptosis. Lesson from an uncleavable mutant. J Biol Chem 273, 33533-33539, 1998.
31) Stennicke HR, Salvesen GS. Properties of the caspases. Bio- chim Biophys Acta 1387, 17-31, 1998.
32) Hengartner MO. The biochemistry of apoptosis. Nature 407, 770-776, 2000.
33) Xia Z, Dickens M, Raingeaud J, Davis RJ, Greenberg ME.
Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 270, 1326-1331, 1995.
34) Kennedy SG, Wagner AJ, Conzen SD, Jordan J, Bellacosa A, Tsichlis PN. The PI 3-kinase/Akt signaling pathway delivers an anti-apoptotic signal. Genes Dev 11, 701-713, 1997.
35) Jakubikova J, Sedlak J, Mithen R, Bao Y. Role of PI3K/Akt and MEK/ERK signaling pathways in sulforaphane- and erucin- induced phase II enzymes and MRP2 transcription, G2/M arrest and cell death in Caco-2 cells. Biochem Pharmacol 69, 1543-1552, 2005.
36) Xu C, Shen G, Yuan X, Kim J, Gopalkrishnan A, Keum YS, Nair S, Kong AN. ERK and JNK signaling pathways are involved in the regulation of activator protein 1 and cell death elicited by three isothiocyanates in human prostate cancer PC-3 cells. Carcinogenesis 27, 437-445, 2006.