Lipopolysaccharide로 유도된 RAW 264.7 세포에서 유색미 에탄올 추출물의 항염증 작용
최은영․장영아․이진태
대구한의대학교 바이오산업융합학부 화장품약리학과
The Anti-Inflammatory Effect of Ethanol Extracts of Colored Rice Varieties against Lipopolysaccharide-Stimulated RAW 264.7 Cells
Eun-Young Choi, Young-Ah Jang, and Jin-Tae Lee
Department of Cosmeceutical Science, Division of Bio-Technology and Convergence, Daegu Haany University
ABSTRACT This study was conducted to investigate the anti-inflammatory effects of ethanol extracts of Josaengheugchal (JE) and Shintoheug rice (SRE). To accomplish this, the expression of nitric oxide (NO), inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) in lipopolysaccharide (LPS)-stimulated RAW 264.7 macro- phage cells was inhibited. To determine if the extracts were strong enough to inhibit NO production, cell viability was measured. Cells were not affected by JE or SRE, as indicated by a survival rate of 90% or more at a concentration of less than 50 μg/mL. The expression of iNOS and COX-2, which was expressed by inflammatory response when the extracts were treated with LPS-stimulated RAW 264.7 cells, was analyzed by western blotting and reverse tran- scription-polymerase chain reaction (RT-PCR). The results showed that NO, iNOS, and COX-2 were suppressed in a concentration-dependent manner in response to JE and SRE, indicating that these compounds have the potential for use as functional ingredients to inhibit inflammation by the cosmetic and food industries.
Key words: anti-inflammatory, colored rice, COX-2, iNOS, NO
Received 3 July 2018; Accepted 24 August 2018
Corresponding author: Eun-Young Choi, Department of Cosmeceut- ical Science, Daegu Haany University, Gyeongsan, Gyeongbuk 38578, Korea
E-mail: [email protected], Phone: +82-53-819-7745
서 론
우리나라에서 유통되고 있는 유색미는 흑미와 적미, 흑향 미가 대표적이며(1), 흑자색의 현미는 안토시아닌이 풍부하 고 적갈색의 흑미는 탄닌이 포함된 것으로 알려져 있으며 흑향미는 적미와 색이 유사하나 향이 강한 것이 특징이다(2).
유색미는 일반미 품종에 비해서 cyanidin 3-O-β-D-glu- coside 및 peonidin 3-O-β-D-glucoside 등(3,4)의 항산 화 성분이 보고되었고, 지금까지 유색미에 대한 연구를 살펴 보면 유색미 에탄올 추출물(5) 및 색소성분(6)의 항산화 활 성 및 항암 활성이 보고되었다. 유색미 겨 추출물에서 항산 화 활성 및 활성산소종에 대한 소거 활성, DNA쇄의 산화적 손상에 대한 억제 활성 등이 일반 취반용 쌀보다 전반적으로 우수하다는 보고가 있었고(5), 비만세포주인 RBL-2H3 세 포주를 사용하여 알레르기 염증 발생에 대한 억제효과를 검 토한 결과에서도 유색미가 염증 유발에 중요한 인자로 작용 하는 histamine과 matrix metalloproteinase(MMP)의 분
비를 저해하는 능력이 일반미 품종들보다 우수하다는 사실 을 보여주었다(7). 특히 오늘날 현대인들은 식생활의 변화로 인해 만성질병 및 성인병이 사회적인 문제로 대두됨에 따라 육류보다는 건강한 삶을 위한 웰빙(well-being) 및 기능성 식품에 대한 관심이 높아지면서 주식으로 기존의 백미에서 유색미로 대체하여 섭취하는 경향이 증가하고 있다(8). 항산 화 활성이 우수하다고 알려진 유색미 중에 조생흑찰과 신토 흑미는 안토시아닌 고함유 흑자색 찰벼, 흑색메벼 품종이다 (9). 지금까지 유색미에 대한 과학적인 효능 검증이 다양하 게 이루어지고 있으나 아직 조생흑찰과 신토흑미의 염증억 제 활성에 대한 연구는 보고된 바가 없다.
따라서 본 연구에서는 조생흑찰과 신토흑미의 에탄올 추 출물을 이용하여 macrophage cell인 RAW 264.7 세포에서 의 염증 발생에 대한 억제효과를 조사하여 식품 및 화장품의 기능성 원료로 활용하고자 한다.
재료 및 방법
시료의 추출
본 실험에 사용된 선별된 유색미는 조생흑찰(밀량194호) 과 신토흑미(전북3호)의 2종으로 2015년 국립식량과학원 (Miryang, Korea)에서 제공받아 에탄올을 이용하여 추출을
실시하였다. 시료 중량의 10배 양의 70% 에탄올에 침지하 여 상온에서 24시간 방치하여 상등액과 침전물을 분리하여 3회 반복 추출하였다. 각 추출물은 여과지(Whatman No.2, Sigma-Aldrich Co., St. Louis, MO, USA)를 이용하여 여 과한 후 농축・동결건조 하여 추출물을 얻었으며 각각의 수 율은 조생흑찰이 1.71%, 신토흑미가 1.5%였다. 추출이 완 료된 유색미는 냉장 보관하면서 실험에 사용되었다.
세포의 배양
Mouse 유래 macrophage cell line인 RAW 264.7 세포 는 한국세포주은행(Seoul, Korea)으로부터 분양받았으며, 10% fetal bovine serum(FBS)과 penicillin/streptomycin (Sigma-Aldrich Co.) 100 unit/mL가 함유된 Dulbecco’s modified Eagle’s medium(DMEM, Gibco BRL Co., Grand Island, NY, USA) 배지를 사용하여 37°C, 5% CO2 incuba- tor(MCO-18AIC, SANYO Co., Sakata, Japan)에서 배양 하였으며, 2~3일에 한 번씩 계대 배양을 시행하였다.
세포 생존율
세포 생존율 측정은 Mosmann(10)의 방법에 따라 측정하 였다. 세포를 96-well plate에 2×104 cells/well이 되게 0.18 mL 분주하고, 시료를 농도별로 조제한 후 0.02 mL 첨가 하여 5% CO2 incubator에서 24시간 배양하였다. 여기에 5 mg/mL 농도로 제조한 3-(4,5-dimethylthiazol-2-yl)- 2,5-diphenyltetrazolium bromide(MTT, Sigma-Aldrich Co.) 용액 0.02 mL를 첨가하여 4시간 배양한 후 상층액을 제거하고, 형성된 formazan에 각 well당 0.1 mL의 DMSO 용액을 가한 후 ELISA reader(PowerWave XS, BioTek Instruments, Inc., Winooski, VT, USA)로 540 nm에서 흡 광도를 측정하였다.
Nitric oxide(NO) 저해능 측정
RAW 264.7 세포에서 생성되는 NO의 양은 Green 등(11) 의 방법에 따라 측정하였다. 세포를 6-well plate에 1×105 cells/mL가 되게 seeding 한 후 24시간 동안 배양하여 confluence가 80%일 때 PBS로 2번 세척한 다음, 무혈청 배지를 사용하여 24시간 동안 배양한 후 시료를 농도별로 처리하고 나서 4시간 후에 lipopolysaccharide(LPS, Sig- ma-Aldrich Co.) 1 μg/mL를 대조군(Nor)을 뺀 모든 well 에 넣어서 4시간 동안 자극시킨 다음 상층액을 모아 동량의 Griess reagent로 10분간 반응시킨 후 NO의 생성량은 ELISA reader를 이용하여 540 nm에서 흡광도를 측정하였 다.
Western blot을 통한 단백질의 발현 측정
Inducible nitric oxide synthase(iNOS)와 cyclooxyge- nase-2(COX-2)의 발현을 보기 위하여 RAW 264.7 세포를 96-well culture plate에 1×106 cells/mL로 cell seeding
후 24시간 동안 배양하여 세포를 안정화시켰다. 배지를 제 거한 후 시료를 농도별로 처리하고 30분간 반응시킨 다음 LPS를 1 μg/mL 농도로 처리하고 24시간 동안 배양한 후에 상등액을 제거하고 PBS로 2번 세척하였다. Cell을 harvest 하여 Radio-immuno-precipitation assay(RIPA) lysis buffer 200 μL에 세포를 용출시켜 단백질을 수확・원심분리 하여 얻은 상층액은 Bradford assay로 정량하고 SDS- PAGE에서 전기영동 하여 분리하였다. Gel을 3시간 동안 polyvinylidenedifluoride(PVDF) membrane(Sigma-Al- drich Co.)에 옮긴 후 4°C에서 4시간 동안 5% skim milk로 blocking 하였다. Primary antibody(Santa Cruz Biotech- nology, Inc., Dallas, TX, USA)를 1:1,000으로 희석하여 4°C에서 overnight 하고 나서 TBST로 30분간 교반하여 세척하는 과정을 3회 반복하였다. 각각의 HRP-conjugated secondary antibody(Santa Cruz Biotechnology, Inc.)를 희석하여 상온에서 1시간 동안 처리하고 다시 TBST로 10 분간 교반하여 세척하는 과정을 3회 반복한 후 ECL 용액을 가한 다음 Western imaging system(EZ-Capture MG, ATTO, Amherst, NY, USA) 기기를 이용하여 밴드 확인 및 정량하였다.
RNA 추출 및 reverse transcription-polymerase chain reaction(RT-PCR) 과정
RNA는 High Pure RNA Isolation Kit(Roche, Basel, Switzerland)을 이용하였고 제조회사의 방법에 준하여 to- tal RNA를 추출하였으며, 1 μg/μL의 농도로 정량하고 추출 된 total RNA는 Transcriptor First Strand cDNA Syn- thesis Kit(Roche)을 이용하여 역전사를 진행시켜 cDNA를 합성시켰다. iNOS, COX-2의 mRNA 발현을 알아보기 위하 여 Fast start Essential DNA Green Master kit(Roche)을 사용하여 RT-PCR을 수행하였다. 이 분석에서 internal control은 GAPDH(Santa Cruz Biotechnology, Inc.)를 사 용하였으며 primers의 sequence는 Table 1과 같다.
PCR의 증폭 조건은 다음과 같다. iNOS는 95°C에서 60초 간 30 cycle을 증폭시킨 후 60°C에서 60초간, 72°C에서 65초간 실행시켰다. COX-2는 94°C에서 60초간 30 cycle 을 증폭시킨 후 60°C에서 60초간, 72°C에서 65초간 실행시 켰다.
RT-PCR 증폭으로 생산된 DNA는 0.5 μg/mL의 ethi- dium bromide가 포함된 1.5% metaphor agarose gel에 전기영동 하여 cooled CCD camera system EZ-Capture
Ⅱ와 CA analyzer ver. 3.00 software(ATTO)를 이용하여 mRNA의 발현 정도를 확인하였다.
통계처리
실험 결과에 대한 통계처리는 SPSS 23.0(IBM SPSS Inc., New York, NY, USA)을 이용하여 평균과 표준편차로 나타내었고, 각 처리군 간의 유의성에 대한 검증은 분산분석
0 20 40 60 80 100
Cont 12.5 25 50 100
Concentration (µg/mL)
Cell viability (%) .
A JE
** **
0
20 40 60 80 100
Cont 12.5 25 50 100
Concentration (µg/mL)
Cell viability (%) .
SRE
*
B
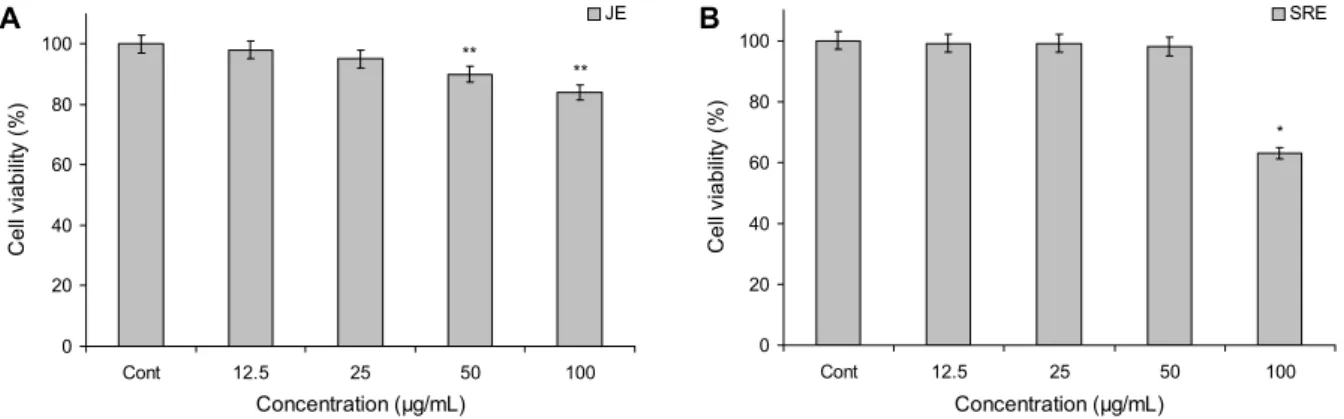
Fig. 1. Cell viability assay on macrophage RAW 264.7 cells of colored rice extracts. The cells were treated for 24 h with the indicated concentrations of colored rice extracts. Cont: not treated any extract. (A) JE: 70% ethanol extract of Josaengheugchal, (B) SRE:
70% ethanol extract of Shintoheug rice. Results are mean±SD of triplicate data. One-way ANOVA was used for comparisons of multiple group means followed by t-test (significant as compared to control; *P<0.05, **P<0.01).
Table 1. Sequence of the primers used for PCR
Gene Primer Sequence (5’→3’)
iNOS COX-2 GAPDH
Sense Anti-sense
Sense Anti-sense
Sense Anti-sense
5’-ATTCGCAACATCAGGTCGGCCATCACT-3’
5’-GCTGTGTGTCACAGAACTCTCGAACTC-3’
5’-GGAGAGACTATCAAGATACT-3’
5’-ATGGTCAGTAGACTTTTACA-3’
5’-ACCACAGTCCATGCCATCAC-3’
5’-TCCACCACCCTGTTGCTGTA-3’
(analysis of variance, ANOVA)을 이용하여 유의성(P<
0.05, 0.01, 0.001)을 확인한 후 Tukey’s test로 다중비교 를 실시하여 분석하였다.
결과 및 고찰
세포 생존율 측정 결과
Yellow tetrazolium salt MTT는 담황색 기질로서 살아 있는 세포의 미토콘드리아 내의 reductase에 의해 환원되 어 formazan을 생성하는데, 죽어있는 세포에서는 형성되지 않고 살아있는 세포의 수가 많을수록 formazan의 생성도 많아지고 세포의 성장을 측정할 수 있다(12).
선별된 유색미 2종인 조생흑찰 에탄올 추출물(JE)과 신토 흑미 에탄올 추출물(SRE)의 mouse 유래 macrophage cell 인 RAW 264.7 세포에서의 생존율을 확인하기 위하여 MTT assay를 실시하였으며, 그 결과는 Fig. 1에 나타내었다.
RAW 264.7 세포에 대하여 JE와 SRE 모두 50 μg/mL 이하 의 농도에서 각각 90%와 98% 이상의 세포 생존율을 나타내 었다. 이러한 결과를 바탕으로 세포의 NO 소거능 실험은 추출물 자체의 세포독성으로 인한 지시세포 사멸의 가능성 을 배제할 수 있도록 50 μg/mL의 농도까지 처치하였다.
NO 저해 활성 측정 결과
체내에 과도하게 생성된 reactive oxygen species(ROS) 는 세포들의 산화적 손상을 유발할 뿐만 아니라 염증반응의
신호 전달계와 연결되어 암, 치매, 당뇨병, 류마티스 관절염 과 같은 퇴행성 질환이나 노화를 촉진한다고 알려져 있다 (13). 활성산소종의 일종인 NO는 대식세포의 항균 활성 및 세포의 신호전달에 중요한 역할을 하지만, 생체에서 생성된 superoxide radical과 반응하여 세포독성이 더욱 강한 re- active peroxynitrite와 같은 이차적인 라디칼을 생성시킨 다고 보고되어 있다(14). 생체 내 고농도의 NO 생성은 숙주 세포의 파괴와 shock에 의한 혈관 확장, 염증 반응 유발에 의한 조직의 상해를 초래할 수 있는 이중적인 생물학적인 성질을 가진다(15).
LPS의 자극에 의해 발현된 iNOS는 많은 양의 NO를 생성 하게 되며, 이에 의한 세포 독성은 염증 반응, 세포의 돌연변 이 및 종양 발생 등에도 관여하므로 염증반응과 관련된 조직 손상에 NO와 iNOS의 발현이 증가함이 보고되어 있다(16).
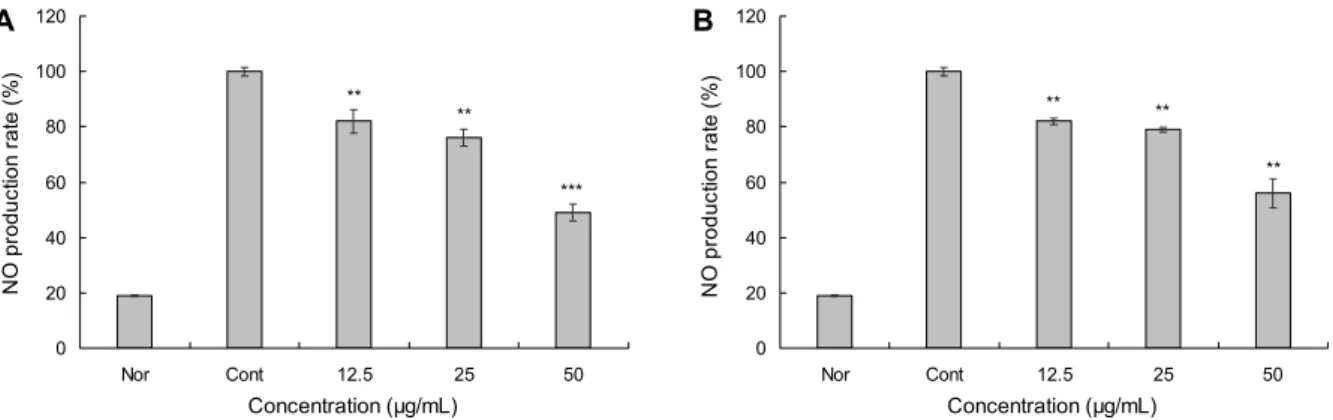
우리는 RAW 264.7 세포에 세균 유래의 LPS를 처리하여 자극하였을 때 유색미 에탄올 추출물의 첨가로 NO의 생성 을 저해하는지 알아보고자 하였으며, 그 결과는 Fig. 2에 나타내었다. LPS 단독 처리구(Cont) 기준으로 LPS와 함께 JE와 SRE를 처리하였을 때 두 추출물 모두 농도 의존적으로 NO의 생성량이 유의성 있게 감소하였으며, JE와 SRE는 50 μg/mL의 농도에서 51%와 44%의 NO 저해 효과를 보였다.
Choi 등(7)이 연구한 NO 소거 활성 결과 유색미 추출물 가 운데 LK1A-2-12-1-1이 100 μg/mL의 농도에서 20.8%
의 저해효과를 나타내었고, 일반미인 추청은 28.2%의 저해 효과를 보였다고 보고하였다. 이는 본 연구 결과에서 보여준
***
**
**
0 20 40 60 80 100 120
Nor Cont 12.5 25 50
Concentration (μg/mL)
NO production rate (%) .
A
**
** **
0 20 40 60 80 100 120
Nor Cont 12.5 25 50
Concentration (μg/mL)
NO production rate (%) .
B
Fig. 2. Nitric oxide (NO) inhibition assay on macrophage RAW 264.7 cells of colored rice extracts. The cells were treated for 24 h with the indicated concentrations of colored rice extracts. Nor: not treated any extract and LPS, Cont: only treated LPS.
(A) JE: 70% ethanol extract of Josaengheugchal, (B) SRE: 70% ethanol extract of Shintoheug rice. Results are mean±SD of triplicate data. One-way ANOVA was used for comparisons of multiple group means followed by t-test (significant as compared to control;
**P<0.01, ***P<0.001).
추출물의 소거효능이 훨씬 우수함을 나타낸 결과이다.
iNOS 및 COX-2 단백질 발현 억제 결과
염증 반응은 생체나 조직에 물리적 작용이나 화학적 물 질, 세균 감염 등의 어떠한 기질적인 변화를 가져오는 침습 이 가해질 때 그 손상 부위를 수복하고 재생하려는 기전이다 (17). 자극이 가해지면 국소적으로 염증성 성분과 같은 혈관 활성 물질이 유리되어 혈관 투과성이 증대되면서 염증을 유 발하며, 지속적인 염증반응은 점막 손상을 촉진하고 그 결과 여러 질환을 일부 발생시킬 수 있다(18). 염증반응 시에는 NO와 cytokine을 생산하여 감염 초기에 생체 방어에 중요 한 역할을 한다(19). 포유동물 세포의 nitric oxide syn- thase(NOS)의 경우 유사형태가 3가지 존재하는데 neuro- nal NOS(nNOS), endothelial NOS(eNOS), 그리고 iNOS이 다. 그중에서 특히 iNOS가 염증반응에 관여한다. nNOS와 eNOS는 항상 발현되어 있으며, iNOS의 경우 평소에는 존 재하지 않으나 interferon-γ, LPS, 그리고 여러 가지 염증 사이토카인의 자극이 있을 때 발현된다. 일단 유도되면 장시 간 동안 다량의 NO를 생성하게 되며 이로 인해 혈관 투과성 과 부종 등의 염증반응을 촉진시키고 염증 매개체의 생합성 을 촉진하여 염증을 심화시키는 것으로 알려져 있다(20).
Cyclooxygenase(COX)에 대해서는 1990년대 초반에 주로 연구되었는데, COX는 cyclooxygenase와 peroxidase 활 성을 모두 가지고 있는 효소이다. Cyclooxygenase 기능으 로서 arachidonic acid를 prostaglandin으로 변환하고, peroxidase 기능으로서는 endoperoxide를 prostaglandin 으로 변환시키며, prostaglandin은 prostaglandins, throm- boxane 및 prostacyclins의 전구체로 사용된다(21). 이 또 한 유사형태가 2가지 존재하는데, COX-1은 정상세포의 항 상성을 유지하기 위하여 거의 모든 조직에 발현되어 있고, prostaglandin을 생산하여 신장의 혈액 흐름을 조절하거나 위장의 세포를 보호하는 등의 생리적인 기능을 조절한다.
반대로 COX-2의 경우는 미생물에 의한 감염이나 손상 혹은 여러 요인의 스트레스에 반응한 대식세포에서 발현된다. 특 히 급성염증반응에서 prostaglandins의 합성에 관여하고, LPS 및 cytokine에 의해 발현이 유도된다(22). 즉 iNOS와 COX-2의 발현과 NO, prostaglandin E2(PGE2)는 면역세포 의 대표적인 염증인자이다(21). 본 실험에서는 mouse 유래 의 macrophage cell인 RAW 264.7에 LPS를 1 μg/mL 농도 로 처치한 후 유색미 에탄올 추출물을 농도별로 주입했을 때의 iNOS와 COX-2 발현의 억제 정도를 western blot을 통해 확인하였으며, 결과는 Fig. 3과 같이 나타났다. 염증 발현 인자인 iNOS와 COX-2의 단백질 발현은 농도 의존적 으로 억제되었음을 알 수 있었고, 특히 50 μg/mL의 농도에 서 iNOS와 COX-2가 JE의 경우 71%, 69%의 억제효과를 나타내었으며, SRE는 52%, 75%가 각각 억제되었다. Choi 등(7)은 서울대학교 농학과에서 분양받은 Elwee와 LK1- 3-6-12-1-1 등의 유색미에 대한 염증유발의 지표로서 MMP 생성 정도를 확인하였으며, 그 결과 MMP-2는 10 μg/
mL에서 LK1-3-6-12-1-1이 약 36%의 억제효과를 보인 것으로 보고하였고, MMP-9에 대한 저해활성 결과에서는 LK1-3-6-12-1-1과 Elwee가 완전히 억제한 결과를 나타 내었다고 보고하였다. 그러나 아직 우리나라에서 생산되는 유색미의 iNOS와 COX-2에 대한 발현억제 연구는 보고된 바가 없어 향후 연구의 중요한 기준이 될 수 있을 것으로 기대된다.
iNOS 및 COX-2의 mRNA 발현 억제 결과
염증 반응이 일어나면 염증매개물질인 NO, PGE2, 염증 성 cytokine 등이 분비된다. 그중 염증 반응의 지표물질인 NO는 L-arginine에서 NOS에 의해 합성되며, NOS의 종류 가운데 iNOS에 의한 NO 형성은 병리학적으로 중요한 역할 을 한다(23). 다른 염증인자 중 하나인 COX는 COX-1과 COX-2가 존재하며 이는 다양한 세포에서 다른 경향을 나타
iNOS COX-2 α-tubulin
iNOS COX-2 α-tubulin
A B
0 20 40 60 80 100 120
Nor Cont 12.5 25 50
Concentration (μg/mL)
Protein expression rate (%) .
iNOS COX-2
**
**
** **
0 20 40 60 80 100 120
Nor Cont 12.5 25 50
Concentration (μg/mL)
Protein expression rate (%) .
iNOS COX-2
*
**
**
**
Fig. 3. iNOS and COX-2 protein expression in LPS-stimulated RAW 264.7 cells of colored rice extracts. The cells were treated for 24 h with the indicated concentrations of colored rice extracts. The results were analyzed by western blot. Nor: not treated any extract and LPS, Cont: only treated LPS. (A) JE: 70% ethanol extract of Josaengheugchal, (B) SRE: 70% ethanol extract of Shintoheug rice. Results are mean±SD of triplicate data. One-way ANOVA was used for comparisons of multiple group means followed by t-test (significant as compared to control; *P<0.05, **P<0.01).
A B
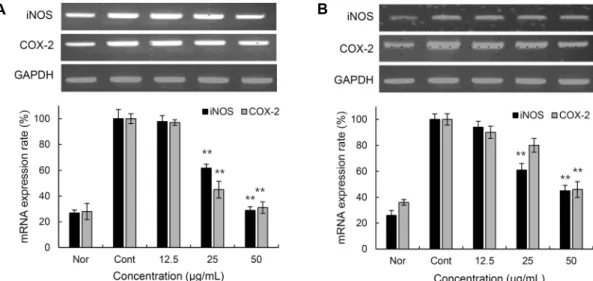
Fig. 4. iNOS and COX-2 mRNA suppression in LPS-stimulated RAW 264.7 cells of colored rice extracts. The cells were treated for 24 h with the indicated concentrations of colored rice extracts. The results were analyzed by RT-PCR. Nor: not treated any extract and LPS, Cont: only treated LPS. (A) JE: 70% ethanol extract of Josaengheugchal, (B) SRE: 70% ethanol extract of Shintoheug rice. Results are mean±SD of triplicate data. One-way ANOVA was used for comparisons of multiple group means followed by t-test (significant as compared to control; **P<0.01).
낸다. 이 중 COX-2는 염증반응 부위에서 발현이 된다. COX- 2에 의해 생성되는 PGE2는 통증, 발열 등에 관여하는 염증 매개체로서 염증반응, 면역반응에 관여하고 혈관신생을 촉 진시키는 것에 깊이 관여하고 있는 것으로 알려져 있다(24).
우리는 유색미 에탄올 추출물의 iNOS와 COX-2의 mRNA 발현억제를 확인하기 위한 RT-PCR을 실시하였으며, 이때 세포의 여러 조건에서도 그 발현 정도의 차이가 거의 없는 house keeping gene인 GAPDH를 positive control로 사용 하였다. 그 결과는 Fig. 4와 같이 JE는 20 μg/mL의 농도에
서 iNOS와 COX-2의 mRNA가 각각 71%, 69%의 발현을 억제시켰으며, SRE는 같은 농도에서 각각 55%, 54%의 발 현을 억제시켰다. 또한 두 추출물 모두 농도 의존적인 억제 효능을 나타내었다.
요 약
본 연구에서는 선별된 유색미 2품종에 대한 LPS에 유도된 RAW 264.7 세포주에서 발현되는 염증인자인 NO 및 iNOS
와 COX-2의 억제효과를 확인하고자 하였다. 유색미 에탄 올 추출물인 조생흑찰 에탄올 추출물과 신토흑미 에탄올 추 출물 모두 50 μg/mL 이하의 농도에서 RAW 264.7 대식세 포에 대하여 90% 이상의 세포 생존율을 나타내었으며, LPS 에 유도된 RAW 264.7 세포주의 NO 발현, iNOS와 COX-2 의 단백질 발현 및 mRNA 발현이 농도 의존적으로 매우 유 의하게 억제되었음을 확인하였다. 우리는 선별된 유색미 2 종의 에탄올 추출물이 염증 관련 인자들의 생성을 억제 또는 조절한다는 결과를 확인함에 따라 식품・화장품 분야에 유용 한 항염증 기능성 소재로써 활용될 수 있을 것으로 생각된 다.
감사의 글
본 연구는 농림축산식품부에서 지원하는 2015년도 농생명 산업기술개발사업(과제번호: 15-162-004)의 연구수행으 로 진행되었습니다.
REFERENCES
1. Nam SH, Chang SM, Kang MY. 2003. Varietal difference in antioxidative activity of ethanolic extracts from colored rice bran. J Korean Soc Agric Chem Biotechnol 46: 16-22.
2. Choi HC, Oh SK. 1996. Diversity and function of pigments in colored rice. Korean J Corp Sci 41: 1-9.
3. Choi SW, Kang WW, Osawa T. 1994. Isolation and identi- fication of anthocyanin pigments in black rice. Food Bio- technol 3: 131-135.
4. Tsuda T, Watanabe M, Ohshima K, Norinobu S, Choi SW, Kawakishi S, Osawa T. 1994. Antioxidative activity of the anthocyanin pigments cyanidin 3-O-β-D-glucoside and cya- nidin. J Agric Food Chem 42: 2407-2411.
5. Nam SH, Kang MY. 1997. In vitro inhibitory effect of colored rice bran extracts carcinogenicity. Agric Chem Biotechnol 40: 307-312.
6. Choi SW, Nam SH, Choi HC. 1996. Antioxidative activity of ethanolic extracts of rice brans. Food Biotechnol 5: 305- 309.
7. Choi SP, Kang MY, Nam SH. 2004. Inhibitory activity of the extracts from the pigmented rice brans on inflammatory reactions. J Korean Soc Appl Biol Chem 47: 222-227.
8. Ryu SN, Park SZ, Kang SS. 2005. Studies on exploration and expansive use of genetic variation of functional sub- stances in rice. Report of Rural Development Administra- tion, Suwon, Korea.
9. Park JY, Ham HM, Han SI, Oh SH, Song YC, Cho JH, Hur YJ, Lee YY, Lee BW, Choi YH. 2016. Comparison
of antioxidant components and antioxidant activities of col- ored rice varieties cultivated in southern plain. J Korean Soc Food Sci Nutr 45: 1214-1220.
10. Mosmann T. 1983. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cyto- toxicity assays. J Immunol Methods 65: 55-63.
11. Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SR. 1982. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal Biochem 126: 131- 138.
12. Gross J, Lapiere CM. 1962. Collagenolytic activity in am- phibian tissues: a tissue culture assay. Proc Natl Acad Sci 54: 1197-1204.
13. Li C, Wang MH. 2011. Antioxidant activity of peach blos- som extracts. J Korean Soc Appl Biol Chem 54: 46-53.
14. Pryor WA, Squadrito GL. 1995. The chemistry of peroxyni- trite: a product from the reaction of nitric oxide with super- oxide. Am J Physiol 68: L699-L722.
15. Kawamata H, Ochiai H, Mantani N, Terasawa K. 2000.
Enhanced expression of inducible nitric oxide synthase by Juzen-taiho-to in LPS-activated RAW264.7 cells, a murine macrophage cell line. Am J Chin Med 28: 217-226.
16. Mori M. 2007. Regulation of nitric oxide synthesis and apop- tosis by arginase and arginine recycling. J Nutr 137: 1616S- 1620S.
17. Tizard IR. 2004. Veterinary immunology: an introduction.
9th ed. WB Saunders Company, Philadelphia, PA, USA. p 480.
18. Wiloughby DA. 1975. Heberden oration, 1974. Human arthri- tis applied to animal models. Towards a better therapy. Ann Rheum Dis 34: 471-478.
19. Higuchi M, Higashi N, Taki H, Osawa T. 1990. Cytolytic mechanisms of activated macrophages. Tumor necrosis fac- tor and L-arginine-dependent mechanisms act synergistically as the major cytolytic mechanisms of activated macrophages.
J Immunol 144: 1425-1431.
20. Kim RG, Shin KM, Chun SK, Ji SY, Seo SH, Park HJ, Choi JW, Lee KT. 2002. In vitro antiinflammatory activity of the essential oil from Ligularia fischeri var. spiciformis in mur- ine macrophage Raw 264.7 cells. Yakhak Hoeji 46: 343-347.
21. Yun HJ, Heo SK, Yi HS, Kim CH, Kim BW, Park SD. 2008.
Anti-inflammatory effect of injinho-tang in RAW264.7 cells.
Kor J Herbol 23: 169-178.
22. Hume DA, Wells CA, Ravasi T. 2007. Transcriptional regu- latory networks in macrophages. Novartis Found Symp 281:
2-18.
23. Nathan C. 1992. Nitric oxide as a secretory product of mam- malian cells. FASEB J 6: 3051-3064.
24. Masferrer JL, Zweifel BS, Manning PT, Hauser SD, Leahy KM, Smith WG, Isakson PC, Seibert K. 1994. Selective inhibition of inducible cyclooxygenase 2 in vivo is antiin- flammatory and nonulcerogenic. Proc Natl Acad Sci U S A 91: 3228-3232.