꽃송이버섯 추출물의 간세포에서 대사 활성 효과
백소정1, 조남준1, 조성진2, 김은미3*, 김기광1*
The Metabolic Activity of Sparassis crispa Extract in Hepatocytes
So-Jeong Baek1, Namjoon Cho1, Sung-Jin Cho2, Eun-Mi Kim3*, and Kee Kwang Kim1*
Received: 5 November 2019 / Revised: 29 December 2019 / Accepted: 9 January 2020
© 2020 The Korean Society for Biotechnology and Bioengineering
Abstract: Sulfonylurea treatment is used for diabetes mellitus, a metabolic disease caused by abnormal absorption of glucose in the body. But it is accompanied by side effects in vivo when taken for a long time. Therefore, there is a need for research on natural materials having similar effects to these therapeutic agents and having fewer side effects that can effectively regu- late blood sugar. In this study, we investigated the antioxidative efficacy of Sparassis crispa (S. crispa) extract and the activity of ATP, glycolysis, and mitochondrial membrane potential using human hepatocyte HepG2 cells. As a result, S. crispa extract showed high antioxidant activity and induced a significant increase in HepG2 cell activity and ATP production. S. crispa extract also significantly accelerated the process and mitochon- drial membrane potential. Taken together, these results suggest that S. crispa extract has the potential to be developed as a natu- ral material for improving diabetic and diabetic complications by activating hepatic energy metabolism.
Keywords: Sparassis crispa, glycolysis, mitochondria, hepato- cyte, metabolism
1. INTRODUCTION
천연물 유래 생리활성 기능이 뛰어난 식품의 개발은 질병의 치료 및 예방적 측면에서 매우 중요하다. 다양한 기능성 식 품 중에서도 버섯은 각종 영양소를 함유하고 있을 뿐만 아니 라 생리활성 물질도 풍부하여 예로부터 식용 및 약용으로 널 리 이용되어 온 식품 중의 하나이다 [1]. 그중 꽃송이버섯 (Sparassis crispa; S. crispa)은 항당뇨, 면역 증강, 혈중 콜레 스테롤 저하, 항암 및 항염증 효과 등 광범위한 생리활성 기 능을 가지고 있다 [2-7]. 꽃송이버섯은 담자균의 민주름버섯 목, 꽃송이버섯과에 속하는 갈색부후균으로 여름철에 낙엽 송을 비롯한 침엽수림에서 발생한다 [8]. 꽃송이버섯은 버섯 류 중에서도 β-glucan 함유량이 높은 것으로 알려졌다. 꽃송 이버섯의 β-glucan 함유량은 약 43.6%로 다른 버섯 (표고버 섯 33.50%, 애느타리버섯 25.57%, 상황버섯 23.92%)에 비해 매우 높다 [3,9]. Yan Cao etc. (2016) 연구진은 high-fat diet 마 우스 모델 및 제2형 당뇨병 마우스 모델에 baker's yeast β- glucan의 경구투여가 혈액 내 glucose level을 효과적으로 억 제한다는 연구 결과를 보고하였다 [10]. 또한 L Jenkins etc.
연구진은 제2형 당뇨 환자들에게 β-glucan 고함량 식이요법 을 진행한 결과 혈당수치를 낮추는 연구 결과를 보고하여 β- glucan이 혈액 내 glucose level을 억제하는 효능은 마우스 모 델 뿐만 아니라 임상연구를 통해서도 증명된 바 있다 [9]. 하 지만 아직 높은 농도로 β-glucan을 함유한 꽃송이버섯이 당 뇨병에 미치는 영향 및 분자세포 생물학적인 작용 메커니즘 에 관한 연구는 부족한 실정이다.
당뇨병은 비정상적인 체내 포도당의 흡수로 인한 대사질 환으로 제1형과 제2형으로 구분된다. 제1형 당뇨병은 췌장 베타 세포의 파괴를 초래하는 자가면역 질환이며, 제2형 당 뇨병은 주로 췌장 베타세포의 기능 장애와 인슐린 저항성의 조합으로 인해 점차적으로 포도당 조절이 손상되어 발병된다 [11-12]. 췌장 베타세포 (Pancreatic β-cell)는 혈액 내 포도당을
*These authors contributed equally.
1
충남대학교 생화학과
1
Department of Biochemistry, College of Natural Sciences, Chungnam National University, Daejeon 34134, Korea
Tel: +82-42-821-7525, Fax: +82-42-822-7548 E-mail: [email protected]
2
충북대학교 생물학과
2
Department of Biology, Chungbuk National University, Cheongju 28644, Korea
3
안전성평가연구소 예측독성연구본부
3
Department of Predictive Toxicology, Korea Institute of Toxicology, 141 Gajeong-ro, Yuseong-gu, Daejeon 34114, Korea
Tel: +82-42-610-8263, Fax: +82-42-610-8157 E-mail: [email protected]
Research Paper
인지하여 ATP 농도를 증가시키고 ATP-sensitive potassium channel을 차단해 미토콘드리아 막의 탈분극, Ca²+의 유입, 마지막으로 혈액 내로 인슐린을 분비한다 [13-14]. 분비된 인 슐린은 근육 및 지방조직으로의 포도당 흡수를 촉진하고 간 에서 포도당 생성을 감소시켜 혈당을 낮춘다 [15]. 따라서 혈 당을 효율적으로 낮추기 위해서는 췌장 베타세포의 ATP 생 성량 조절을 통해 인슐린의 분비를 촉진하는 작용이 필요하 다. 제2형 당뇨병 환자의 췌장 베타세포는 ATP 생성량이 감 소되어 있을 뿐만 아니라 인슐린 분비량이 감소되어 있다.
또한 근육 및 지방조직이 인슐린 작용에 반응하지 않는 인슐 린 저항성이 나타나며 그 결과 혈당이 증가한다 [16-17]. 이 는 고혈당으로 이어지며 고지혈증과 동맥경화 등 다양한 합 병증 위험을 증가시킨다 [15].
최근 급격한 경제성장 및 생활 수준의 향상으로 단순 당의 섭취와 고열량, 고지방 식이와 같은 식습관의 서구화 현상이 가속화되었고, 이러한 현상은 비만과 운동 부족을 초래하였 다 [18-19]. 그 결과 만성 대사질환 중 하나인 당뇨병의 발병 률이 지속해서 증가하고 있다 [20]. 비만은 인슐린 저항성과 산화적 스트레스를 유발한다고 알려져 있으며 A.C Maritim etc. 연구진들은 산화적 스트레스의 증가가 당뇨 발병을 유 도함을 보고하였다 [21-23]. 따라서 서구화된 식습관에 따른 산화적 스트레스는 당뇨와 합병증의 발병 및 악화 요인 중 하나이다. 산화적 스트레스는 활성 산소종 (Reactive oxygen species, ROS)이 세포의 항산화 능력보다 과다하여 발생하거 나, 항산화 방어체계의 활성 저하로 인해 발생하게 된다 [24]. 그 결과 탄수화물, 단백질, 지방 세포의 물질대사에 영 향을 미치며, genome 및 epigenome의 불안정성, 세포 손상, 염증 및 기능 손상으로 이어진다 [25-28]
현재 sulfonylurea계 약물은 베타세포의 인슐린 분비를 촉 진하는 가장 대표적인 제2형 당뇨병 치료제로 사용되고 있 지만, 약물 투여에 따라 저혈당증과 같은 생체 내 부작용이 동반된다 [29-30]. 따라서 효과적으로 혈당을 조절할 수 있으 며 항당뇨 질환에 높은 활성을 보이는 부작용이 적은 천연 소재에 대한 연구가 절실하다 [31]. 따라서 본 연구에서는 꽃 송이버섯 추출물에 대한 인간 간세포의 에너지 대사에 미치 는 영향을 분자적 수준으로 규명함으로써 꽃송이버섯 추출 물이 당뇨병 예방 및 치료에 있어서 기능성 소재로의 가능성 을 규명하였다.
2. MATERIALS AND METHODS
2.1. 꽃송이버섯 추출물 제조
꽃송이버섯은 화순군에 있는 백아산 꽃송이버섯 농장에서 구입하였다. 꽃송이버섯 50 g과 증류수 2 L를 함께 환류 추 출기에 넣어 탕액이 끓는 시점으로부터 2시간 동안 가열하 여 추출하였다. 그 후 추출액을 filter paper를 이용하여 감압 여과한 후 rotary vacuum evaporator로 농축액을 얻었으며 동 결건조기를 이용해 powder 형태로 제작하였다. 제작된 동결
건조 분말의 무게는 9.4 g로 추출 수율은 18.8%이다.
2.2. 세포 배양
인간 간암 세포인 HepG2는 37
oC, 5% CO
2조건에서 10%
Fetal bovine serum (FBS, WELGENE, Korea)과 1% penicillin, streptomycin (WELGENE, Korea)이 첨가된 Dulbecco's modified Eagle medium (DMEM, WELGENE, Korea)을 이용하여 배 양하였으며 3~4일에 한 번씩 계대 배양하였다.
2.3. 항산화 효능 측정
항산화 효능은 2, 2'-azino-bis-3-ethylbenzothiazoline-6-sulphonic acid (ABTS) 와 2, 2-diphenyl-1-picryl-hydrazyl-hydrate (DPPH) assay 를 이용하여 측정하였다. ABTS assay는 potassium persulfate (SIGMA, USA) 2.4 mM과 ABTS (SIGMA, USA) 7 mM을 1 : 1로 혼합해주고 실온에서 차광 된 상태로 24시간 동안 반응 시켜 ABTS radical 상태로 만들어 주었다. 그 후 ABTS radical 상태를 0.7 부근이 되도록 증류수로 희석하였다. 96 well plate의 각 well에 ABTS working solution 80 μL와 꽃송 이버섯 추출물 농도 별 (0.5, 1, 2 mg/mL) 20 μL씩 넣어주고, 차광시킨 뒤 4분간 반응시키고 microplate reader (Molecular Devices EMax Plus, USA)로 650 nm 파장에서 흡광도를 측 정하였다. 측정값을 이용하여 꽃송이버섯의 항산화 효능을 꽃송이버섯을 처리하지 않은 대조군 대비 백분율로 다음의 식에 따라 항산화 효능을 계산하였다. DPPH assay는 DPPH powder (SIGMA, USA)를 methanol에 용해시켜 0.2 mM의 DPPH radical 상태로 만들어 주었다. 그 후에 96 well plate의 각 well에 DPPH 100 μL와 꽃송이버섯 추출물을 농도 별로 100 μL씩 넣어준 뒤, 차광 된 상태로 30분간 반응시키고 microplate reader (Molecular Devices EMax Plus, USA) 520 nm 파장에서의 흡광도를 측정하였다. 측정값은 ABTS assay 와 동일하게 계산하였다.
ABTS and DPPH radical scavenging activity (%) = {1- (A
Sample– A
Sample blank/ A
Blank)} × 100
2.4. 세포 활성도 측정
꽃송이버섯 추출물이 HepG2 세포 활성도에 미치는 영향을 측 정하기 위해 3-(4, 5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl) -2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay를 수행하였다.
96 well plate에 HepG2 세포를 3,000 cells/well로 분주하여 37
oC, 5% CO
2조건에서 24시간 동안 배양한 뒤 각각의 well 에 농도 별 꽃송이버섯 추출물을 처리하고 36시간 동안 배양 하였다. 그 후 MTS 시약 (Promega, USA) 20 μL을 넣어준 뒤 microplate reader (Molecular Devices EMax Plus, USA)를 이 용하여 490 nm 파장에서 흡광도를 측정하였다. 세포의 생존 율은 다음과 같은 식으로 계산하였다.
세포 활성도 (%) = {(시료첨가군의 흡광도-시료자체의 흡
광도) / 대조군의 흡광도} × 100
2.5. ATP 생성 측정
꽃송이버섯 추출물이 HepG2 세포의 ATP 생성에 미치는 영 향을 확인하기 위하여 ATP bioluminescence assay를 수행하 였다. 96 well plate에 HepG2 세포를 3,000 cells/well로 분주 하여 37
oC, 5% CO
2조건에서 24시간 동안 배양한 뒤 각각의 well에 농도 별 꽃송이버섯 추출물을 처리하고 36시간 동안 배양하였다. 그 후 CellTiter-Glo® 2.0 Assay (Promega, USA) kit의 protocol에 제시된 방법을 이용하여 ATP 생성량을 확인 하였으며 발광 정도는 Luminometer (Thermo Fisher Scientific, USA)로 측정하였다. 측정값을 이용하여 꽃송이버섯 추출물 을 처리하지 않은 대조군을 기준으로 상대적인 ATP 생성량 을 계산하였다.
2.6. 해당 과정 활성 측정
꽃송이버섯 추출물이 HepG2 세포의 해당 과정 활성에 미치 는 영향을 측정하기 위해 해당 과정의 marker인 lactate의 농 도를 측정하였다. 96 well plate에 HepG2 세포를 3,000 cells/
well로 분주하여 37
oC, 5% CO
2조건에서 24시간 동안 배양 한 뒤 각각의 well에 농도 별 꽃송이버섯 추출물을 처리하고 36시간 동안 배양하였다. 그 후 각 배양액 속 lactate의 농도 를 Lactate Pro 2 (Arkray, Japan)을 이용해 측정하였다. 세포 가 포함되어 있지 않은 well에 꽃송이버섯 추출물을 동일한 농도로 처리한 배양액을 이용해 보정한 뒤 상대적인 lactate 농도를 계산하였다.
2.7. 미토콘드리아 활성 측정
꽃송이버섯 추출물이 HepG2 세포의 미토콘드리아 활성에 미치는 영향을 측정하기 위해 미토콘드리아 막 전위를 측정 하는 Tetramethylrhodamine, methyl ester (TMRM) assay를 수 행하였다. HepG2 세포를 6 well plate에 10,000 cells/well로 분주하여 37
oC, 5% CO
2조건에서 24시간 동안 배양한 뒤 각 각의 well에 농도 별 꽃송이버섯 추출물을 처리하고 36시간 동안 배양하였다. 그 후 배양액을 제거한 후에 각 well에 배
양액에 100 nM 농도 TMRM (SIGMA, USA) 용액을 처리한 뒤 37
oC, 5% CO₂ 조건에서 1시간 동안 반응시켰다. 음성 대조군으로 미토콘드리아 uncoupler로서 전자전달계에서 양성자 농도 기울기 형성을 억제하는 carbonyl cyanide m- chlorophenylhydrazone (CCCP, SIGMA, USA)을 사용하였으 며 37
oC, 5% CO
2조건에서 1시간 동안 반응시켰다 [32]. 그 후 배양액을 완전히 제거한 후 각 well 당 PBS 2 mL씩 넣은 뒤 형 광현미경을 통해 미토콘드리아의 활성을 확인하였다.
2.8. 자료 분석 및 통계처리
대조군에 대한 실험군의 유의성을 student's t-test로 계산하여 p-value 값이 0.05 미만일 때 통계적으로 유의한 것으로 판단 하였다.
3. RESULTS AND DISCUSSION
3.1. 꽃송이버섯 추출물의 항산화 효능 확인
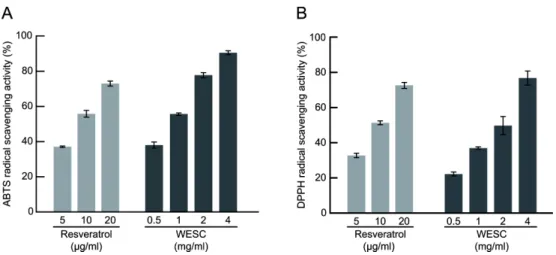
활성 산소종의 과다 생성으로 인한 산화적 스트레스는 당뇨 와 그 합병증의 주요한 원인으로 작용한다 [28]. 따라서 꽃송 이버섯 추출물의 활성 산소종 제거 효능을 ABTS 및 DPPH assay 를 이용하여 측정하였다. 두 실험 방법은 항산화 물질 의 radical 소거 능을 측정하는 원리로 항산화 효능에 대해 잘 알려진 resveratrol을 실험에 대한 신뢰성 확보를 위한 양성 대조군으로 사용하였다 [33]. 꽃송이버섯 추출물은 0.5, 1, 2, 4 mg/mL 농도에서 resveratrol은 25, 50, 100 μM 농도에서 의 존적 ABTS radical 및 DPPH radical 소거 능을 확인하였다.
그 결과 꽃송이버섯 추출물의 농도 의존적 ABTS radical 및 DPPH radical 소거 능이 확인되었다 (Fig. 1(A), Fig. 1(B)). 이 러한 결과는 꽃송이버섯 추출물은 뛰어난 항산화 효능을 가 지고 있어 당뇨병 예방에 긍정적 효능을 가지고 있을 가능성 을 말해준다.
Fig. 1. Antioxidant effects of the water extract of S. crispa (WESC). Resveratrol was tested at the concentration of 5, 10, 20 µg/ml and
WESC was tested at the concentration of 0.5, 1, 2, 4 mg/ml. (A) ABTS radical scavenging activities of resveratrol and WESC. (B)
DPPH radical scavenging activities of resveratrol and WESC. n = 3 (biological replicates).
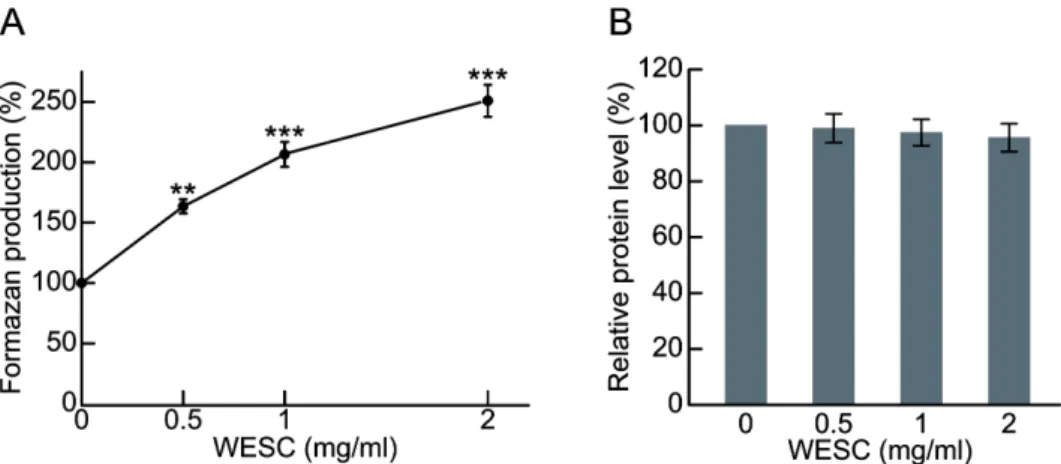
3.2. 꽃송이버섯 추출물에 의한 HepG2 세포 활성도 확인 간은 탄수화물, 지방산, 아미노산 등의 체내 물질대사에 매 우 중요한 기관으로 인슐린 및 기타 호르몬에 의해 복잡한 조절 메커니즘에 의해 조절된다 [34-35]. 간은 글리코겐 합성 (glycogenesis)과 글리코겐 분해 (glycogenolysis)로 혈액의 포 도당 항상성을 조절하며 간 질환 환자의 경우, 인슐린 저항 성, 당 불내성 (glucose intolerance) 및 당뇨병과 같은 질환으 로 인해 에너지 대사의 항상성이 손상된다 [36-37]. 따라서 꽃송이버섯이 간세포 활성에 미치는 영향을 알아보기 위해 꽃송이버섯 추출물 0.5, 1, 2 mg/mL 농도로 인간 간암 유래 HepG2 세포에 처리하여 MTS assay를 수행하였다. 그 결과 흥미롭게도 꽃송이버섯 추출물 2 mg/mL 농도 처리에서 formazan의 생성량이 2.5배 이상 증가함을 확인하였다 (Fig.
2(A)). 이러한 formazan 생성량 증가가 세포 수 증가에 따른 결과인지 확인하기 위해 꽃송이버섯 추출물을 동일한 농도 로 HepG2 세포에 처리한 후 Bradford protein assay를 통해 단 백질의 농도를 측정하였다. 그 결과 꽃송이버섯 추출물의 처 리는 HepG2 세포의 단백질 농도에 유의미한 변화를 주지 않 는다는 걸 확인하였다 (Fig. 2(B)). 또한 cell counting 결과 꽃 송이버섯 추출물의 처리는 세포 수에 유의미한 변화를 주지 않았다 (데이터 제공되지 않음). MTS assay는 살아있는 세포 의 mitochondrial dehydrogenase 등에 의하여 생성되는 NADH, NADPH와 같은 환원 당량 (reducing equivalent)에 의 해 triazolium salt인 MTS가 환원되어 생성되는 formazan의 흡광도를 측정하는 방법으로, 죽은 세포의 경우 tetrazolium 을 환원시키는 능력을 소실하게 된다 [38-39]. 따라서 꽃송이 버섯 추출물 처리에 따른 formazan 생성량의 증가는 세포 증 식 촉진 효능이 아닌 reducing equivalent 생성 촉진 효능을 통해 나타난 것으로 판단하였다.
3.3. 꽃송이버섯 추출물이 HepG2 세포의 ATP 생성에 미 치는 영향 확인
NADH는 해당 작용, TCA 회로 및 산화적 인산화를 포함한
여러 에너지 대사를 통해 생성되어 ATP의 합성을 유도하는 reducing equivalent로 dehydrogenase 및 oxidoreductase 등에 의해 생성된다 [40]. 꽃송이버섯 추출물을 0.5, 1, 2 mg/mL 농 도로 HepG2 세포에 처리할 경우 formazan 생성량이 증가함 을 확인하였다. 이 결과를 통해 우리는 꽃송이버섯 추출물 처리로 인해 HepG2 세포의 reducing equivalent 생성량이 증 가하여 ATP 생성이 촉진될 것으로 판단하였다. 이러한 가능 성을 확인하기 위해 ATP bioluminescence assay를 통해 꽃송 이버섯 추출물 처리에 의한 ATP 생성을 측정하였다. 그 결과 꽃송이버섯 추출물을 처리하지 않은 대조군을 기준으로 2 mg/mL에서 154.63%로 농도 의존적으로 ATP 생성량이 증 가하였다 (Fig. 3). 당뇨병 환자의 간세포에서는 미토콘드리 아의 기능이 저하되어 ATP 생성이 억제되어 있음이 보고되 었으며 마우스 모델 연구를 통해 췌장 베타세포에서 ATP 생 성량 촉진은 insulin의 분비량을 유의미하게 유도한다는 결 과가 보고되었다 [15,17]. 따라서 꽃송이버섯 추출물은 ATP 생성 촉진 효능을 나타냄으로써 제2형 당뇨병 환자의 증상
Fig. 2. The cytotoxicity of the WESC on HepG2 cells. HepG2 cells were treated with indicated concentrations of WESC for 36 h. (A) Cell viability was measured by MTS assay and expressed in percentage. n=3 (biological replicates), **p < 0.01, ***p < 0.001 versus control. (B) Protein concentration was measured by Bradford protein assay. There was no significant difference in protein content by the indicated concentration of WESC.
Fig. 3. ATP level in HepG2 cells treated with WESC. HepG2
cells were treated with indicated concentrations of WESC for 36
h. ATP contents were detected by ATP bioluminescence assay in
the HepG2 cells treated with WESC. ATP levels in HepG2 cells
increased depending on the concentration of the WESC. n = 3
(biological replicates), *P < 0.05 versus control.
3.4. 꽃송이버섯 추출물이 HepG2 세포의 에너지 대사에 미치는 효과 확인
ATP는 세포의 중요한 에너지 분자로써 이온 구배, 생체물질 합성, 소낭 수송과 같은 중요한 세포 기능에 필수적이다
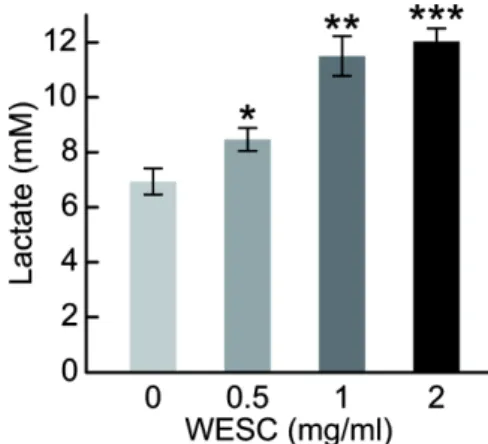
[41]. 해당 과정 및 산화적 인산화는 세포가 ATP를 생성하기 위한 필수적인 에너지 대사과정이다 [42]. 해당 과정은 세포 질에서 일어나며 한 분자의 포도당이 두 분자의 pyruvate로 전환되면서 두 분자의 ATP를 생성한다. 해당 과정의 최종 생 성물인 pyruvate는 산화적 인산화의 초기물질이며 산화적 인 산화는 미토콘드리아 내막을 가로지르는 양성자 농도 기울 기를 형서하고 이는 ATP synthase에 의해 ATP를 합성한다 [42-43]. 또한 해당 과정과 산화적 인산화는 서로 상호전환 시스템 역할을 하여 산화적 인산화가 억제되면 해당 과정은 증가된다 [42,44]. 따라서 꽃송이버섯 추출물 처리에 의한 HepG2 세포의 ATP 생성량의 증가가 어떠한 에너지 대사과정 을 통해 유도되는지 확인하기 위하여 lactate assay와 TMRM assay를 수행하였다. 해당 과정 활성을 확인하기 위해 해당 과정 마커인 lactate의 농도를 측정하였다 [45]. 그 결과 꽃송 이버섯 추출물을 처리하지 않은 대조군에서는 6.93 mM의 lactate 농도가 측정되었으나 꽃송이버섯 추출물을 2 mg/ml 로 처리하였을 때 lactate 농도가 12.03 mM로 꽃송이버섯 추 출물의 농도 의존적으로 lactate 농도가 증가하였다 (Fig. 4).
이러한 결과는 꽃송이버섯 추출물이 HepG2 세포의 해당 과 정 활성에 긍정적 영향을 주고 있음을 보여준다.
꽃송이버섯 추출물에 의한 미토콘드리아 활성 변화를 확 Fig. 4. Lactate level in HepG2 cells treated with WESC. HepG2
cells were treated with indicated concentrations of WESC for 36 h. Lactate concentration was measured by Lactate Pro 2 (Arkray, Japan). n = 3 (biological replicates), *P < 0.05, **p < 0.01,
***p < 0.001 versus control.
Fig. 5. Mitochondrial activity in HepG2 cells treated with WESC. HepG2 cells were treated with indicated concentrations of WESC for
36 h. (A) Fluorescence microscope images of HepG2 cells that were stained with TMRM (100 µM). The CCCP (10 µM) was treated for
negative control. Scale bar = 100 µm. (B) The box graph shows quantification of intensity of TMRM signal. Intensity was quantified by
the ImageJ software. n = 30 (biological replicates), n.s. = not significant, **p < 0.01, ***p < 0.001 versus control.
인하기 위하여 미토콘드리아 막 전위 (Membrane potential) 를 측정하는 TMRM assay를 수행하였다. 그 결과 TMRM 형 광 세기는 0.5 mg/ml에서는 대조군 대비 유의하게 변하지 않 았다. 1 mg/ml에서는 TMRM 형광 세기가 대조군에 비해 유 의한 증가를 나타냈으며 2 mg/ml에서는 1 mg/ml과 유사한 변화가 확인되었다. 이를 통해 꽃송이버섯 추출물 1 mg/ml 이상의 농도에서 미토콘드리아 막전위를 효과적으로 증가 함을 확인하였다 (Fig. 5(A), Fig. 5(B)).
이러한 결과들을 통하여 꽃송이버섯 추출물이 HepG2 세 포의 해당 과정과 미토콘드리아의 활성을 모두 증가시켜 ATP 생성을 촉진함을 확인하였다. 꽃송이버섯 추출물이 HepG2 세포에서 어떠한 작용 기전을 통해 해당 과정과 미토 콘드리아 활성을 증가시키는지 규명하기 위해 추가적인 연 구가 필요할 것으로 판단된다. 이러한 연구 결과들을 통하여 꽃송이버섯 추출물의 당뇨병 예방 및 치료를 위한 천연 기능 성 물질로의 개발이 가능할 것으로 판단된다.
4. CONCLUSION
꽃송이버섯 추출물의 항산화 효능을 확인하기 위해 ABTS와 DPPH assay 를 수행하였으며 그 결과 꽃송이버섯 추출물의 높은 항산화 효능을 확인하였다. 꽃송이버섯 추출물이 HepG2 세포의 활성도에 미치는 영향을 확인하기 위한 MTS assay를 통해 HepG2 세포의 reducing equivalent이 생성됨에 따라 생성되는 formazan 양이 꽃송이버섯 추출물 농도 의존 적으로 증가함을 확인하였다. 또한 꽃송이버섯 추출물은 HepG2 세포에서 농도 의존적으로 ATP 생성량을 증가시켰 으며, 해당 과정 산물인 lactate의 양적 증가를 유도할 뿐만 아니라 미토콘드리아의 막 전위를 유의적으로 촉진함을 확 인하였다. 꽃송이버섯에 의한 당뇨병 개선 효과가 어떠한 물 질에 의해 나타나는지 아직 규명되지 않았으며, 추후 성분분 석을 통해 본 연구에서 사용한 꽃송이버섯 열수 추출물에 포 함된 약리 물질을 분석한 뒤 약리 효능을 나타내는 단일 물 질을 규명할 예정이다. 따라서 본 연구 수행을 통하여 꽃송 이버섯 추출물은 산화적 스트레스 억제 효능 및 간세포의 에 너지 대사를 촉진함으로써 당뇨병 개선을 위한 천연 치료제 및 기능성 식품으로 개발할 수 있는 가능성이 확인되었다.
Acknowledgements
이 논문은 충남대학교 국립대학육성사업 (2018-2019) 지원 을 받아 작성되었음.
REFERENCES