대한소아혈액종양학회지
제 11 권 제 1 호 2004
7
책임저자:안효섭, 서울시 종로구 연건동 28번지, 서울대학교 병원 소아과, 110-744 Tel: 02-760-3625, Fax: 02-743-3455, E-mail: [email protected]
위 연구는 21세기 프론티어연구개발사업인 세포응용연구사업단의 연구비지원(과제관리번호 SC13062)에 의하여 수행되 었음.
증식과 분화
서울대학교 의과대학 소아과학교실
최형수․고상혁․박은실․신희영․안효섭
The Effect of Stromal Cells and Hematopoietic Growth Factors on Proliferation and Differentiation of Megakaryocytic
Progenitors from Umbilical Cord Blood
Hyoung Soo Choi, M.D., Sang Hyeok Koh, M.A., Eun Sil Park, M.D., Hee Young Shin, M.D. and Hyo Seop Ahn, M.D.
Department of Pediatrics, Seoul National University College of Medicine, Seoul, Korea
Purpose: CD34
+cells from umbilical cord blood (UCB) were cultured with stromal cells and growth factors, and cell proliferation and megakaryocytic differentiation were observed. The purposes of this study are to find out the optimal culture condition for the megakaryocytic differentiation of CD34
+cells from UCB. Methods: CD34
+cells of mobilized peripheral blood (PB) and UCB were cultured in IMDM at a concentration of 1×10
5cells/mL for 11 days.
Thrombopoietin (TPO) 5 ng/mL and 50 ng/mL, flt-3 ligand (FL) 50 ng/mL and stem cell factor
(SCF) 50 ng/mL were used as growth factors. Stromal cells cultured from bone marrow (BM)
mononuclear cells appeared as a single layer of fusiform adherent cells with expression of SH-2
and ASMA. Results: When PB CD34
+cells were cultured with growth factors only, the cell
number increased in TPO 5 ng/mL and 50 ng/mL, TPO and FL, TPO, FL and SCF group
increased upto mean 2.0 (in TPO 5 ng/mL), 2.9 (in TPO 50 ng/mL), 2.4 (TPO and FL), 2.8
(TPO, FL and SCF) fold, and the expression of CD61a were mean 11.5%, 12.5%, 13.2% and
17.0%, respectively. When the stromal cells were added to the growth factors, the cell number
increased upto mean 4.6, 4.9, 9.2 and 68.5 fold and CD61a were expressed in mean 43.1%,
48.4%, 35.6%, and 6.2% of cultured cells. For UCB CD34
+cells, the cell number were increased
upto 7.5, 7.6, 8.2 and 23.6 fold with growth factors and stromal cells. Expression of CD61a were
mean 26.0%, 34.3%, 31.5% and 23.5%, respectively. Conclusion: Stromal cell enhanced the
cellular proliferation and megakaryocytic differentiation of UCB CD34
+cells. The combination
of TPO, FL and SCF increased total cell number upto the highest value but the proportion of
the CD61a
+cells were relatively low. The combination of TPO and FL induced both cellular
proliferation and megakaryocytic differentiation in a large amount. (Korean J Pediatr Hematol Oncol 2004;11:7~16)
Key Words: Umbilical cord blood, Stromal cells, Growth factors, Megakaryocytic differ- entiation
서 론
Gluckman과 Broxmeyer가 처음으로 1988년에 Fanconi 빈혈 환자에서 성공적인 제대혈 이식을 보고한 이후
1)전 세계적으로 2,000례 이상의 비혈 연 혹은 혈연간 제대혈 이식을 시행하였다
2,3). 제 대혈은 공급원이 풍부하고 이식 후 이식편대 숙 주반응이 적은 장점이 있지만 불충분한 세포수와 일부 관련되어 이식 후 생착이 지연되고 특히 혈 소판 회복이 느린 것이 임상적인 제한점이다.
제대혈 이식의 이러한 단점을 극복하기 위하여 액상배양과 여러 가지 성장인자의 조합을 이용한 체외증식을 시도하여 임상응용 단계에 도달하였 다
4). 그러나 현재까지 조혈모세포의 체외증식으 로 수임된 세포 수의 증가 이외에 재생능력을 가 진 세포의 수를 늘리려는 시도는 성공적이지 못 하여, 제대혈의 일부를 체외증식하여 이식한 결과 중성구의 회복기간을 약간 단축하였을 뿐 혈소판 회복은 차이가 없었다
5). 조혈모세포의 체외증식 의 목표는 세포의 분화를 억제하고 증식을 유도 하여 재생능력을 가진 원시 조혈모세포의 수를 증가시키고 또한 특정계열의 세포로 분화시키기 위한 것이다
5). 이를 위하여 가장 효과적인 성장인 자의 조합을 찾고자 하는 연구를 활발히 진행하 고 있지만 조혈모세포에서 여러 가지 계열로 분 화에 필요한 최적의 성장인자의 조합은 아직 확 정되지 않았으며 거핵구와 혈소판 전구체를 증식 시키는 배양조건도 연구자마다 다르다
6,7). 이 연구에서는 제대혈 조혈모세포의 체외배양 에서 기질세포가 골수의 조혈미세환경과 유사한 배양조건을 제공하여 조혈모세포의 증식과 특정 계열로의 분화를 촉진할 것이라는 가설을 세우고 조혈 성장인자와 함께 사람의 골수에서 분리한
기질세포를 이용하여 제대혈 조혈모세포를 배양 하고 거핵구 계열로 분화시키고자 하였다.
재료 및 방법 1) 골수 기질세포의 배양
고형종양 환자에서 골수침범을 확인하기 위하 여 시행한 골수검사에서 정상 골수를 채취하여 Ficoll-Hypaque (density 1.077; Pharmacia Biotech, Uppsala, Sweden)에 조심스럽게 중층한 후 400 g 에서 30분간 원심 분리하여 단핵세포를 분리하였 다. 얻어진 단핵세포를 IMDM으로 재부유하고 37
oC, 5% CO
2에서 1시간 배양하여 바닥에 붙은 세포를 제외하고 상층부의 비유착세포 분획을 얻 었다. 이 세포를 DMEM-LG (Gibco BRL, Gaithers- burg, USA)에 10% 우태아혈청 (Gibco BRL, Gai- thersburg, USA)을 혼합하여 매 4일마다 배지를 교 환하면서 4주간 배양하였다. FACScan (Beckton Dickinson, San Jose, USA) 유세포분석으로 anti- SH-2 (PharMingen, San Diego, USA)와 anti-ASMA (Biomeda, Foster City, USA) 단클론 항체와의 반 응을 검사하며 동시에 anti-CD34 (Beckton- Dickin- son, San Jose, USA)와 anti-CD45 (DAKO, Carpen- teria, USA) 단클론 항체와의 반응은 음성임을 확 인하였다.
2) 말초혈액 CD34
+세포의 분리
관해 상태에 있는 4기 신경모세포종 환자에서
고용량 항암치료와 자가 조혈모세포 이식을 목적
으로 항암제와 granulocyte-colony stimulating factor
(G-CSF)를 이용하여 말초혈액 조혈모세포를 가동
화하였다. 백혈구 분반술로 얻어진 말초혈액 조혈
모세포에서 CliniMACS (Miltenyi Biotech, Bergishi
Gladbach, Germany)를 이용하여 CD34
+세포를 분 리하였다.
3) 제대혈 CD34
+세포의 분리
만삭 정상 분만에서 얻어진 제대혈에서 Ficoll- Hypaque (density 1.077; Pharmacia Biotech, Uppsa- la, Sweden)를 이용하여 단핵세포를 분리하고 MiniMACS CD34 direct purification kit (Miltenyi Biotech, Bergishi Gladbach, Germany)를 사용하여 CD34
+세포를 분리하였다.
4) 기질세포와 성장인자를 이용한 CD34
+세포의 배양과 거핵구 분화
골수의 단핵세포 배양에서 얻어진 기질세포를 trypsin-EDTA (Gibco BRL, Gaithersburg, USA)로 처리하여 분리하고 60Gy의 방사선을 조사시킨 후, 4×10
4/well의 농도로 6 well tissue culture plate 에 한 층으로 분주하였다. 기질세포가 깔려 있는 각 well에 1×10
5의 말초혈액 혹은 제대혈 CD34
+세포가 포함된 2 mL IMDM 용액(Gibco BRL, Gai- thersburg, USA)과 10% 우태아혈청 (Gibco BRL, Gaithersburg, USA)을 넣어 주었다. 각각의 well에 thrombopoietin (이하 TPO, R&D Systems, Minne- apolis, USA) 5 ng/mL와 50 ng/mL, TPO 50ng/mL 와 flt-3 ligand (이하 FL, R & D Systems, Minnea- polis, USA) 50 ng/mL의 조합, TPO 50 ng/ mL와 FL 50 ng/mL, stem cell factor (이하 SCF, R&D Systems, Minneapolis, USA) 50 ng/mL의 조합을 넣 고 배양하였다. 기질세포를 넣지 않고 성장인자의 각 조합만으로 배양하여 기질세포를 이용한 결과 와 비교하였다. 37
oC, 5% CO
2배양기에서 3~4일 간격으로 배지와 성장인자를 절반씩 교환하며 배 양하였다. 배양 11일째에 세포를 수확하여 hemo- cytometer로 세포수를 측정하고 phycoerythrin이 결 합된 mouse anti-human CD61a (PharMingen, San Diego, USA), FITC가 결합된 anti-CD42b (Beck- ton-ickinson, San Diego, USA)와 anti-CD34 (Beck- ton-Dickinson, San Diego, USA) 단클론 항체를 반 응시킨 후 FACScan (Beckton-Dickinson, San Diego,
USA)을 이용하여 유세포분석을 시행하였다.
결 과 1) 골수 기질세포의 배양결과
골수의 단핵세포를 4주간 배양한 결과 균일한 형태의 방추형 세포들이 바닥에 붙어 성장하고 있는 것을 관찰하였다(Fig. 1A). 이 세포들을 기질 세포로 이용하여 말초혈액과 제대혈의 CD34
+세 포를 공동 배양하였다(Fig. 1B). 이 세포들에서 유 세포분석을 시행한 결과 기질세포에서 발현되는 SH-2와 ASMA는 양성이었고 조혈모세포의 표지 자인 CD34와 CD45는 음성으로 나타났다(Fig. 2).
2) 가동화된 말초혈액 CD34
+세포의 배양결과
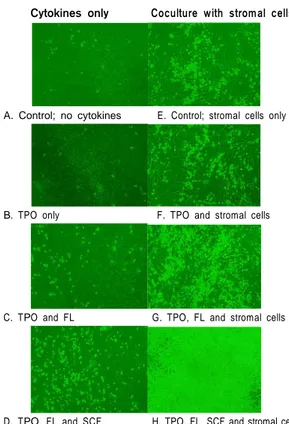
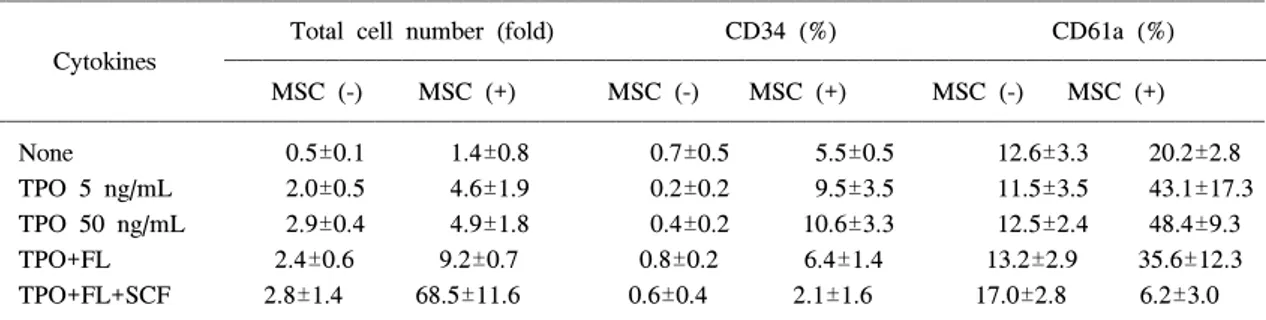
가동화된 말초혈액 3례를 배양하여 검사하였 다. 성장인자만을 첨가하여 액상배양한 경우 세포 수의 증가는 TPO 5 ng/mL과 TPO 50 ng/mL 단독 군, TPO와 FL의 조합, 그리고 TPO, FL과 SCF 조 합에서 각각 평균 2.0 (범위 1.4~2.3), 2.9 (범위 2.4~3.2), 2.4 (범위 2.0~3.0), 그리고 2.8배(범위 1.2~3.8)로 TPO의 농도 차이나 FL 및 SCF의 첨 가로 인한 차이가 없었다. 세포의 크기는 균일하 게 작게 나타났다(Table 1, Fig. 3).
기질세포를 배양에 이용한 경우에는 성장인자 가 첨가되지 않은 대조군에서는 세포수의 증가가 처음 배양당시의 평균 1.4배(범위 0.6~2.2)였고 성장인자의 사용시 TPO 5 ng/mL과 TPO 50 ng/
mL 단독군, TPO와 FL의 조합, 그리고 TPO, FL과 SCF 조합에서 각각 평균 4.6, 4.9, 9.2, 그리고 68.5 배로 세포수가 증가하였다. 세포의 크기는 성장인 자만으로 배양한 경우와 달리 작은 세포에서 중 간 크기 세포, 큰 세포 등 여러 가지로 나타났다 (Fig. 3).
기질세포와의 배양에서 성장인자로 TPO 단독
으로 사용하여 TPO 농도의 증가에 따는 차이를
관찰한 결과 TPO 5 ng/mL에서 세포수가 평균 4.6
배(범위 2.8~6.6)로 증가하여 TPO 50 ng/mL의
Cytokines only Coculture with stromal cells
A. Control; no cytokines E. Control; stromal cells only
B. TPO only F. TPO and stromal cells
C. TPO and FL G. TPO, FL and stromal cells
D. TPO, FL and SCF H. TPO, FL, SCF and stromal cells
Fig. 3. Morphology of the in vitro cultured cells (Phase
contrast microscope with green filter, ×40). PB CD34+ cells were cultured for 11 days with cytokines and BM stromal cells.SH-2 (CD105) ASMA
CD34 CD45
Fig. 2. Flow cytometric analysis of BM stromal cells.
They express markers of stromal cells like SH-2 and ASMA, but not CD34 and CD45 of hem- atopoietic stem cells.
A B
Fig. 1. Stromal cells from human bone marrow (Phase contrast microscope with green filter). (A) Adherent fusiform
cells after 4 weeks of culture (×100). (B) Stromal cells coculture with umbilical cord blood CD34+ cells for 11 days (×40).Table 1. Total Cell Number (fold) and Percentage of PB CD34
+ Cells and CD61a+ Megakaryocytic Cells after 11 Days of Culture with Cytokines and Stromal Cells (n=3)
Total cell number (fold) CD34 (%) CD61a (%)
Cytokines
MSC (-) MSC (+) MSC (-) MSC (+) MSC (-) MSC (+)
None 0.5±0.1 1.4±0.8 0.7±0.5 5.5±0.5 12.6±3.3 20.2±2.8
TPO 5 ng/mL 2.0±0.5 4.6±1.9 0.2±0.2 9.5±3.5 11.5±3.5 43.1±17.3
TPO 50 ng/mL 2.9±0.4 4.9±1.8 0.4±0.2 10.6±3.3 12.5±2.4 48.4±9.3
TPO+FL 2.4±0.6 9.2±0.7 0.8±0.2 6.4±1.4 13.2±2.9 35.6±12.3
TPO+FL+SCF 2.8±1.4 68.5±11.6 0.6±0.4 2.1±1.6 17.0±2.8 6.2±3.0
TPO; 50 ng/mL, FL; 50 ng/mL, SCF; 50 ng/mL
Abbreviations: PB; mobilized peripheral blood stem cells, FL; flt-3 ligand, MSC; marrow stromal cells
4.9배(범위 2.9~6.1)와 차이가 없었다(Table 1). 기
질세포와 함께 FL과 SCF를 사용한 경우에 세포 수의 증가가 현저하였는데 TPO와 FL의 조합에서 는 평균 9.2배(범위 8.3~9.7)였고 SCF까지 첨가한 경우 평균 68.5배(범위 55.7~78.2)로 큰 증식을 보였다(Table 1).
3) 말초혈액 CD34
+세포에서 거핵구 표지자 CD61a의 발현
성장인자만을 사용한 경우 CD61a의 발현 비율 은 TPO 5 ng/mL과 TPO 50 ng/mL 단독군, TPO와 FL의 조합, 그리고 TPO, FL과 SCF 조합에서 각 각 평균 11.5% (범위 7.6~14.2), 12.5% (범위 9.
7~13.9), 13.2% (범위 10.0~15.4), 그리고 17.0%
(범위 14.2~19.8)로 성장인자를 넣지 않은 대조군 의 평균 12.6% (범위 9.1~15.6)와 차이가 없었다 (Table 1).
골수의 기질세포에서 배양한 경우 세포수의 증 가와 함께 CD61a의 발현도 대조군의 평균 20.2%
(범위 17.9~23.0)에 비하여 증가하는 경향이 나타 났다. TPO 5 ng/mL와 50 ng/mL를 이용한 경우 CD61a의 발현이 각각 평균 43.1% (범위 24.1~
57.9)와 평균 48.4% (범위 41.9~59.0)였고 TPO 50 ng/mL에 FL을 첨가한 경우 세포수의 증가가 평균 9.2배(범위 8.3~9.7)이면서 CD61a의 비율이 35.6%
로 나타났다. TPO, FL과 함께 SCF를 사용한 경우
에는 세포수의 증가가 평균 68.5배(범위 55.7~
78.2)이고 CD61a를 발현하는 세포의 증가도 가장 많았지만 CD61a의 발현비율은 평균 6.2% (범위 4.3~9.6)로 낮아서 거핵구 이외 다른 계통의 세포 가 많이 증가한 것으로 생각할 수 있었다(Table 1).
CD34 항원의 발현은 배양 시작당시 평균 95%
이상의 값을 보였지만 성장인자만을 사용하여 배 양한 경우 11일째에 대조군의 0.7%와 비슷하거나 더 감소하여 TPO 5 ng/mL과 TPO 50 ng/mL 단독 군, TPO와 FL의 조합, 그리고 TPO, FL과 SCF 조 합에서 각각 평균 0.2% (범위 0.1~0.4), 0.4% (범 위 0.2~0.5), 0.8% (범위 0.5~0.9), 그리고 0.6%
(범위 0.3~1.0)의 비율이었다. 기질세포가 존재하 는 경우에는 성장인자를 사용하지 않은 대조군에 서는 CD34 항원의 발현비율이 평균 5.5% (범위 5.0~5.9)이고 TPO 5 ng/mL와 50 ng/mL를 사용하 는 경우 각각 평균 9.5% (범위 5.5~12)와 10.6%
(범위 7.5~14.1)로 상대적으로 높은 값이 나타났 다. TPO와 FL의 조합에서는 CD34 항원의 발현비 율이 평균 6.4% (범위 5.0~7.8)이었고 TPO, FL 및 SCF의 조합에서는 평균 2.1% (범위 1.0~3.9) 로 가장 낮았지만 세포수는 평균 5.5배(범위 4.4~
6.2)로 증가하여 절대수가 가장 높았다(Table 1).
4) 제대혈 CD34
+세포의 배양결과
만삭 정상 분만에서 얻은 제대혈 3례를 배양하
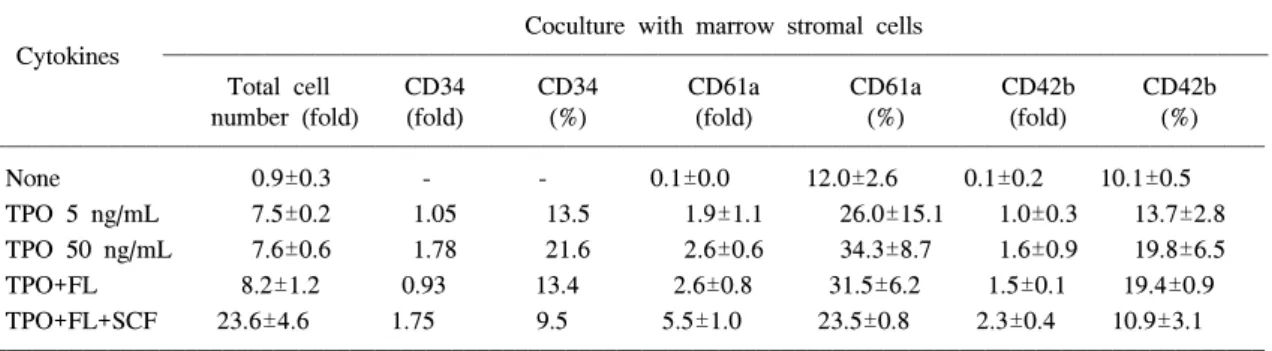
Table 2. Total Cell Number (fold) and Percentage of Umbilical Cord Blood CD34
+ Cells and CD61a+, CD42b+ Megakaryocytic Cells after 11 Days of Culture with Cytokines and Stromal Cells (n=3)
Coculture with marrow stromal cells
Cytokines
Total cell CD34 CD34 CD61a CD61a CD42b CD42b
number (fold) (fold) (%) (fold) (%) (fold) (%)
None 0.9±0.3 - - 0.1±0.0 12.0±2.6 0.1±0.2 10.1±0.5
TPO 5 ng/mL 7.5±0.2 1.05 13.5 1.9±1.1 26.0±15.1 1.0±0.3 13.7±2.8
TPO 50 ng/mL 7.6±0.6 1.78 21.6 2.6±0.6 34.3±8.7 1.6±0.9 19.8±6.5
TPO+FL 8.2±1.2 0.93 13.4 2.6±0.8 31.5±6.2 1.5±0.1 19.4±0.9
TPO+FL+SCF 23.6±4.6 1.75 9.5 5.5±1.0 23.5±0.8 2.3±0.4 10.9±3.1
TPO; 50 ng/mL, FL; 50 ng/mL, SCF; 50 ng/mL
여 검사하였다. 성장인자와 기질세포를 이용하여 11일간 배양 후 초기의 CD34
+세포에 대비하여 기질세포만을 이용한 대조군에서는 세포수의 증 가배수가 평균 0.9배(범위 0.6~1.1)였다(Table 2).
성장인자로 TPO, FL, 그리고 SCF 조합을 사용 한 경우 평균 23.6배(범위 18.4~27.2배)의 가장 높은 증식을 보였으며, TPO 5 ng/mL에서 평균 7.5배(범위 7.3~7.8배), TPO 50 ng/mL에서 평균 7.6배(범위 7.0~8.2배), TPO와 FL을 병합하여 사 용한 경우에는 평균 8.2배(범위 6.9~9.3배) 등으 로 세포수의 증가에서 큰 차이가 없었다(Table 2).
5) 제대혈 CD34
+세포에서 거핵구 표지자 CD61a와 CD42b의 발현
배양된 세포에서 거핵구 표지자의 발현은 초기 거핵구 표지자인 CD61a의 경우 TPO 5 ng/mL과 TPO 50 ng/mL 단독군, TPO와 FL의 조합, 그리고 TPO, FL과 SCF 조합에서 각각 평균 26.0% (범위 16.9~43.5%), 34.3% (범위 24.6~41.3%), 31.5% (범 위 24.8~36.9%), 23.5% (범위 22.6~24.2%)로서 TPO 50 ng/mL 군에서 최고치를 보였지만 성장인 자의 조합에 따른 차이가 거의 없었다(Table 2).
후기 거핵구 표지자인 CD42b의 발현은 제대혈 2례에서 검사하여 TPO 5 ng/mL과 TPO 50 ng/mL 단독군, TPO와 FL의 조합, 그리고 TPO, FL과 SCF 조합에서 각각 평균 13.7% (범위 10.9~
16.4%), 19.8% (범위 13.2~26.3%), 19.4% (범위 18.5~20.3%), 10.9% (범위 7.8~14.0%) 정도로 역 시 큰 차이는 없었으나 TPO 50 ng/mL 군과 TPO, FL 군에서 다소 높은 값을 보였으며 TPO와 FL, SCF를 병합한 경우에 그 비율이 가장 낮았다 (Table 2).
CD34 항원의 발현은 제대혈 3례 중 1례에서 검 사하여 TPO 50 ng/mL 군에서 21.6%로 가장 높았고 TPO 5 ng/mL 군에서는 13.5%, FL과 TPO의 조합에 서는 13.4%의 비율이었다. FL, TPO와 SCF의 조합 에서는 9.5%로 그 비율이 가장 낮았다(Table 2).
고 찰
조혈모세포 이식은 여러 가지 악성 및 비악성 질환에서 점차 적응증을 넓혀가고 있으며 조혈모 세포의 공급원도 골수에서 말초혈액과 제대혈로 확장되었다
5). 조혈모세포 이식에서 생착의 지연 이나 혈소판 감소증의 지속은 이식 후 유병률을 높이는 심각한 문제이다. 백혈구 감소기간이 길어 질수록 여러 가지 감염의 위험이 높고 혈소판 감 소에 따른 출혈을 예방하기 위하여 반복적으로 혈소판 수혈이 필요하다
6). 조혈모세포의 체외증 식은 이러한 문제점을 해결할 수 있는 방법으로 기대를 모으고 있다.
조혈미세환경에서 기질세포는 줄기세포의 생성
과 분화를 유지하는 중요한 외부신호를 제공하는 데 이러한 신호는 아직 확인되지 않은 물질이나 상호작용에 의한 조절일 가능성이 있다
8). 이러한 물질은 아마도 기질세포 표면의 성장인자, 유착분 자, 혹은 세포외간질 등인 것으로 추측할 수 있으 며 따라서 조혈모세포와 공동배양에서 기질세포 와의 직접 접촉이 세포성장에 중요한 인자임을 설명할 수 있다
9). 사람의 조혈모세포 증식을 위한 기질세포와의 공동배양에서 생쥐의 기질세포를 주로 사용하여 왔으나
10,11), 감염의 우려가 있고 종특이성의 차이 때문에 그 안전성이 논란이 되 었다. 이에 대한 대안으로 사람의 기질세포를 이 용하기 시작하였는데, 환자 자신이나 조혈모세포 의 공여자에서 얻은 기질세포는 감염의 위험을 줄일 수 있다
12). 조혈모세포의 체외배양에서 기질 세포의 이용은 여러 연구자들이 보고하고 있으며 최근에는 유전자 변형을 통해 사람의 기질세포주 를 확립하기에 이르렀다
13). 앞으로 조혈모세포의 성장과 분화에 필수적인 성장인자를 생성하는 유 전자를 도입한 형질변환 기질세포주를 개발한다 면 조혈모세포의 체외배양에 이용할 수도 있을 것이다.
조혈모세포의 체외배양을 위해서 여러 가지 성 장인자들을 이용하고 있는데 이 연구에서는 조혈 모세포의 초기 단계와 거핵구 생성에 핵심적으로 작용하는 것으로 알려진 TPO, FL과 SCF 등을 성 장인자로 선택하였다
14). TPO는 c-mpl ligand로서 여러 연구자들이 거핵구 형성의 조절에 중추적인 역할을 담당하는 것으로 보고하였다. TPO는 거핵 구에 우세하게 작용하지만 다른 계열의 분화와 증식에도 작용하므로
15), 혈소판 생성 이외에도 골 수구 계열과 적혈구 계열에 주목할 만한 효과가 있다. FL은 SCF와 유사하게 초기 조혈과정을 조 절하는 중심적인 역할을 한다
16). SCF의 수용체인 c-kit
17)는 비록 세포유형에 따라 발현량의 차이가 있지만 대부분의 조혈모세포가 이를 발현한다
18). 어떤 성장인자 단독의 역할에 대한 결론을 내리 기는 어렵지만 조혈모세포의 체외배양에 효과적 인 조합의 대부분에서 SCF를 포함한다.
이 연구에서는 골수의 조혈미세환경과 유사한 배양조건을 제공하여 조혈모세포의 증식과 특정 계열로의 분화를 촉진하고자 성장인자와 함께 사 람의 골수에서 분리한 기질세포를 이용하여 말초 혈액과 제대혈의 CD34
+세포를 배양하고 거핵구 계열로 분화시켰다. 가동화된 말초혈액 CD34
+세 포를 성장인자 단독이나 성장인자와 기질세포를 함께 사용하여 배양하면서 기질세포의 유무가 세 포증식에 미치는 영향을 관찰하고, 제대혈 CD34
+세포를 골수의 기질세포의 존재하에 TPO, flt-3와 SCF 등의 성장인자를 투여하면서 체외배양 하였 다.
본 연구자는 예비실험에서 제대혈 CD34
+세포 를 골수 기질세포와 공동배양하는 경우 성장인자 를 첨가하지 않아도 세포의 생존과 증식이 가능 함을 관찰하였다(자료제시 안 됨). 반면에 기질세 포가 없이 성장인자만을 투여한 경우에서는 세포 의 생존이 미약하였고 세포증식이 일어나지 않았 다. 이런 결과를 바탕으로 하여 이번 실험에서는 가동화된 말초혈액 CD34
+세포에서 기질세포의 유무가 세포증식에 미치는 영향을 관찰하고 제대 혈에서 분리한 CD34
+세포를 성장인자와 기질세 포를 이용하여 배양하였다.
이번 연구에서 가동화된 말초혈액 CD34
+세포 를 대상으로 기질세포 없이 성장인자만을 사용하 여 배양한 경우 세포증식 정도가 평균 2.0~2.9배 로 매우 낮았고 거핵구로의 분화도 미약하였다.
또한 세포의 크기가 고르게 작은 것을 관찰하였
다. 이에 반하여 기질세포와 공동배양한 경우에는
세포수의 증가가 평균 4.6~68.5배에 이르렀으며
거핵구 표지자를 가진 세포의 비율도 성장인자의
조합에 따라 평균 6.2~48.4%까지 증가한 것을 관
찰하였다. 또한 세포의 크기에 차이가 있어서 연
구자 임의로 작은(small) 세포-중간(medium-sized)
세포 - 큰(large) 세포로 분류를 하고 앞으로 그 의
미에 대한 검토가 필요하다. TPO, FL과 SCF를 병
합하여 사용한 경우 전체 세포수의 증가와 CD34
+세포의 증가배수가 가장 높았으며 CD61a
+세포
의 증가 또한 최고치를 보였으나 거핵구가 차지
하는 비율은 가장 낮았다. 이 경우 거핵구 이외 골수구나 적혈구 등 다른 계열의 세포가 많이 생 기는 것으로 추정할 수 있으며 향후 이에 대한 검 사가 필요할 것이다. TPO와 FL을 사용한 경우 세 포수의 증가는 SCF 군보다 낮았지만 거핵구의 비 율은 더 높아서 세포수의 증가와 함께 거핵구로 의 분화유도에 유리한 것으로 생각된다. 실제로 임상에서 체외 배양된 세포를 이식에 사용하는 경우 세포수의 증가와 함께 목표한 특정 계열의 증식을 고려해야 할 것이므로 목적에 따라 적정 한 성장인자의 조합을 선택해야 할 것으로 생각 된다. 이 실험에서는 TPO를 다른 성장인자와 조 합할 때 50 ng/mL를 사용하였는데 TPO 단독 사 용에서 5 ng/mL와 50 ng/mL의 차이가 거의 없었 으므로 성장인자 조합에서 TPO의 사용량을 줄일 수 있는 가능성이 있다.
또한 제대혈 CD34
+세포에서는 성장인자와 기 질세포를 이용하여 배양한 결과 세포수의 증가가 평균 7.6~23.6배로서 말초혈액의 경우와 큰 차이 는 없었지만 최고 증가치는 상당한 차이가 있었 다. 성장인자에 대한 반응은 말초혈액에서와 마찬 가지로 TPO, FL, 그리고 SCF 조합을 사용한 경우 가장 높게 증식되었지만 말초혈액의 평균 68.5배 에 비하여 낮은 23.6배 정도로 증가하였다. 같은 조합에서 거핵구 표지자의 발현은 역시 다른 성 장인자를 사용한 군에 비하여 약간 낮은 편이었 다. 실제로 임상에서 체외배양된 세포를 이식에 사용하는 경우 CD34
+세포수의 증가와 함께 목 표한 특정 계열의 증식을 고려해야 할 것이므로 목적에 따라 적정한 성장인자의 조합을 선택해야 할 것으로 생각된다.
제대혈의 결과와 말초혈액 CD34
+세포의 배양 결과의 차이를 설명할 수 있는 것은 첫째, 제대혈 의 반응이 말초혈액과 다르거나, 둘째, 배양액에 대한 세포농도가 적정 수준에 미치지 못하였거나, 셋째, 배양기간이 충분하지 못하기 때문일 수 있 다. 앞으로 반복실험을 통하여 배양액에 대한 세 포의 농도와 성장인자의 조합, 그리고 배양기간의 적정성에 대한 검토가 필요할 것으로 생각된다.
또한 이 실험에서 정상인이 아닌 종양환자에서 항암제와 G-CSF로 가동화한 말초혈액 CD34+ 세 포를 사용한 것이 결과에 영향을 줄 수 있을 것이 다.
이번 실험 결과의 한계로 지적할 수 있는 것은 전반적인 배양조건이 적절한지 여부의 확인이다.
조혈모세포의 체외배양 결과는 보고마다 큰 차이 가 있지만 이번 연구에서 세포수의 증가폭은 상 당히 낮은 편이다. 또한 기질세포가 없는 경우 조 혈모세포의 성장인자에 대한 반응이 거의 없는 것도 의외의 결과라고 할 수 있겠다. 여러 검체에 서 반복적으로 배양을 시도하여 결과를 분석할 필요성이 있다.
이 연구에서와 같이 기질세포를 이용하여 체외 배양한 조혈모세포를 실제로 이식에 사용하는 경 우 한 가지 고려할 사항은 기질세포와 조혈모세 포의 분리이다. 세포를 분리하기 위하여 trypsin이 나 collagenase를 사용하는 경우 조혈모세포에 손 상을 초래할 수 있으므로 신중하게 처리해야 할 것이다. 본 연구자는 기질세포에서 분비되는 수용 성 물질의 역할을 확인하기 위하여 Schattner 등
19)이 제시한 것과 같이 기질세포의 배양시 상층액 을 사용하여 배양을 시도하고 있으며, 좋은 결과 가 있을 경우 실제로 임상에 적용하는 적절한 대 안으로 사용할 수 있을 것이다.
현재까지 성장인자의 조합을 이용하여 체외증
식된 제대혈 조혈모세포가 세포수의 증가에도 불
구하고 이식 후 혈소판의 회복을 단축시키지 못
하는 것은 생착에 관련된 조혈모세포의 특성이
변화되기 때문이라고 추정할 수 있다. 지금까지
몇몇 연구에서 체외증식에 따른 거핵구의 세포고
사
20), 이주능의 감소
21,22)등의 변화를 관찰하였고
또한 기질세포와의 공동배양이 거핵구의 분화를
억제하여 증식능을 유지함을 보고하였다
23).
이 연구에서는 제대혈에서 거핵구 계열의 증식
과 분화를 목표로 하여 세포수의 증가와 거핵구
표지자의 발현을 관찰하였는데 실제로 임상적으
로 적용을 위해서는 배양된 조혈모세포의 생착능
력과 분화단계를 확인하는 과정이 필요하다. 또한
기질세포의 조혈지지 작용이 나타나는 기전을 분 석하여 이를 조혈모세포 배양에 응용하려는 노력 이 병행되어야 할 것이다.
요 약
목적: 조혈 성장인자와 함께 사람의 골수에서 분리한 기질세포를 이용하여 제대혈 조혈모세포 를 배양하고 거핵구 계열로 분화시키고자 하였다.
방법: 가동화된 말초혈액 조혈모세포와 제대혈 에서 CD34
+세포를 분리하여 10% 우태아혈청이 포함된 IMDM에 1×10
5/mL의 농도로 넣고 성장 인자로는 TPO 5 ng/mL와 50 ng/mL, FL 50 ng/mL, 그리고 SCF 50 ng/mL를 사용하여 11일간 배양하 였다. 기질세포로는 골수의 단핵세포를 DMEM- LG에서 4주간 배양하여 SH-2와 ASMA의 발현이 나타나는 유착세포를 이용하였다.
결과: 가동화된 말초혈액 CD34
+세포에 성장인 자만을 사용하여 배양한 경우 세포수의 증가는 TPO 5 ng/mL와 50 ng/mL 단독군, TPO와 FL의 조 합, 그리고 TPO, FL과 SCF의 조합에서 각각 평균 2.0, 2.9, 2.4와 2.8배 정도이고 CD61a의 발현비율 은 평균 각각 11.5, 12.5, 13.2, 그리고 17.0%로서 TPO의 농도나 FL 및 SCF의 첨가로 인한 차이가 없었다. 성장인자와 함께 기질세포를 배양에 이용 한 경우에는 세포수의 증가가 TPO 5 ng/mL와 50 ng/mL 단독군, TPO와 FL의 조합, 그리고 TPO, FL과 SCF의 조합에서 각각 평균 4.6, 4.9, 9.2와 68.5배였고, CD61a는 각각 평균 43.1, 48.4, 35.6와 6.2%까지 발현하였다. 제대혈 CD34
+세포에서 성장인자와 기질세포를 함께 이용하여 배양한 경 우 TPO 5 ng/mL와 50 ng/mL 단독군, TPO와 FL의 조합, 그리고 TPO, FL과 SCF의 조합에서 세포수 가 각각 평균 7.5, 7.6, 8.2와 23.6배로 증가하였다.
배양된 세포에서 CD61a의 발현은 TPO 5 ng/mL와 50 ng/mL 단독군, TPO와 FL의 조합, 그리고 TPO, FL과 SCF의 조합에서 각각 평균 26.0, 34.3, 31.5 와 23.5%였다.
결론: 성장인자와 함께 기질세포를 사용한 경우
말초혈액과 제대혈 CD34
+세포의 증식과 거핵구 로의 분화가 활발하였다. 말초혈액과 제대혈의 CD34
+세포의 체외배양에서 기질세포와 함께 성 장인자 TPO, FL과 SCF를 사용한 경우 전체 세포 수와 CD34
+세포수가 가장 많이 증가하였지만 CD61a
+를 발현하는 거핵구 계열의 비율은 다른 조합과 비교할 때 가장 낮았다. TPO와 FL을 사용 한 경우 세포수의 증식과 함께 CD61a
+세포의 비 율이 가장 높아서 거핵구로의 분화에 유리할 것 으로 판단되었다.
참 고 문 헌
1. Gluckman E, Broxmeyer HA, Auerbach AD, Friedman HS, Douglas GW, Devergie A, et al. Hematopoietic reconstitution in a patient with Fanconi's anemia by means of umbilical-cord blood from an HLA-identical sibling. N Engl J Med 1989;321:1174-8
2. Barker JN, Davies SM, DeFor T, Ramsay NK, Weis- dorf DJ, Wagner JE. Survival after transplantation of unrelated donor umbilical cord blood is comparable to that of human leukocyte antigen-matched unrelated donor bone marrow: Results of a matched-pair analy- sis. Blood 2001;97:2957-61
3. Burgio GR, Gluckman E, Locatelli F. Ethical reapprai- sal of 15 years of cord-blood transplantation. Lancet 2003;361:250-2
4. Fibbe WE, Noort WA, Schipper F, Willemze R. Ex vivo expansion and engraftment potential of cord blood-derived CD34+ cells in NOD/SCID mice. Ann N Y Acad Sci 2001;938:9-17
5. McNiece I, Briddell R. Ex vivo expansion of hema- topoietic progenitor cells and mature cells. Exp Hematol 2001;29:3-11
6. Pick M, Eldor A, Grisaru D, Zander AR, Shenhav M, Deutsch VR. Ex vivo expansion of megakaryocyte progenitors from cryopreserved umbilical cord blood.
A potential source of megakaryocytes for transplanta- tion. Exp Hematol 2002;30:1079-87
7. Ryu KH, Seoh JY, Chung WS, Kie JH, Shin HY, Ahn HS. Stem Cell Factor Inhibits Premature Senescence of Magakaryocytes during ex vivo Expansion of Human Cord Blood CD34+ Cells Using Thrombo- poietin. Korean J Pediatr Hematol-Oncol 200;7:212-23 8. Dorshkind K. Regulation of hemopoiesis by bone mar- row stromal cells and their products. Annu Rev Imm-
unol 1990;8:111-37
9. Moore MA, Schneider JG, Shapiro F, Bengala C. Ex vivo expansion of CD34+ hematopoietic progenitors.
Prog Clin Biol Res 1994;389:217-28
10. Xu MJ, Tsuji K, Ueda T, Mukouyama YS, Hara T, Yang FC, et al. Stimulation of mouse and human primitive hematopoiesis by murine embryonic aorta- gonad-mesonephros-derived stromal cell lines. Blood 1998;92:2032-40
11. Kawada H, Ando K, Tsuji T, Shimakura Y, Nakamura Y, Chargui J, et al. Rapid ex vivo expansion of human umbilical cord hematopoietic progenitors using a novel culture system. Exp Hematol 1999;27:904-15 12. Loeuillet C, Bernard G, Remy-Martin J, Saas P, Herve
P, Douay L, et al. Distinct hematopoietic support by two human stromal cell lines. Exp Hematol 2001;29:
736-45
13. Kawano Y, Kobune M, Yamaguchi M, Nakamura K, Ito Y, Sasaki K, et al. Ex vivo expansion of human umbilical cord hematopoietic progenitor cells using a coculture system with human telomerase catalytic subunit (hTERT)-transfected human stromal cells.
Blood 2003;101:532-40
14. Hassan HT, Zander A. Stem cell factor as a survival and growth factor in human normal and malignant hematopoiesis. Acta Haematol 1996;95:257-62 15. Kaushansky K. Thrombopoietin: Biological and pre-
clinical properties. Leukemia 1996;10:S46-8 16. Broxmeyer HE, Lu L, Cooper S, Ruggieri L, Li ZH,
Lyman SD. Flt3 ligand stimulates/costimulates the growth of myeloid stem/progenitor cells. Exp Hematol 1995;23:1121-9
17. Galli SJ, Zsebo KM, Geissler EN. The kit ligand, stem cell factor. Adv Immunol 1994;55:1-96
18. Brandt J, Briddell RA, Srour EF, Leemhuis TB, Hoff- man R. Role of c-kit ligand in the expansion of human hematopoietic progenitor cells. Blood 1992;79:634-41 19. Schattner M, Green D, Cohen I. Stromal-conditioned medium synergizes with thrombopoietin in stimulating megakaryocytopoiesis. Stem Cells 1998;16:61-5 20. Ryu KH, Chun S, Carbonierre S, Im SA, Kim HL,
Shin MH, et al. Apoptosis and megakaryocytic differ- entiation during ex vivo expansion of human cord blood CD34+ cells using thrombopoietin. Br J Hae- matol 2001;113:470-8
21. Drayer AL, Sibinga CT, Blom NR, De Wolf JT, Vellenga E. The in vitro effects of cytokines on expansion and migration of megakaryocyte progeni- tors. Br J Haematol 2000;109:776-84
22. Mathur A, Hong Y, Martin J, Erusalimsky J. Mega- karyocytic differentiation is accompanied by a reduction in cell migratory potential. Br J Haematol 2001;112:459-65
23. Zweegman S, Veenhof MA, Huijgens PC, Schuurhuis GJ, Drager AM. Regulation of megakaryocytopoiesis in an in vitro stroma model: preferential adhesion of megakaryocytic progenitors and subsequent inhibition of maturation. Exp Hematol 2000;28:401-10