익모초 에탄올 추출물의 항산화 및 항염증 활성*
1)
최유나
***․최유경
****․난리
*****․추병길
**Anti-oxidant and Anti-inflammatory Effects of Ethanol Extracts from Leonurus japonicus Houtt. on LPS-induced RAW 264.7 Cells
Choi, You-Na․Choi, Yu-Kyung․Nan, Li․Choo, Byung-Kil
Leonurus japonicus (L. japonicus) Houtt., a biennial plant in the Lamiaceae family is broadly distributed in Asia such as Korea, China, Japan. The aerial part of L.
japonicus is used as a traditional medicine to treat uterine disease including dysmenorrhea, amenorrhea, sterility. In this study, we examined the antioxidant and anti-inflammatory effects of L. japonicus ethanol extracts. The antioxidant activity of L. japonicus was measured by total polyphenol and flavonoid content, and DPPH, ABTS scavenging, reducing power activity, and intracellular ROS express- ion assay. The anti-inflammatory effects were measured by nitric oxide (NO), cytokines (TNF- α and IL-1β) production and inflammatory protein expression in LPS-induced RAW 264.7 cells. Total polyphenol and flavonoid content of L.
japonicus were 51.40 ± 0.47 mg of gallic acid equivalents/g and 73.28 ± 0.10 mg of rutin equivalents/g respectively. DPPH, ABTS radical scavenging activity and reducing power activity tended to increase concentration-dependent and treatment L. japonicus with 400 µg/mL reduced ROS production by 69.5%. Furthermore, L.
japonicus inhibited NO, TNF-α and IL-1β production in a concentration-depen- dant manner and reduced the expression levels of inflammatory proteins via regulating NF-ĸB, MAPK pathway. Therefore, we suggest that L. japonicus could be a natural antioxidants and medicinal source to treat oxidative stress and inflam- mation-related disease.
Key words : anti-inflammatory, antioxidant, Leonurus japonicus, MAPK, NF-ĸB
*****
본 논문은 농촌진흥청 연구사업(세부과제번호: PJ0142272020)의 지원에 의해 이루어진 것임.
*****
Corresponding author, 전북대학교 농업생명과학대학 작물생명과학과 교수([email protected])
*****
First author, 전북대학교 농업생명과학대학 농학과 석사과정
*****
전북대학교 농업생명과학대학 작물생명과학과 학부생
*****
전북대학교 농업생명과학대학 작물생명과학과 박사
Ⅰ. 서 론
신체는 에너지 생산을 위하여 호흡과정을 통해 들어온 산소를 이용해 여러 대사활동을 하 는데, 대사과정 중에 산소가 불완전하게 환원되면 활성이 높은 중간산물인 reactive oxygen species (ROS)이 생성된다(Lee et al., 2015). 활성산소는 대사과정뿐만 아니라 환경오염물질, 과도한 스트레스, 흡연 등의 외부적 요인에 의해서도 발생하는데 인체는 이들을 제거할 수 있는 superoxide dismutase, catalase 등의 효소를 통한 항산화 시스템을 구축하고 있지만, 과 량의 활성산소 생성은 세포 내의 지질 과산화, DNA 손상 등을 통해 암, 골다공증, 심장질 환, 호흡기질환과 같은 다양한 문제들을 야기한다(Chapple, 1997; Birben et al., 2012). 이와 같이 활성산소로 인한 질병의 예방과 치료를 위해 butylated hydroxy anisol (BHA), butylated hydroxy toluene (BHT)과 같은 합성 항산화제들이 사용되고 있으나, 다양한 부작용 발생으 로 안정성 문제가 지속적으로 제기되고 있다(Anand and Sati, 2013).
염증은 감염과 같은 외부 자극에 대한 신체의 정상적인 방어반응으로 일정량의 염증반 응은 감염상태를 회복시키지만, 과도하게 지속될 경우 세포나 조직에 손상을 일으켜 자가 면역질환, 패혈증, 암 등을 유발시킨다(Dinarello, 2010). 염증반응은 대식세포 및 면역세포 의 활성으로 시작되며, 염증반응을 유도하는 lipopolysaccharide (LPS)로 자극된 대식세포는 mitogen-activated protein kinases (MAPKs) 및 nuclear factor kappa B (NF-ĸB) 전사인자를 활 성화시켜 nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), tumor necrosis factor alpha (TNF-α), interleukin 1 beta (IL-1β)와 같은 염증 매개체 발현을 촉진시킨다. iNOS와 COX-2 는 각각 nitric oxide (NO)과 prostaglandin E2 (PGE2)와 같은 염증 반응물질을 과량 발생시키 고 이로써 초기 면역 반응과 염증반응이 악화된다(Hwang et al., 1997; Cheng et al., 2017).
염증을 완화시키기 위한 물질로 주로 아스피린과 비스테로이드 항염증제 (non-steroidal anti-inflammatory drugs, NSAIDs/NAIDs)가 사용되고 있지만, NSAID는 장기 복용 시 소장에 염증이 생기고 상부 위장관의 점막에 손상이 생기는 등의 부작용이 보고되었다(Allison et al., 1992). 이로 인해 질환의 예방 및 개선 효과가 있고 안전성이 보장된 합성 의약품을 대 체할 수 있는 천연 의약품의 중요성이 더욱 부각됨에 따라 천연물 유래 기능성 소재를 탐 색하는 연구가 요구되고 있다.
익모초(Leonurus japonicus Houtt.)는 꿀풀과(Labiaceae)에 속하는 2년생 초본 식물로 한국, 중국, 일본 등의 아시아 지역에 널리 분포하고 있다. 익모초(益母草)라는 이름은 여성에게 좋은 약초라는 뜻으로 월경불순, 생리통, 자궁출혈 등 여성질환 치료에 이용되어 왔다 (Shang et al., 2014). 익모초에는 leonurine, leonuridine, rutin과 같은 다양한 활성성분이 함유 되어 있으며, 항산화(Kim et al., 2005), 항염증(Shin et al., 2009), 기억력 및 인지기능 개선 효과(Lee et al., 2020) 등의 활성이 보고되었으나, 익모초 지상부 70% 에탄올 추출물의 염 증 조절 기전 연구는 부족한 실정이다.
따라서 항산화 및 항염증 활성을 가진 천연물 소재 탐색의 일환으로써 본 연구에서는 익 모초 추출물을 이용하여 항산화 활성을 평가하고, MAPKs/NF-ĸB pathway에서의 염증 조절 반응을 통해 항염증 효능을 검증하고자 한다.
Ⅱ. 재료 및 방법
1. 실험재료본 실험에서 사용한 익모초(Leonurus japonicus Houtt.)는 경북 영천에서 재배되어 지상부 만 건조된 것을 구입하여 사용하였다. 건조된 시료는 분쇄기(HR2171, Artreal Huiyang, Man- ufacturing Ltd, China)를 사용하여 분말로 분쇄한 후 실험에 사용하였다.
2. 시료 추출
분쇄한 익모초 분말 10 g에 70% ethanol을 100 mL 가하여 총 20 g을 2시간씩 3회 추출한 뒤 여과(filter paper, kimble-filtering flask)하였다. 추출물은 rotary vacuum evaporator (JP/N- 1000X, ELISA)을 이용해 45℃에서 감압 농축하여 용매를 증발시킨 뒤 동결건조 하였다. 동 결건조 된 분말 시료는 dimethyl sulfoxide (DMSO)에 녹여 50 mg/mL의 stock 용액으로 제조 한 뒤 -20℃에 보관하여 실험에 사용하였다.
3. 폴리페놀 함량 측정
총 폴리페놀 함량은 Folin-Denis 방법을 사용하여 분석하였다(Kiselova et al., 2006). 익모 초 추출 시료 25 µL에 500 µL의 10% folin-ciocalteu’s phenol reagent를 가하고 차광 상태로 5분간 반응시킨 뒤, 500 µL의 7.5% sodium carbonate를 첨가하여 37.5℃에서 1시간 동안 반 응시켰다. Microplate reader (Multiscan spectrum, Thermo Scientific)를 이용하여 725 nm에서 흡광도를 측정하였고 gallic acid (GAE)를 표준물질로 하여 검량선을 그린 뒤, 이로부터 시 료의 총 폴리페놀 함량을 구하였다.
4. 플라보노이드 함량 측정
총 플라보노이드 함량은 Davis법을 변형해서 측정하였다(Byun et al., 2016). 익모초 추출 시료 300 µL에 diethylene glycol 600 µL을 넣고 혼합한 뒤 차광 상태로 5분 반응시킨 후, 1 N
NaOH를 10 µL 더하여 37.5℃에서 1시간 동안 반응시켰다. 이후에 microplate reader (Multiscan spectrum, Thermo Scientific)를 이용하여 420 nm에서 흡광도를 측정하였고, 총 플라보노이드 함량은 Rutin을 표준물질로 한 검량선에 의해 계산하였다.
5. DPPH radical 소거활성
DPPH radical 소거 활성은 Blois (1958)의 방법에 기초하여 시료와 안정적인 자유 라디칼 인 1,1-diphenyl-2-picryl hydrazyl (DPPH)가 반응했을 때 시료의 수소공여효과 및 라디칼 소 거 능력으로 측정하였다(Marinova and Batchvarov, 2011). 100 µL의 추출물에 methanol로 희 석시킨 0.2 mM의 DPPH 용액 100 µL를 혼합한 뒤 차광 후 37.5℃에서 30분 동안 반응시켰 다. 이후 microplate reader (Multiscan spectrum, Thermo Scientific)를 이용하여 517 nm에서 흡 광도를 측정하고, ascorbic acid를 양성대조구로 사용한 DPPH radical 소거활성은 다음과 같 은 식을 이용해 계산하였다.
DPPH radical 소거활성(%)
= [(무첨가구의 흡광도 - 시료첨가구의 흡광도) / 무첨가구의 흡광도]×100 6. ABTS radical 소거활성
ABTS radical 소거 활성은 7 mM ABTS 용액을 2.45 mM의 potassium persulfate와 2:1로 섞 은 후 혼합물을 12~16 h 동안 차광하여 ABTS 라디칼 양이온(ABTS+)을 생성시키고 100%
ethanol로 희석하여 734 nm에서 흡광도 값이 0.70 ± 0.02가 되도록 하였다(Sudha et al., 2011).
시료 100 µL와 ABTS 용액 900 µL를 혼합한 뒤 차광상태로 실온에서 3분간 반응 후 micro- plate reader를 이용하여 734 nm에서 흡광도를 측정하였다. Ascorbic acid를 양성대조구로 사 용하였고 ABTS radical 소거활성은 다음과 같은 식을 이용해 계산하였다.
ABTS radical 소거활성(%)
= [(무첨가구의 흡광도 - 시료첨가구의 흡광도) / 무첨가구의 흡광도]×100 7. 환원력 측정
환원력은 Senevirathne 등(2006)의 방법을 변형하여 측정하였다. 100 µL 시료에 0.2 M sodium phosphate (pH 6.6) 100 µL를 더하여 상온에서 5분간 반응시킨 다음 1% potassium ferricyanide 100 µL를 넣고 50℃의 항온기에서 20분간 열을 가하였다. 혼합 용액에 100 µL
의 10% trichloroacetic acid를 첨가하고 12,000 rpm에서 10분간 원심분리하였다. 96 well plate 에 100 µL의 증류수와 원심분리 한 혼합 용액의 상층액을 100 µL씩 첨가 후 0.1% ferric chloride를 10 µL씩 혼합하고 700 nm에서 흡광도를 측정하였다. 시료의 환원력은 반응 혼합 물의 흡광도에 정비례하는데, 이를 1 mM Rutin과 비교하여 백분율(%)로 나타내었다.
8. 세포배양
실험에 사용된 마우스 대식세포 RAW 264.7은 American Type Culture Collection (ATCC, Rockville, MD, USA)에서 구매해서 사용하였다. 세포의 배양에는 10% FBS (fatal bovine serum)와 1% P/S (penicillin, streptomycin)이 첨가된 DMEM (dulbecco’s modified Eagle’s medium) 배지가 사용되었으며, 세포는 37℃, 5% CO2 조건의 incubator에서 배양되었다.
9. 세포 생존율 측정
세포의 생존율은 cytotoxicity assay kit를 이용해 측정하였다. 96 well plate에 RAW 264.7을 5×105 cells/mL의 농도로 100 µL씩 분주하고 37℃, 5% CO2의 incubator에서 24시간 배양 후 익모초 추출물을 농도별로 (50, 100, 200, 400 µg/mL) 처리하였다. 1시간 뒤 1 µg/mL의 LPS 를 처리 하고 incubator에서 18시간 반응시킨다. 2,500 rpm에서 5분간 원심분리하고 상층액 을 100 µL씩 분리 후 남은 상층액 100 µL에 EZ-Cytox 용액을 10 µL씩 넣고 incubator에서 30분간 반응시킨 후 microplate reader를 사용해 540 nm에서 흡광도를 측정하였다.
10. 세포 내 ROS 측정
세포 내 ROS 수준은 ROS와 반응해 DCF로 탈아세틸화 되며 형광을 나타내는 물질인 DCFH-DA를 사용해 측정하였다. 96 well black plate에 RAW 264.7을 3×105 cells/mL의 농도 로 100 µL씩 분주하고 24시간동안 incubator에서 배양하였다. 익모초 추출물을 50 µL씩 농 도별로 (100, 200, 400 µg/mL) 분주하고 1시간 뒤 LPS (1 µg/mL)를 처리하여 6시간 후 1 mM의 DCFH-DA를 100 µL씩 넣어 Multimode Microplate Reader System (Perkin Elmer, EnSpire)를 이용해 fluorescence intensity를 측정하였다.
11. NO 생성량 측정
LPS에 의해 유도되는 RAW 264.7에서의 NO 발생량은 Griess 반응에 기초하여 측정하였 다. 시료와 LPS 처리 후 incubator에서 18시간 반응시킨 다음 2,500 rpm에서 5분간 원심분리
하고 상층액 100 µL 중 50 µL를 1% sulfanilamide와 0.1% N-1-naphthylethyenediamine (NED) 50 µL와 함께 반응시킨 뒤 microplate reader를 이용하여 540 nm에서 흡광도를 측정하였다.
NO 생성량은 sodium nitrate를 표준물질로 하여 표준곡선을 그려 계산하였다.
12. RAW264.7 cell의 형태학적 변화
LPS와 익모초 추출물 처리에 의한 세포의 형태학적 변화를 관찰하기 위해 다음과 같은 실험을 수행하였다. 96 well plate에 RAW 264.7 cell을 3×105 cells/mL로 37℃, 5%의 incubator 에서 24시간 배양한 후, 익모초 추출물을 농도별로 (50, 100, 200 µg/mL) 처리하여 LPS (1 µg/
mL)와 함께 18시간 반응시켰다. Nikon eclipse TS100 위상차현미경 (Nikon Instruments Inc.
Melville, NY, USA) 및 HK Basic 3.7 program (Koptic)을 이용해 세포 image를 촬영하였다.
13. Cytokine 생성량 측정
LPS 자극에 의해 생성되는 cytokine인 TNF-α및 IL-1β의 생성량은 ELISA kit (R&D systems, Mineeapolis, MN, USA, Cat No. SMLB00C, SMTA00B)를 사용하여 제조사의 실험 방법에 따라 측정하였다. 96 well plate에 RAW 264.7을 5×105 cells/mL의 농도로 100 µL씩 분주하고 24시간 배양한 다음, 익모초 추출물을 농도별로 1시간 전처리하여 LPS (1 µg/mL) 와 함께 incubator에서 18시간 반응시켰다. 2,500 rpm에서 5분간 원심분리 후 상층액의 TNF-α 및 IL-1β의 발현 수준을 측정하였다.
14. Western blot analysis
익모초 추출물의 염증성 단백질 발현에 미치는 영향을 확인하기 위해 western blot analysis를 실시했다. 6 well plate에 RAW 264.7 cell을 5×105 cells/mL 및 1×106 cells/mL 농도 로 분주하고 익모초 추출물을 농도별로 1시간 전처리 후 LPS 1 µg/mL와 함께 18시간 또는 1시간 배양하였다. 세포 단백질은 RIPA lysis buffer를 이용하여 추출하였으며, bio-rad protein assay reagent (Santa Cruz, CA, USA)를 이용해 단백질 정량 후 분석에 이용하였다. SDS- PAGE를 이용해 단백질을 크기별로 분리 후 100V, 1시간 10분 동안 transfer하였다. 이후 5% skim milk를 이용해 1시간 30분 동안 blocking 한 뒤 1X PBST를 이용해 세척 후 1차 항 체(1:1000)를 반응시켰다. 2차 항체는 1:10000의 희석비율로 2시간 동안 실온에서 배양한 후 western blotting luminol reagent A&B (sc-2048, Santa Cruz, CA, USA)를 1:1로 혼합하여 발색 시켰다. 단백질 발현량은 chemidoc 및 image software (Bio-Rad Laboratories, Inc, USA)을 이 용해 확인하였다.
15. 통계처리
실험 결과는 3반복으로 하여 평균 ± 표준편차로 나타내고 모든 통계 처리는 SPSS v12.0 (SPSS Inc., Chicago, IL, USA)를 이용하여 one-way ANOVA 분석 및 LSD 사후분석을 실시 해 p<0.05일 경우 통계학적으로 유의성이 있다고 판단하였다(*p<0.05; **p<0.01; ***p<0.001).
Ⅲ. 결과 및 고찰
1. 총 폴리페놀 및 플라보노이드 함량페놀화합물은 식물계에 가장 많고 널리 분포하는 2차 대사산물로서 플라보노이드, 안토 시아닌, 탄닌, 카테킨 등 방향환을 가진 화합물의 총칭이며, 이들은 이상적인 구조 화학을 가지고 있어 자유 라디칼을 소거하는 등의 항산화제로서 역할을 한다고 알려져 있다(Urquiaga and Leighton, 2000). 본 실험에서 익모초 추출물의 총 폴리페놀 함량은 1 mg/mL의 농도에 서 51.40 ± 0.47 mg GAE/g으로 나타났다(Table 1). Ravipati 등(2012)의 연구에서 보고된 익모 초 증류수 추출물 6.46 ± 0.82 mg GAE/g, 95% 에탄올 추출물 0.84 ± 2.66 mg GAE/g로 나타 난 측정 결과와 비교하였을 때 본 연구에서의 70% 에탄올 추출물에서 더 높은 함량이 측 정되었음을 확인할 수 있었다.
페놀화합물에 속하는 플라보노이드는 야채와 과일뿐만 아니라 곡물, 견과류, 향신료 및 약용식물에서도 잘 발견되며 과산화물 음이온 생성을 담당하는 효소를 억제시키고, 산화되 는 자유 라디칼을 줄이는 역할을 하여 폴리페놀과 마찬가지로 항산화 효과가 있는 것으로 알려졌다(Pietta, 2000). 본 실험에서 확인된 1 mg/mL 익모초 추출물의 총 플라보노이드 함 량은 73.28 ± 0.10 mg rutin/g으로 확인되었다(Table 1).
Table 1. Total polyphenol and flavonoid contents of L. japonicus extract
L. japonicus Polyphenol
(mg GAE/g)
Flavonoid (mg rutin/g)
Contents 51.40 ± 0.47ª 73.28 ± 0.10
ª mean ± standard deviation (SD).
2. DPPH 및 ABTS radical 소거활성
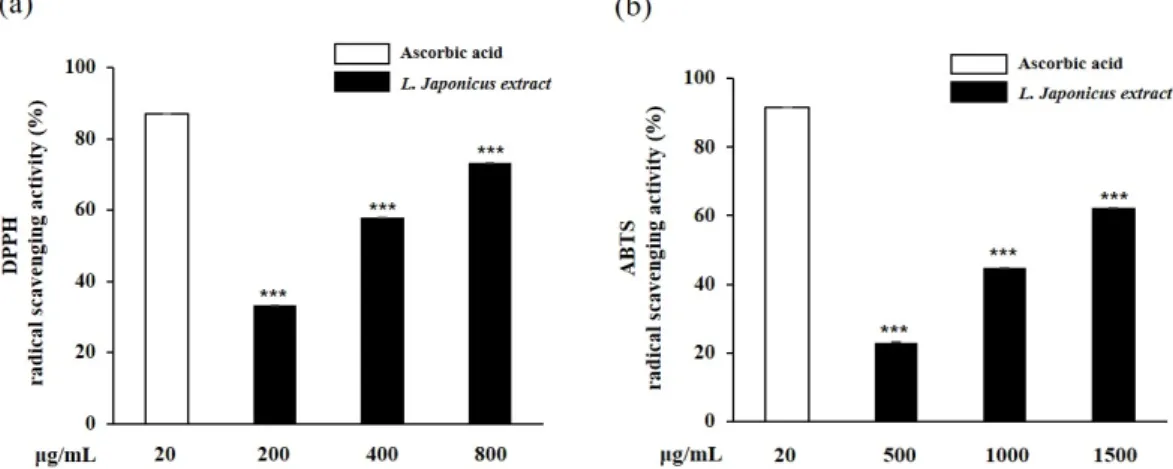
익모초 추출물의 농도별 DPPH radical 소거능을 평가한 결과, 익모초 200, 400, 800 µg/
mL의 농도에서 각각 33.2%, 57.8%, 73.3%의 농도 의존적인 소거활성을 나타냈다(Fig. 1a).
Lee 등(2017)의 연구에서는 익모초 줄기 추출물의 IC50값이 52.77 µg/mL로 본 연구결과에 비해 다소 소거능이 높게 나타났는데, 이는 추출 부위 및 추출 용매의 차이일 것으로 판단 된다.
ABTS 소거능 측정방법은 DPPH 소거능과 함께 항산화 활성 평가에 자주 이용되지만 자 유라디칼이 제거되는 DPPH assay와는 달리 양이온 라디칼이 제거되는 차이가 있다(Yu et al., 2012). 익모초 추출물의 ABTS radical 소거능을 평가한 결과 500, 1000, 1500 µg/mL의 농도에서 각각 23.0%, 44.7%, 62.3%의 소거활성을 보였는데(Fig. 1b), 익모초 95% 에탄올 추출물의 ABTS radical 소거활성의 IC50이 7894.3 µg/mL인 Oliveira 등(2017)의 연구 결과와 비교하였을 때 더 낮은 농도에서 높은 효과를 보였으며, 이는 추출 용매의 극성에 따른 페 놀성 물질의 용출 차이로 인해 나타나는 항산화력의 차이일 것으로 사료된다.
Fig. 1. DPPH (a) and ABTS (b) radical scavenging activities of L. japonicus extract. Each value represented mean±SD (n=3). *** p<0.001 compared with untreated control.
3. 환원력
환원력은 산화물을 환원시키는 힘으로 산화물에 전자를 공여하는 능력이 클수록 환원력 이 크다. 활성산소가 전자를 얻게 되면 금속이온 ferricyanide (Fe3+)를 ferrous (Fe2+)로 환원 시켜 노란색의 혼합용액이 녹색으로 변하게 되는데, 이와 같은 반응을 적용하여 시료의 전자 공여 능력에 따른 환원력 정도를 측정하여 항산화 능력을 알 수 있다(Ferreira et al., 2007).
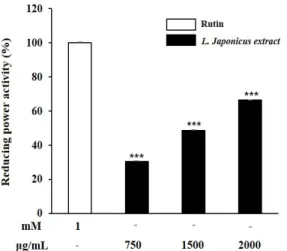
표준물질로 사용된 1 mM Rutin과 비교했을 때 익모초 70% 에탄올 추출물은 750, 1500, 2000 µg/mL의 농도에서 30.6%, 48.7%, 57.9%의 환원력을 보였다(Fig. 2).
Fig. 2. Reducing power activity of L. japonicus extract. Each value represented mean±SD (n=3). ***p<0.001 compared with 1 mM Rutin standard control.
4. 세포 내 ROS 소거능
ROS는 산소분자가 호흡과정을 통해 불완전하게 환원되며 생기는 일종의 부산물로 염증 반응 시 NO와 함께 ROS가 생성된다(Das and Roychoudhury, 2014). ROS가 과도하게 생성되 면 DNA, 단백질 및 지질에 손상을 입히고 이로 인해 종양 유발 신호의 활성화 등으로 인 체에 유해한 반응이 일어나게 된다(Moloney and Cotter, 2018). LPS를 처리해 ROS 생성을
Fig. 3. Effect of L. japonicus extract on ROS production in LPS-induced RAW 264.7 cells.
Each value represented mean ± SD (n=6). ###p<0.001 compared with normal con- trol; ***p<0.001 compared with LPS control.
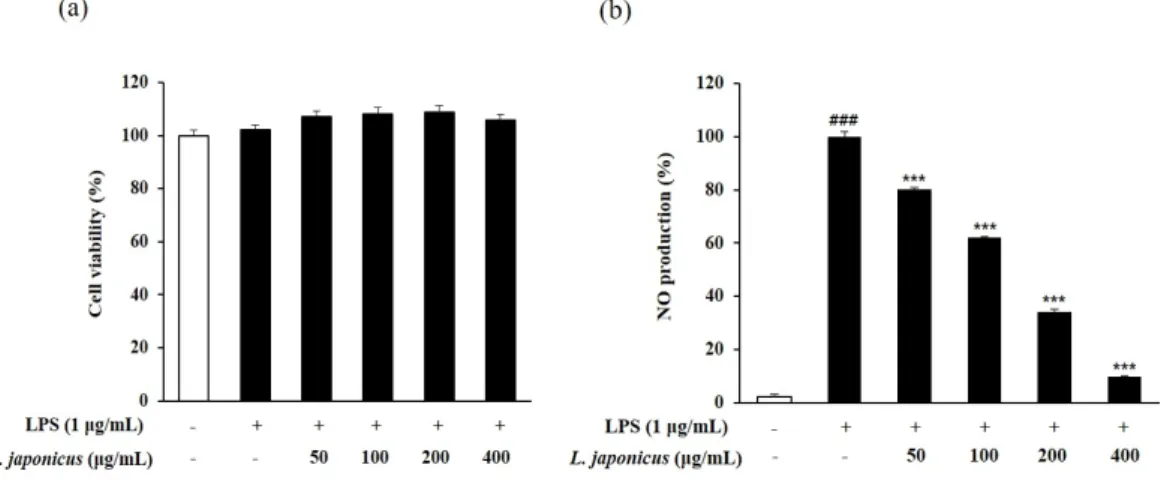
유도한 RAW 264.7에서 익모초 추출물의 ROS 소거능을 확인한 결과 LPS의 자극으로 인해 ROS의 생성이 유의미하게 증가했지만, 익모초 추출물을 농도별로 (100, 200, 400 µg/mL) 처리하였을 때 ROS의 생성이 각각 93.0%, 78.7%, 69.5%로 점차 감소하였다(Fig. 3).
5. 세포 생존율
RAW 264.7 cell에서 익모초 추출물의 독성 여부를 판단하기 위해 cell viability를 측정한 결과 익모초 추출물 50~400 µg/mL의 농도에서 세포 생존율이 모두 100% 이상으로 나타나 (Fig. 4a), 익모초는 RAW 264.7 cell에 대한 독성이 없음을 확인하였다.
Fig. 4. Effect of L. japonicus extract on cell viability (a) and NO production (b) on LPS induced RAW 264.7 cells. Each value represented mean±SD (n=3). ###p<0.001 compared with normal control; ***p<0.001 compared with LPS control.
6. NO 생성 억제 효과
NO는 세포 신호전달에 중요한 분자로 작용하지만, LPS 또는 염증성 사이토카인 등의 자 극에 의해 NO가 과도하게 생성되면서 염증반응이 촉진되고 염증성 장 질환, 관절염, 동맥 경화증, 종양과 같은 다양한 만성 질환이 유발된다(Zhai et al., 2009; Kenny et al., 2013). 이 에 본 실험에서는 LPS를 처리한 마우스 대식세포에서 익모초 추출물이 NO 생성에 미치는 영향을 확인하였다. 무처리군에 비해 NO 생성량이 크게 증가한 LPS 처리군과 비교하였을 때, 익모초 추출물 처리군에서 NO 발생량이 농도 의존적으로 유의하게 감소하였고, 400 µg/mL에서는 90.3%의 NO 생성 억제 활성이 확인되었다(Fig. 4b).
7. RAW 264.7 cell의 형태학적 변화
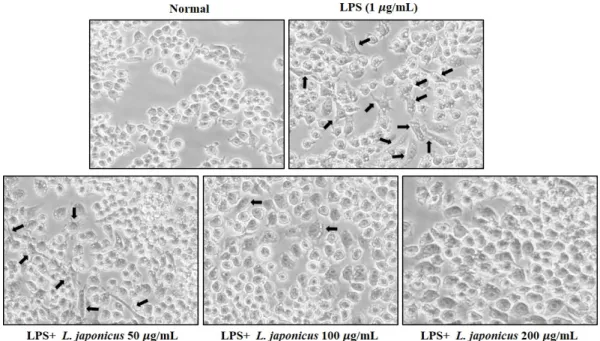
RAW 264.7에 LPS와 같은 자극이 가해지면, 염증성 전사인자 및 단백질이 발현될 뿐만 아 니라 세포의 형태학적 변화가 관찰된다. LPS를 처리한 RAW 264.7 cell에서 익모초 추출물이 세포의 형태학적 변화에 미치는 영향을 확인하였다. 정상 세포와 비교하였을 때 LPS를 처리 한 세포에서 유의한 형태학적 변화가 관찰되었으며, 익모초 추출물을 50, 100, 200 µg/mL로 처리한 세포에서는 LPS에 의해 변형된 세포의 수가 점차 감소하는 것을 확인하였다(Fig. 5).
Fig. 5. Morphological transformation of RAW 264.7 cell treated with LPS (1 µg/mL) and L.
japonicus extract (50, 100, 200 µg/mL).
8. Cytokine 생성량 측정
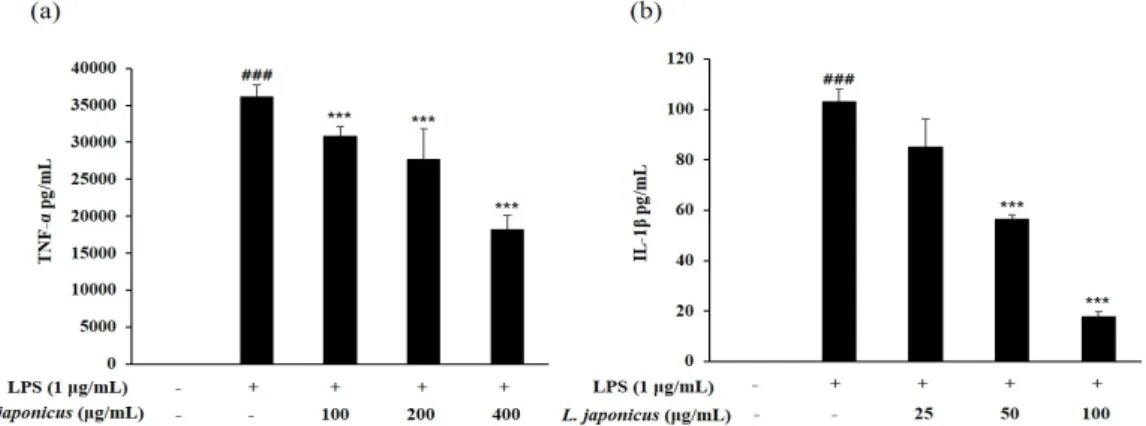
Cytokine은 우리 몸 면역 체계의 신호전달 물질로서 면역과 염증반응에 관여하며 대식세 포가 바이러스와 같은 병원체에 의해 자극을 받게 되면 대표적인 염증성 사이토카인인 TNF-α와 IL-1β이 발현이 된다. TNF-α는 내피세포 및 백혈구를 자극해 NO 생성 및 IL1, IL6 등의 방출을 유도하고(Hu et al., 2008), IL-1β는 국소 및 전신 수준에서 염증반응을 매 개하는데(Hu et al., 2014), 이러한 염증 매개 물질의 과도한 생성은 염증반응을 지속시켜 만 성 염증성 질환을 유발하는 것으로 알려져 있다. 따라서 본 연구에서는 LPS로 자극한 대식 세포에 익모초 추출물을 처리하였을 때 TNF-α 및 IL-1β의 생성에 미치는 영향을 확인하
고자 하였다. TNF-α 함량이 35000 pg/mL까지 증가한 LPS 처리군과 비교하여 익모초 추출 물을 처리한 경우 100, 200, 400 µg/mL 농도 모두에서 TNF-α생성량이 유의적으로 감소하 였다(Fig. 6a). IL-1β 또한 무처리군에 비해 LPS 처리구에서 함량이 크게 증가했으며 익모 초 추출물 농도가 증가함에 따라 생성량이 감소해 100 µg/mL의 농도에서는 82.7% 억제됨 을 확인하였다(Fig. 6b). 이러한 결과는 익모초 70% 에탄올 추출물은 염증성 사이토카인인 TNF-α와 IL-1β의 생성을 억제함으로써 항염증 효과를 보이는 것으로 사료된다.
Fig. 6. Effects of L. japonicus extract on LPS-induced cytokine production in RAW 264.7 cells. Each value represented mean±SD (n=3). ###p<0.001 compared with normal control; ***p<0.001 compared with LPS control.
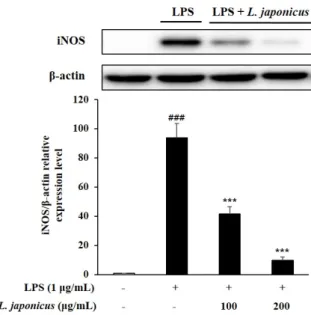
9. iNOS 발현 억제 효과
iNOS는 염증반응에 중요한 역할을 하는 단백질로 면역세포 활성화에 의해 발현되고 이 로 인해 다량의 NO가 생성된다. 본 연구에서 익모초 추출물에 의한 iNOS의 발현양상을 확 인한 결과, LPS 처리시 iNOS의 발현량이 무처리구에 비해 확연히 증가하였으나 익모초 추 출물 처리 시 발현량이 농도 의존적으로 감소하였고, 특히 200 µg/mL에서는 무처리구와 거 의 유사한 수준의 발현량이 확인되었다(Fig. 7). 익모초 추출물 처리에 의한 iNOS 발현 억 제 양상은 NO 생성 억제 효과와 유사한 경향을 나타냈으며, 이는 익모초 추출물이 NO 생 성 유도체인 iNOS의 발현을 억제함에 따라 NO의 생성이 감소한 것으로 사료된다.
Fig. 7. Effects of L. japonicus extract on LPS-induced iNOS protein expression in RAW 264.7 cells. Each value represented mean±SD (n=3). ###p<0.001 compared with normal control; ***p<0.001 compared with LPS control.
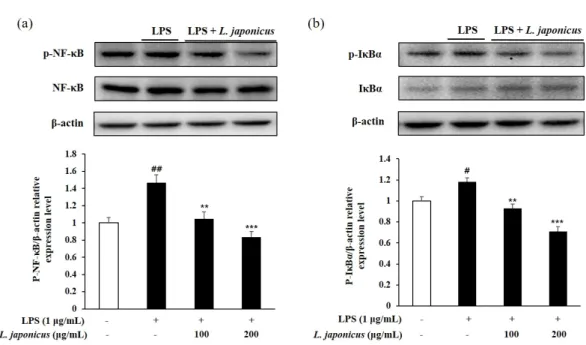
10. NF-ĸB 및 IĸB-α 인산화 억제 효과
NF-ĸB는 DNA의 전사, 염증반응, 세포 증식에 관여하는 진핵 전사인자 중 하나로 정상 조건에서는 억제 단백질인 IĸB-α에 결합하여 세포질에 존재하지만, LPS 및 ROS와 같은 외부 자극이 가해지면 IĸB-α의 인산화가 촉진되고 NF-ĸB가 이로부터 해리 및 핵으로 전 위되어 cytokine, iNOS, COX-2와 같은 염증 매개체들의 전사를 촉진한다(Surh et al., 2001).
이에 본 연구에서는 LPS를 처리한 RAW 264.7에서 익모초 추출물을 처리했을 때 NF-ĸB 및 IĸB-α 인산화 억제 정도를 확인하였다. NF-ĸB와 IĸB-α 모두 무처리구에 비해 LPS 처리구 에서 인산화 수준이 증가하였고 100, 200 µg/mL 농도의 익모초 추출물을 처리했을 때 인산 화 수준이 유의적으로 감소하였다(Fig. 8). 익모초 추출물은 억제 단백질인 IĸB-α의 인산화 를 조절함으로써 NF-ĸB의 인산화 및 해리를 억제했을 뿐만 아니라 iNOS 및 NO의 생성 또 한 유의적으로 감소시켰는데 이를 종합해볼 때 iNOS의 발현은 NF-ĸB에 의해 조절된다는 것을 확인할 수 있다.
Fig. 8. Inhibit phosphorylation level of NF-ĸB (a) and IĸB-α (b) in LPS-induced RAW 264.7 cell. Each value represented mean±SD (n=3). ##p<0.01 and #p<0.05 compared with normal control; ***p<0.001 and **p<0.01 compared with LPS control.
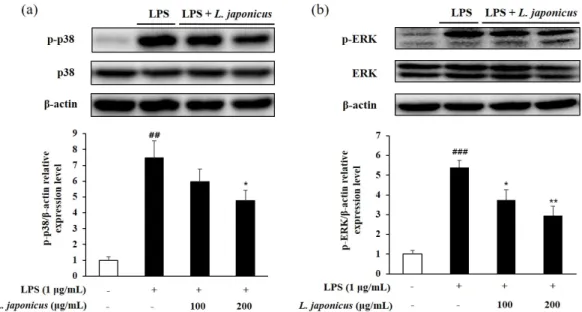
11. ERK, JNK, p38 인산화 억제 효과
MAPK는 외부 스트레스 신호에 대한 염증반응, 세포 분화 및 증식, 세포 사멸 등과 같은 많은 세포 반응을 조절하는 데 중요한 역할을 하며 ERK 1/2, JNK, p38 MAP kinase가 MAPK의 주요한 subgroup으로 알려져 있다(Kaminska, 2005). LPS와 같은 외부 자극으로 MAPK cascade pathway가 유도되면 ERK, JNK, p38 등의 MAPK 인자들이 인산화 되며 이는 NF-ĸB의 활성화까지 이어진다(Lee et al., 2012). LPS로 활성화된 RAW 264.7에서 익모초 추 출물을 처리했을 때 ERK, p38, JNK의 인산화 억제 수준을 확인한 결과 LPS로 활성화된 cell에서는 ERK 및 p38의 인산화 수준이 증가하였으나 익모초 추출물을 처리한 cell에서는 그 수준이 유의적으로 감소하였다(Fig. 9). 그러나 JNK 인산화에는 영향을 미치지 않는 것 으로 확인되었다. p38 MAPK는 cytokine의 생산과 TNF-α가 매개하는 downstream signaling pathway에도 관여해 염증반응에 중요한 역할을 하는 것으로 보고되었는데(English and Cobb, 2002; Muthusamy and Piva, 2010), 익모초 추출물이 p38의 인산화 및 TNF-α 및 IL1-β의 생 산을 억제한 것을 보아 p38의 인산화 수준이 염증성 사이토카인과 관련이 있음을 확인할 수 있다.
Fig. 9. Inhibit phosphorylation level of p38 (a) and ERK (b) in LPS-induced RAW 264.7 cell. Each value represented mean±SD (n=3). ###p<0.001 and ##p<0.01 compared with normal control; **p<0.01 and *p<0.05 compared with LPS control.
Ⅴ. 적 요
본 실험에서는 익모초 지상부 에탄올 추출물의 항산화 및 항염증 효과를 확인하여 건강 기능성 소재로서의 가능성을 평가하고자 하였다. DPPH 및 ABTS radical 소거 활성, 환원력, 총 폴리페놀 및 플라보노이드 함량을 통해 익모초의 항산화능을 측정한 결과, 400 µg/mL, 1500 µg/mL의 농도에서 57.8%, 62.3%의 DPPH 및 ABTS radical 소거활성을 보였고 환원력 또한 농도 의존적으로 증가하는 경향을 보였다. 총 폴리페놀 함량 및 플라보노이드 함량은 1 mg/mL의 농도에서 각각 51.40 ± 0.47 mg of gallic acid equivalents/g, 73.28 ± 0.10 mg of rutin equivalents/g로 나타났으며, 익모초 추출물은 세포 내 ROS의 생성 억제에 있어서 유의 적인 효과를 보였다. 익모초의 항염증 효과를 측정한 결과, LPS로 자극해 활성화된 RAW 264.7 cell에서 익모초 추출물(0~400 µg/mL)의 세포 독성은 없었으며 LPS 처리로 유도된 세 포의 형태학적 변화도 농도의존적으로 완화되는 경향을 보였다. NO 발생량은 LPS 처리군 과 비교해 익모초 추출물 처리 시 농도 의존적으로 감소하였고, 400 µg/mL에서는 90.3%로 NO의 생성이 억제되었다. 염증성 cytokine (TNF-α, IL1-β)의 생성도 유의적으로 감소하였 고 NO를 생성하는 염증성 단백질 iNOS의 발현 또한 억제되었으며 이와 같은 염증성 단백 질의 전사를 조절하는 NF-ĸB (NF-ĸB, IĸB-α) 및 MAPK (ERK, p38)의 인산화 및 활성화 또
한 익모초 추출물 처리로 인해 억제됨을 확인하였다. 따라서 익모초 추출물은 NF-ĸB signaling pathway 및 ERK/p38 MAPK cascade pathway의 조절을 통해 염증성 단백질 및 염증인자의 발현을 감소시킨다고 볼 수 있다. 이러한 결과를 토대로 익모초 지상부 에탄올 추출물은 천연 기능성 소재로서 활용될 가능성이 있으며, 본 연구 결과는 익모초의 고부가가치 향상 을 위한 기초자료로 도움이 될 것으로 생각된다.
[Submitted, October. 26, 2020; Revised, November. 3, 2020; Accepted, November. 4, 2020]
References
1. Allison, M. C., A. G. Howatson, C. J. Torrance, F. D. Lee, and R. I. Russell. 1992.
Gastrointestinal Damage Associated with the Use of Nonsteroidal Antiinflammatory Drugs.
N. Engl. J. Med. 327: 1882-1883.
2. Anand, S. P. and N. Sati. 2013. Artificial Preservatives and Their Harmful Effects: Looking Toward Nature for Safer Alternatives. Int. J. Pharm. Sci. Res. 4: 2496-2501.
3. Birben, E., U. M. Sahiner, C. Sackesen, S. Erzurum, and O. Kalayci. 2012. Oxidative stress and antioxidant defense. World Allergy Organ. J. 5: 9-19.
4. Blois, M. S. 1958. Antioxidant determinations by the use of a stable free radical. Nature.
181: 1199-1200.
5. Byun, E. B., W. Y. Park, D. H. Ahn, Y. C. Yoo, C. Park, B. S. Jang, W. J. Park, E. H.
Byun, and N. Y. Sung. 2016. Comparison study of three varieties of red peppers in terms of total polyphenol, total flavonoid contents, and antioxidant activities. Korean J. Food Sci.
Nutr. 45: 765-770.
6. Chapple, I. L. 1997. Reactive oxygen species and antioxidants in inflammatory diseases. J.
Clinl. Periodontol. 24: 278-296.
7. Cheng, P., T. Wang, W. Li, I. Muhammad, H. Wang, X. Sun, Y. Yang, J. Li, T. Xiao, and X. Zhang. 2017. Baicalin alleviates lipopolysaccharide-induced liver inflammation in chicken by suppressing TLR4-mediated NF-κB pathway. Front Pharmacol. 8: 547.
8. Das, K. and A. Roychoudhury. 2014. Reactive oxygen species (ROS) and response of anti- oxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2: 53.
9. Dinarello, C. A. 2010. Anti-inflammatory Agents: Present and Future. Cell. 140: 935-950.
10. English, J. M. and M. H. Cobb. 2002. Pharmacological inhibitors of MAPK pathways. In
Trends Pharmacol. Sci. 23: 40-45.
11. Ferreira, I. C. F. R., P. Baptista, M. Boas, and L. Barros. 2007. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal: Individual cap and stipe activity. Food Chem. 100: 1511-1516.
12. Hu, B., H. Zhang, X. Meng, F. Wang, and P. Wang. 2014. Aloe-emodin from rhubarb (Rheum rhabarbarum) inhibits lipopolysaccharide-induced inflammatory responses in RAW264.7 macrophages. J. Ethnopharmacol. 153: 846-853.
13. Hu, X. D., Y. Yang, X. G. Zhong, X. H. Zhang, Y. N. Zhang, Z. P. Zheng, Y. Zhou, W.
Tang, Y. F. Yang, L. H. Hu, and J. P. Zuo. 2008. Anti-inflammatory effects of Z23 on LPS-induced inflammatory responses in RAW264.7 macrophages. J. Ethnopharmacol. 120:
447-151.
14. Hwang, D., B. C. Jang, G. Yu, and M. Boudreau. 1997. Expression of mitogen-inducible cyclooxygenase induced by lipopolysaccharide. Biochem. Pharmacol. 54: 87-96.
15. Kaminska, B. 2005. MAPK signalling pathways as molecular targets for anti-inflammatory therapy - From molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta. 1754:
253-262.
16. Kenny, O. M., C. M. McCarthy, N. P. Brunton, M. B. Hossain, D. K. Rai, S. G. Collins, P. W. Jones, A. R. Maguire, and N. M. O’Brien. 2013. Anti-inflammatory properties of potato glycoalkaloids in stimulated Jurkat and Raw 264.7 mouse macrophages. Life Sci. 92:
775-782.
17. Kim, J. Y., Y. H. Lee, J. Y. Kim, and R. B. Kyung. 2005. Study of Antioxidation Action of Lenonuri herba Extract. Korean J. Cosmetric Sci, 51: 189-196.
18. Kiselova, Y., D. Ivanova, T. Chervenkov, D. Gerova, B. Galunska, and T. Yankova. 2006.
Correlation between the in vitro antioxidant activity and polyphenol content of aqueous extracts from Bulgarian herbs. Phytother. Res. 20: 961-965.
19. Lee, H. S., D. S. Ryu, G. S. Lee, and D. S. Lee. 2012. Anti-inflammatory effects of dichloromethane fraction from Orostachys japonicus in RAW 264.7 cells: Suppression of NF-κB activation and MAPK signaling. J. Ethnopharmacol. 140: 271-276.
20. Lee, J., H.-J. Kim, G. Y. Jang, K. H. Seo, M. R. Kim, Y. H. Choi, and J. W. Jung. 2020.
Effect of Leonurus japonicus Houtt. on Scopolamine-induced Memory Impairment in Mice.
J. Physiol. Pathol. Korean Med. 34: 81-87.
21. Lee, S., D. Lee, J. Baek, E. B. Jung, J. Y. Baek, I. K. Lee, T. S. Jang, K. S. Kang, and K. H. Kim. 2017. In vitro assessment of selected Korean plants for antioxidant and anti- acetylcholinesterase activities. Pharm. Biol. 55: 2205-2210.
22. Lee, S. G., D. J. Jo, H. J. Chang, and H. Kang. 2015. Antioxidant and Anti-inflammatory Activities of Ethanol Extracts from Aralia continentalis Kitagawa, Korean J. Naturopathy. 4:
10-14.
23. Marinova, G. and V. Batchvarov. 2011. Evaluation of the methods for determination of the free radical scavenging activity by DPPH. Bulgarian J. Agri. Sci. 17: 11-24.
24. Moloney, J. N. and T. G. Cotter. 2018. ROS signalling in the biology of cancer. Semin Cell Dev. Biol. 80: 50-64.
25. Muthusamy, V. and T. J. Piva. 2010. The UV response of the skin: A review of the MAPK, NFκB and TNFα signal transduction pathways. Arch Dermatol. Res. 302: 5-17.
26. Oliveira, A. S., L. M. Cercato, M. T. de Santana Souza, A. J. O. Melo, B. D. S. Lima, M.
C. Duarte, A. A. S. Araujo, A. M. de Oliveira E Silva, and E. A. Camargo. 2017. The ethanol extract of Leonurus sibiricus L. induces antioxidant, antinociceptive and topical anti-inflammatory effects. J. Ethnopharmacol. 206: 144-151.
27. Pietta, P. G. 2000. Flavonoids as antioxidants. J. Nat. Prod. 63: 1035-1042.
28. Ravipati, A. S., L. Zhang, S. R. Koyyalamudi, S. C. Jeong, N. Reddy, J. Bartlett, P. T.
Smith, K. Shanmugam, G. Münch, M. J. Wu, M. Satyanarayanan, and B. Vysetti. 2012.
Antioxidant and anti-inflammatory activities of selected Chinese medicinal plants and their relation with antioxidant content. BMC Complement Altern. Med. 12: 173.
29. Senevirathne, M., S. H. Kim, N. Siriwardhana, J. H. Ha, K. W. Lee, and Y. J. Jeon. 2006.
Antioxidant potential of ecklonia cavaon reactive oxygen species scavenging, metal chelat- ing, reducing power and lipid peroxidation inhibition. Food Sci. Technol. Int. 12: 27-38.
30. Shang, X., H. Pan, X. Wang, H. He, and M. Li. 2014. Leonurus japonicus Houtt.:
Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 152: 14-32.
31. Shin, H. Y., S. H. Kim, S. M. Kang, I. J. Chang, S. Y. Kim, H. Jeon, K. H. Leem, W. H.
Park, J. P. Lim, and T. Y. Shin. 2009. Anti-inflammatory activity of Motherwort (Leonurus sibiricus L.). Immunopharmacol. Immunotoxicol. 31: 209-213.
32. Sudha, G., M. S. Priya, R. I. Shree, and S. Vadivukkarasi. 2011. In vitro free radical scavenging activity of raw pepio fruit (Solanum muricatum aiton). Int. J. Curr. Pharml. Res. 3.
33. Surh, Y. J., K. S. Chun, H. H. Cha, S. S. Han, Y. S. Keum, K. K. Park, and S. S. Lee.
2001. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-κB activation. Mutat. Res. 480-481: 243-268.
34. Urquiaga, I. and F. Leighton. 2000. Plant Polyphenol Antioxidants and Oxidative Stress.
Biol. Res. 33: 55-64.
35. Yu, S. Y., Y. J. Lee, H. S. Song, H. J. Lim, H. S. Choi, B. Y. Lee, S. N. Kang, and O.
H. Lee. 2012. Antioxidant Effects and Nitrite Scavenging Ability of Extract from Acanth- opanax cortex Shoot. Korean J. Food Nutr. 25: 793-799.
36. Zhai, Z., A. Solco, L. Wu, E. S. Wurtele, M. L. Kohut, P. A. Murphy, and J. E. Cunnick.
2009. Echinacea increases arginase activity and has anti-inflammatory properties in RAW 264.7 macrophage cells, indicative of alternative macrophage activation. J. Ethnopharmacol.
122: 76-85.