서 론
어류의 초기생활사에 대한 연구는 분류학적으로 구분이 어 려운 유사종 사이에 종을 구분하거나 유연관계를 연구하는 데 있어 매우 유용하며, 특히 발생과정 중에 드러난 많은 형질 들은 근연종들의 분류학적인 유연관계를 밝히는 데 중요한 기 초자료를 제공하게 된다(Blaxter, 1974; Lee et al., 2013).
강담돔(Oplegnathus punctatus)은 농어목(Perciformes) 돌돔 과(Oplegnathidae) 돌돔속(Oplegnathus)에 속하는 어류로 우 리나라에는 강담돔과 돌돔, O. fasciatus 2종이 분포하고 있으 며, 주로 남해안과 제주도, 일본 중부이남 및 동중국해 연안에 서식하고 있다(Kim et al., 2005).
돌돔과 어류에 대한 국내연구로는 돌돔의 난 발생 및 자치 어기 형태(Go and Kim, 1992), 수온 및 광주기 조절에 의한 산란유도(Jung et al., 1998), 성 분화와 생식소 발달(Kim,
1998), 성 성숙 유도(Kim et al., 2000), 초기 성장과 생존율 (Hwang et al., 2005) 및 돌돔과 강담돔의 자연교잡종(Kwun and Kim, 2010) 등이 수행되었고, 국외 연구로는 이빨의 발달 과정(Kakizawa et al., 1980), 꼬리지느러미의 골격(Ishiguro and Nishida, 1998) 및 어미사육 연구(Kawabe and Kimura, 2007) 등이 수행되었다.
강담돔은 단기간 양식을 목표로 높은 소득을 올릴 수 있는 어종으로 각광받고 있으며(Kim, 2007), 돌돔에 이어 해상가두 리 양식 기술개발에 대한 연구가 진행되고 있으나 자치어기 형태 발달에 대한 자료가 부족한 실정이다. 본 연구에서는 강 담돔의 수정난을 부화시켜 사육하면서 난 발생 과정 및 자치 어의 발달 단계별 형태 변화를 조사하였다.
재료 및 방법
1. 실험어 확보 및 채란유도
본 연구에 사용된 실험어는 2002년 5월 경상남도 통영시 한 ISSn: 1225-8598(Print), 2288-3371(online)
—71 — http://www.fishkorea.or.kr
강담돔 (Oplegnathus punctatus)의 난 발생 및 자치어 형태발달
박재민·이성훈1·윤성민2·나해춘3·한경호1,*
경상북도 토속어류산업화센터, 1전남대학교 양식생물학전공, 2경상북도 민물고기연구센터, 3세보수산
Egg Development and Morphology of Larvae and Juveniles of Spotted Knifejaw, Oplegnathus punc
tatus by Jae Min Park, Sung Hun Lee1, Seong Min Yun2, Hae Choon Na3 and Kyeong Ho Han1,* (gyeongsangbuk- do native fish Business center, uiseong 769-921, Korea; 1chonnam national university, Department of aqualife Science, yeosu 550-749, Korea; 2gyeongsangbuk-Do research center for freshwater fishes, uljin 767-873, Korea; 3Sebo Susan, tongyeong 650-942, Korea)
ABSTRACT Morphological changes in eggs and larvae of spotted knifejaw, Oplegnathus punc
tatus were observed by cultivation of fertilized eggs to juvenile stage. Fertilized egg was demersal and transparent. The egg has one oil glouble and its size ranged 1.01~1.15mm(1.08±0.10mm;
mean±SD, n=20). Culture chambers were maintained 22.5~24.5°C in water temperature and 33.5
~34.5ppt in salinity. It took 25mins from fertilization to the blastoderm stage, 55mins to 2 cell stage and 20hrs 50mins to hatching. Newly hatched yolksac larvae ranged 2.59~3.02mm(2.81±0.25mm, n=5) in total length, and mouth and anus were not still open. Yolk and oil globule were absorbed 2 days after hatching(DAH). Flexion larvae in 12 DAH ranged 3.02~3.07mm TL(3.04±0.04mm), and the caudal fin grew to fan-shaped, melanophores appeared on top of digestive canal.
Key words: Egg development, juvenile, larvae, Oplegnathus punctatus
* Corresponding author: Kyeong Ho Han Tel: 82-61-659-7163, Fax: 82-61-659-7169, E-mail: [email protected]
accepted: april 24, 2015
산면에 위치한 한산도와 추봉도 연안에서 정치망에 어획된 어 미(전장 24.5~26.7cm, 평균 25.7±0.91cm) 5개체를 산소비 닐 포장하여 실험실로 운반하였다.
성숙한 암컷 3마리는 복부를 압박하여 채란하였고, 수컷 2 마리의 복부를 압박하여 채정한 뒤 건식법으로 인공수정 시켰 다.
2. 난 발생 및 자치어의 형태발달
수정란은 1,000mL 유리 비이커에 수용하였고, 에어레이션 으로 약하게 산소를 공급시켜 주었다. 사육수온은 22.5~24.5
°C(평균 23.0±0.05°C)였고, 염분농도는 33.5~34.5‰(평균 34.0±0.05‰)의 범위를 유지시켜 주었다. 난 발생 과정 관찰 을 위해 수정란 20개를 무작위로 채취하여 실체현미경(Nikon SMZ18, Japan)을 이용하여 관찰하였으며, 난의 크기는 0.01 mm까지 측정하였다.
부화 직후의 자어는 폴리프로필렌(PP, Polypropylene) 원 형수조(100 ×50 ×80cm)에 수용하여 지수식으로 사육하였 고, 에어레이션으로 산소를 공급시켜 주었다. 자치어의 먹이 는 난황 흡수 후부터 부화 후 12일까지 로티퍼(Brachionus rotundiformis)를 mL당 10~15개체 공급하였고, 12일부터 25 일까지 알테미아(Artemia sp.)를 공급하였으며, 25일부터 배합 사료(Lovelarva, Japan)를 혼합 공급하였다.
자치어의 형태발달은 부화 직후부터 1일마다 10마리씩 마 취제(MS-222, Ethyl 3-aminobenzoate methanesulfonate, Sig- ma Aldrich Co., St. Louis, USA)를 이용하여 마취시킨 후 어 체의 각 부위를 만능투영기(Nikon JP V-12B, Japan)로 0.01 mm까지 측정 및 관찰하였다. 형태발달 단계는 Kendall(1984) 에 따라 구분하였다.
결 과
1. 난의 형태
인공수정 된 난은 무색 투명한 분리 부성란이었고, 형태는 구형으로 1개의 유구를 가지고 있었으며, 난경은 1.01~1.15 mm(평균 1.08±0.10mm, n=20)였다.
2. 난 발생 과정
수정란은 수정 후 물을 흡수하면서 난이 확대되었고, 수정 후 25분에는 난막과 난황이 분리되면서 배반이 형성되었다 (Fig. 1A). 수정 후 55분에는 난할이 2개로 분리되어 2세포기 에 달하였고(Fig. 1B), 수정 후 1시간 15분에는 같은 크기의 난할이 4개로 분리되면서 4세포기에 달하였다(Fig. 1C). 수정
후 1시간 45분에는 8세포기에 달하였고(Fig. 1D), 수정 후 2시 간 30분에는 16세포기에 달하였다(Fig. 1E). 수정 후 3시간에 는 32세포기에 달하였고(Fig. 1F), 수정 후 3시간 30분에는 64 세포기(Fig. 1G)에 달하였으며, 4시간 10분에는 128세포기에 달하였다(Fig. 1H).
수정 후 4시간 59분에는 상실기에 달하였고(Fig. 1I), 6시간 22분에는 포배기에 달하였으며(Fig. 1J), 8시간 30분에는 배반 엽이 난황의 2/3를 덮어 내려와 중기 낭배기에 달하였다(Fig.
1K). 수정 후 11시간 40분에는 배체가 발달하면서 안포와 4개 의 근절이 형성되었다(Fig. 1L).
수정 후 13시간 40분에는 근절이 8개로 증가하였고, 눈에는 렌즈가 형성되었으며, Kuffer’s vesicle이 나타났다(Fig. 1M).
수정 후 15시간 20분에는 근절이 11~12개로 증가하였고, 뇌 가 분화하였다. 흑색소포는 두부 뒷부분과 근절 주위에서 꼬 리부분까지 침착하였고, 유구 주변에는 나뭇가지 모양의 흑색 소포가 3개 침착되었다(Fig. 1N). 수정 후 16시간 30분에는 근 절이 16~17개로 증가하였고, 유구 주위에는 난황물질이 과립 상으로 존재하였으며, 유구에는 나뭇가지 모양의 흑색소포가 5개로 증가하였다(Fig. 1O).
수정 후 18시간에는 심장이 분화하기 시작하였고, 이포가 출현하였으며, Kuffer’s vesicle이 소실되었다. 이 시기의 근절 수는 19~20개로 증가하였고, 배체에는 나뭇가지 모양의 흑 색소포가 넓게 침착되었으며, 꼬리에는 막으로 된 지느러미가 형성되었다(Fig. 1P). 수정 후 20시간 50분에는 꼬리가 난황으 로부터 분리되었고, 이 시기의 근절 수는 27개로 증가하였으 며, 심장박동수는 1분당 29~32회였다. 배체는 머리 부분부터 난막을 뚫고 나오면서 부화하였다(Fig. 1Q).
3. 자치어의 형태발달
부화 직후의 난황자어는 전장 2.10~2.20mm(평균 2.15±
0.07mm, n=10)로 입과 항문은 열려 있지 않았고, 큰 난황을 가지고 있었으며, 몸 전체에는 막으로 된 지느러미가 형성되 었다. 나뭇가지 모양의 흑색소포는 유구 위에 침착되었고, 눈 의 앞쪽과 등 쪽에도 침착하였다. 눈의 뒤쪽과 복부 및 꼬리자 루에 황색소포가 침착되었으며, 이 시기의 근절 수는 27개였 다(Fig. 2A).
부화 후 1일째 난황자어는 전장 2.44~3.60mm(평균 3.02
±0.82mm)로 항문이 열렸고, 유구와 등 쪽에 침착되었던 흑 색소포와 꼬리자루의 황색소포는 다소 감소하였다. 막지느러 미에는 9~11개의 흑색소포가 침착하였고, 소화관 주위에 황 색소포가 나타났다. 이 시기의 항문은 몸길이의 51.6%로 정중 앙에 가깝게 위치하였다(Fig. 2B).
부화 후 2일째 전기자어는 전장 2.69~3.70mm(평균 3.20
±0.71mm)로 난황이 모두 흡수되었고, 입이 열리기 시작하였
으며, 막지느러미에 나타났던 흑색소포는 소실되었다. 꼬리자 루에는 나뭇가지 모양의 흑색소포가 넓게 침착되었으며, 원기 모양의 가슴지느러미가 형성되었다. 항문은 47.6%로 정중앙 으로부터 앞쪽에 위치하였다(Fig. 2C).
부화 후 4일째 중기자어는 전장 3.16~3.90mm(평균 3.53
±0.52mm)로 꼬리자루에 침착되었던 나뭇가지 모양의 흑색 소포는 소실되었고, 몸통 아래 부분과 소화관 위쪽에 넓게 분
포하였다. 원기모양의 가슴지느러미는 발달하여 2번째 근절까 지 길이가 신장되었다(Fig. 2D).
부화 후 8일째 중기자어는 전장 4.21~4.50mm(평균 4.36
±0.21mm)로 몸통 아래 부분에 막으로 된 지느러미는 항문 을 기준으로 나누어졌으며, 유구는 모두 소실되었다. 아가미 뚜껑 부분에는 반점모양의 흑색소포가 침착되었고, 아래턱의 치골이 발달하면서 먹이 섭취운동이 활발해졌다(Fig. 2E).
Fig. 1. Egg developmental stages of Oplegnathus punctatus. A: Fertilized eggs; B: 2 cells stage, 55mins; C: 4 cells stage, 1hrs 15mins; D: 8 cells stage, 1hrs 45mins; E: 16 cells stage, 2hrs 30mins; F: 32 cells stage, 3hrs; G: 64 cells stage, 3hrs 30mins; H: 128 cells stage, 4hrs 10 mins; I: Morula stage, 4hrs 59mins; J: Blastula stage, 6hrs 22mins; K: Gastrula stage, 8hrs 30mins; L: 4 myotomes stage, 11hrs 40mins; M:
Formation of lens, 13hrs 40mins; N: Melanophore appeared on the embryo, 15hrs 20mins; O: Formation of causal fin, 16hrs 30mins; P: For- mation of otidium, 18hrs; Q: Embryo just before hatching, 20hrs 50mins. Scale bar=1.0mm.
A B C D
E F G H
I J K L
M N O P
Q
부화 후 12일째 중기자어는 전장 4.43~4.80mm(평균 4.62
±0.26mm)로 꼬리지느러미는 부채꼴 모양으로 발달하였고,
흑색소포는 소화관 위쪽으로 침착하였다. 등 쪽과 몸통 아래 부분에는 8~10개의 반점 모양 흑색소포가 침착되었고, 나뭇 가지 모양의 흑색소포는 모두 소실되었다. 아래턱은 위턱보다 길게 발달하였으며, 이 시기의 근절 수는 29~30개로 증가하 였다. 이 시기의 항문 위치는 52.3%로 정중앙으로부터 뒤쪽에 위치하였다(Fig. 2F).
부화 후 15일째 중기자어는 전장 6.68~7.79mm(평균 7.24
±0.78mm)로 척색말단이 45°로 굽어졌고, 등지느러미에 줄기 가 4개 형성되었고, 뒷지느러미에는 5개의 줄기가 형성되었다.
등 쪽과 몸통 아래 부분에 침착되었던 반점 모양의 흑색소포 는 등 쪽이 8개로 줄어들었고, 몸통 아래 부분은 6개로 줄어들 었다(Fig. 3A).
부화 후 18일째 후기자어는 전장 8.71~8.94mm(평균 8.83
±0.16mm)로 막으로 된 배지느러미와 뒷지느러미는 분리되 었고, 꼬리지느러미는 정미 형태로 발달하였으며, 줄기가 뚜 렷하게 형성되기 시작하였다. 각 지느러미별 줄기 수는 등지 느러미가 13개, 뒷지느러미 5개로 증가하였고, 가슴지느러미 7개, 꼬리지느러미는 8개가 형성되었다. 흑색소포는 주상악골 상부와 아가미 뚜껑 및 몸의 정중앙 부분에 반점 모양으로 침 착되어 있었다. 정중앙으로부터 뒤쪽에 위치한 항문은 42.9%
로 정중앙보다 앞쪽에 위치하였다(Fig. 3B).
부화 후 23일째 후기자어는 전장 12.2~12.6mm(평균 12.4
±0.28mm)로 막으로 되었던 지느러미는 모두 분리되었다. 각 지느러미별 줄기 수는 등지느러미가 극조 5개 연조 16개로 증 가하였고, 배지느러미는 극조 1개 연조 4개였으며, 뒷지느러미 Fig. 2. Morphological change of Oplegnathus punctatus from hatching to larva stage. A: Newly hatched yolksac larva, 2.15mm TL; B: yolksac larva, 1 days after hatching(DAH), 3.02mm TL; C: preflexion larva, 2 DAH, 3.20mm TL; D: flexion larva, 4 DAH, 3.53mm TL; E: flexion lar- va, 8 DAH, 4.36mm TL; F: flexion larva, 12 DAH, 4.62mm TL. Scale bars=1.0mm.
A
B
C
D
E
F
Fig. 3. Morphological change of Oplegnathus punctatus from hatch- ing to larva stage. A: postflexion larva, 15 DAH, 7.24mm TL; B:
postflexion larva, 18 DAH, 8.89mm TL; C: postflexion larva, 22 DAH, 12.5mm TL. Scale bars=1.0mm.
A
B
C
는 극조 3개 연조 7개로 증가하였다. 꼬리지느러미 줄기 수는 16개로 증가하였고, 가슴지느러미는 13개로 증가하였다. 꼬 리 기저부분에는 6개의 반점 모양 흑색소포가 침착되었고, 두 부와 몸통 중앙부분까지 반점 모양의 흑색소포가 침착되었다 (Fig. 3C).
부화 후 26일째 후기자어는 전장 13.5~14.1mm(평균 13.8
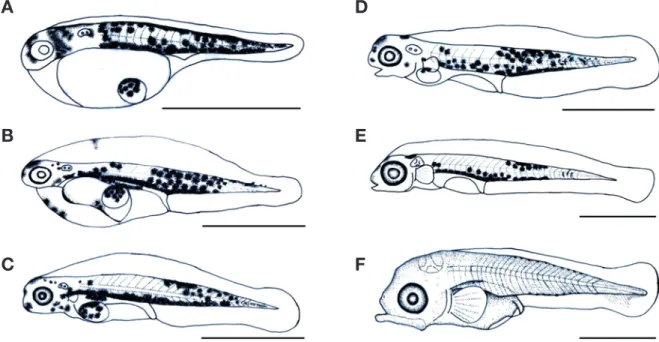
±0.42mm)로 등지느러미의 줄기 수는 연조가 16개로 증가하 였고, 배지느러미가 연조 5개로 증가하였으며, 뒷지느러미 연 조가 10개로 증가하였다. 꼬리 기저부분에 침착된 6개의 반점 모양 흑색소포는 소실되었고, 두부와 몸통 중앙부분에 침착된 흑색소포는 진하게 착색되었으며, 등과 배, 뒷지느러미에는 반 점 모양의 흑색소포가 침착되었다. 이 시기의 항문은 41.9%로 정중앙보다 앞쪽에 위치하였다(Fig. 4A).
부화 후 30일째 후기자어는 전장 16.5~18.0mm(평균 17.3
±1.06mm)로 위턱의 주상악골과 아래턱 치골의 형태가 부리 모양으로 발달하였고, 이빨이 형성되기 시작하였다. 반점 모양 의 흑색소포는 꼬리 기저부분과 등 쪽 및 아가미 뚜껑부분에
진하게 착색되었고, 등과 배, 뒷지느러미에 침착된 반점 모양 의 흑색소포는 진하게 착색되었다. 항문은 52.1%로 정중앙보 다 뒤쪽에 위치하였다(Fig. 4B).
부화 후 35일째 치어는 전장 21.5~24.8mm(평균 23.2±
2.33mm)로 등지느러미의 줄기 수는 극조 10개, 연조 18개로 증가하였고, 뒷지느러미는 연조 12개로 증가하였으며, 가슴지 느러미 줄기 수는 17개로 증가하였다. 흑색소포는 몸통 전체 에 둥근 반점 모양으로 침착되었고, 각 부위별 지느러미에도 반점 모양의 흑색소포가 진하게 착색되었다. 이 시기의 항문 은 60.8%로 정중앙보다 뒤쪽에 위치하였다(Fig. 4C).
고 찰
해산 어류의 생산 기술을 확립하고 자연 상태에서의 자원을 파악하기 위해서는 대상 종들의 초기 발육 과정을 알아야 되 며, 자연에서 채집한 난과 자치어의 종을 동정하는 데 어려움 이 많기 때문에 실내에서 인공수정에 의한 난과 자치어의 발 육 단계로 형태적인 특징을 관찰하고, 다른 종과 비교하는 것 이 중요하다(Byun et al., 2009).
강담돔의 수정란은 무색 투명한 구형의 분리부성란으로 1 개의 유구를 가지고 있으며, 유구에 흑색소포가 침착되는 특 징을 보인다. 분리부성란을 가지는 해산어류는 돌돔(Go and Kim, 1992; Hwang, 2000), 넙치 Paralichthys olivaceus(Han and Kim, 1997), 찰가자미 Microstomus achne(Byun et al., 2009), 능성어 Epinephelus septemfasciatus(Park et al., 2014) 및 붉바리 E. akaara(Lee et al., 1997) 등이 있으며, 모두 1개 의 유구를 가지고 있다. 그러나 강담돔과 돌돔(Go and Kim, 1992; Hwang, 2000)은 유구에 나뭇가지 모양의 흑색소포를 가진다는 점에서 다른 해산어류의 난과 차이를 보였다. 유구 가 소실되는 시기를 비교했을 때 강담돔이 부화 후 8일째에 소실되었고, 돌돔은 부화 후 6일째에 소실되어 다소 차이를 보 였다.
강담돔 난의 크기는 1.01~1.15mm(평균 1.08±0.10mm), 평균 1.06±0.03mm(Kim, 2007)로 유사하였고, 같은 과 어류 인 돌돔(Go and Kim, 1992; Hwang, 2000) 0.82~0.94mm(평 균 0.85mm)보다 큰 편이었다.
수온에 따른 부화 소요시간을 비교해 보면 강담돔은 수온 22.5~24.5°C(평균 23.0±0.05°C)에서 20시간 50분, Kim (2007)에 따르면 수온 22.0°C에서 36시간 30분이 소요되었 다. 돌돔은 수온 20.0~22.3°C에서 35시간(Go and Kim, 1992), 수온 22.5°C에서 36~37시간(Hwang, 2000)이 소요되 었다. 강담돔의 경우 수온차이로 인한 부화 소요시간에 차이 가 있었고, 돌돔(Go and Kim, 1992; Hwang, 2000)의 경우 수 정란 사육수온은 비슷하였으나 강담돔의 부화 소요시간보다 Fig. 4. Morphological change of Oplegnathus punctatus from hatch-
ing to larva and juvenile stage. A: 17.0mm in TL postflexion larvae, 26 days after hatching; B: 19.5mm in TL postflexion larvae, 30 days after hatching; C: 25.3mm in TL juvenile, 35 days after hatching.
A
B
C
길었다.
강담돔의 부화자어 크기는 전장 2.10~2.20mm(평균 2.15
±0.07mm), Kim(2007)에 따르면 평균 전장 2.10mm로 유사 하였고, 돌돔은 평균 전장 1.75mm(Go and Kim, 1992), 평균 전장 2.10mm(Hwang, 2000)로 유사하거나 다소 짧았다.
척색말단이 굽어지는 시기를 비교해 보았을 때 강담돔은 부 화 15일째 평균 전장 7.24mm, 돌돔(Hwang, 2000)은 부화 24 일째 평균 전장 8.90mm일 때 형태적 변화가 관찰되어 강담 돔이 돌돔보다 척색말단이 굽어지는 시기가 빨랐고, 평균 전 장에도 다소 차이가 있었다.
부화자어의 흑색소포는 유구의 위쪽에 나뭇가지 모양으로 침착되었고, 눈의 앞쪽과 등 쪽에도 침착하였으며, 눈의 뒤쪽 과 복부 및 꼬리자루에 황색소포가 침착되었다. 돌돔(Go and Kim, 1992; Hwang, 2000)의 부화자어는 유구의 위쪽에 나뭇 가지 모양의 흑색소포가 침착하였고, 복부에 황색소포가 침착 되었다. 강담돔은 몸통의 위와 아래 부분이 나뭇가지 모양의 흑색소포로 진하게 착색된 점에서 돌돔(Go and Kim, 1992;
Hwang, 2000)과 차이를 보였고, 복부에 침착된 황색소포의 위 치는 두 종이 유사하였다. 이와 같이 흑색소포의 위치, 출현 시기 및 발달 양상은 종에 따라 특수한 형질의 상태를 나타내 는 것이라 생각되며, 이들 흑색소포의 분포 상태는 분류 형질 로서 중요한 역할을 할 것으로 생각된다(Han et al., 2001).
치어기로 이행되는 시기와 크기를 비교해 보았을 때 강담돔 은 부화 후 35일째 평균 전장 23.2mm였고, Kim(2007)에 따 르면 부화 후 30일째 평균 전장 18.2mm로 시기에 따른 크기 차이가 다소 있었다. 돌돔(Go and Kim, 1992; Hwang, 2000) 은 부화 후 37일째 평균 전장 16.7mm로 강담돔보다 짧았다 (Table 1).
일반적으로 어류의 자치어기에는 형태적으로 분류가 어려 우며, 돌돔의 경우는 많은 연구를 통해 자치어 시기의 형태 적 특징에 대한 자료가 확보되어 있다. 그러나 강담돔은 Kim (2007)에 의하여 자치어의 형태에 대해 간략히 설명되었으나 돌돔과 자치어 시기의 형태가 매우 흡사하여 종 간의 차이점
을 밝히기 어렵기 때문에 본 연구에서 사육을 통하여 후기자 어 시기에 나타나는 흑색소포의 분포 위치 및 형태적인 차이 점을 구분할 수 있게 되었다.
요 약
강담돔의 수정난을 부화시켜 사육하면서 난 발생 과정과 자치어 발달과정의 형태변화를 관찰하였다. 수정란은 구형의 무색 투명한 분리 부성란이었고, 1개의 유구를 가지고 있었다.
난의 크기는 1.01~1.15mm 범위였다(평균 1.08±0.10mm, n=20). 사육수온은 22.5~24.5°C(평균 23.0±0.05°C), 염분농 도는 33.5~34.5‰(평균 34.0±0.05‰)의 범위를 유지시켜 주 었다. 난 발생 과정은 수정 25분 후에 난막과 난황이 분리되면 서 배반이 형성되었다. 수정 후 55분에는 난할이 2개로 분리되 어 2세포기에 달하였다. 수정 후 20시간 50분에는 배체가 머 리 부분부터 난막을 뚫고 나오면서 부화하였다. 부화 직후의 난황자어는 전장 2.10~2.20mm(평균 2.15±0.07mm, n=10) 로 입과 항문은 열려 있지 않았고, 난황과 유구는 부화 2일 후 흡수되었다. 부화 후 12일째 중기자어는 전장 4.43~4.80mm (평균 4.62±0.26mm)로 꼬리지느러미는 부채꼴 모양으로 발 달하였고, 흑색소포는 소화관 위쪽으로 침착하였다.
인 용 문 헌
Blaxter, J.H.S. 1974. The early life history of fish. Spring-Verlag Berlin, Heidelberg, 765pp.
Byun, S.G., S.H. Lee, J.H. Hwang, K.H. Han, K.W. Kang, J.D. Kim, Y.C. Kim and B.I. Lee. 2009. Embryonic and Larva Devel- opment of Slime Flounder, Microstomus achne. Dev. Reprod., 13: 281-289.(in Korean)
Go, J.L. and Y.U. Kim. 1992. Embryonic development and mor- phology of larvae and juveniles of parrot Fish, Oplegnathus Table 1. Comparison characters of the eggs and larvae characters in the species Oplegnathidae
Species Fertilized egg
diameter(mm) Time of hatching
(*WT) Larvae size(mm)
at hatching Number of
myomeres Total length of
juveniles(mm) Reference Oplegnathus
punctatus 1.08(1.01~1.15) 20hrs 50mins(23.0°C) 2.15 27 21.5~24.8 Present study
Oplegnathus
punctatus 1.06 36hrs 30mins(22.0°C) 2.10 - 18.2 Kim, 2007
Oplegnathus
fasciatus 0.82~0.94 35hrs(20.0~22.3°C) 1.75 27 16.7 Go and Kim, 1992
Oplegnathus
fasciatus 0.85(0.82~0.94) 36~37hrs(22.5°C) 2.10 - 16.7 Hwang, 2000
*WT: water temperature
fasciatus(Temminck et Schlegel). PhD thesis, Pukyong Nat. Univ., 32: 29-45.(in Korean)
Han, H.K., D.Y. Kang, S.B. Hur and C.Y. Jeon. 2001. Effects of Sa- linity on Spontaneous Maturation and Induced Spawning in Seabass, Lateolabrax japonicus. Kor. J. Aquacult., 14: 213- 220.(in Korean)
Han, K.H. and Y.U. Kim. 1997. The early life history of the floun- der, Paralichthys olivaceus Ⅰ. Development of egg, larvae and juveniles. Bull. Yeosu Nat. Fish. Univ., 11: 105-117.(in Korean)Hwang, H.K. 2000. Parrot Fish, Oplegnathus fas- ciatus of aquaculture. Ministry of Oceans and Fisheries, pp.
13-16.(in Korean)
Hwang, H.K., J.U. Lee, S.G. Yang, S.C. Kim and K.M. Kim. 2005.
Water Temperature and Food on Growth and Survival of Parrot Fish Larvae, Oplegnathus fasciatus. Kor. J. Aqua- cult., 18: 13-18.(in Korean)
Ishiguro, N. and M. Nishida. 1998. A tailless spotted parrot fish Oplegnathus fasciatus lacking the caudal fin. Japan. J. Ich- thyol., 45: 43-45.
Jeong, K.S., S.M. Kim, I.C. Bang, S.Y. Kim and W.K. Lee. 1998.
Induced Spawning of Striped Knife-Jaw, Oplegnathus fas- ciatus by Manipulating Water Temperature and Photoperi- od. Kor. J. Aquacult., 11: 141-149.(in Korean)
Kakizawa, Y., K. Kamishikiryo, M. Shirato, S. Maehara, H. Fuji and S. Iesato. 1980. The tooth development of the Parrot Perch, Oplegnathus fasciatus,(family Oplegnathidae, Tele- ostei). J. Nihon Univ. School of Dentistry, 4: 211-226.
Kawabe, K. and J. Kimura. 2007. Broodstock rearing and egg-taking from spotted parrot fish Oplegnathus punctatus in Chichi- jima, Ogasawara Islands, Southern Japan. Aqua. Sci., 55:
135-136.
Kendall, A.W. Jr., E.H. Ahlstrom and H.G. Moser. 1984. Early life
history stages of fishes and their characters. In: Moser, H.G. et al.(eds.), Ontogeny and Systematics of Fishes. Am.
Soc. Ichthyol. Herpetol., Spec. Publ., 1: 11-22, Allen Press, Lawrence, KS.
Kim, B.H. 1998. Sex differentiation and Gonad development of the Parrot fish, Oplegnathus fasciatus. Jeju Univ., pp. 1-32.(in Korean)
Kim, G.J. 2007. Studies on the production of seed and practical cul- ture of Oplegnathus punctatus. Masan Regio. Mari. Affair.
Port Admin., pp. 44-52.(in Korean)
Kim, I.S., Y. Choi, C.L. Lee, Y.J. Lee, B.J. Kim and J.H. Kim. 2005.
Illustrated book of Korean fishes. Kyo-Hak Publishing, Seoul, 360pp.(in Korean)
Kim, S.Y., I.C. Bang and S.M. Kim. 2000. Sexual Maturation In- ducement of Striped Knife-Jaw, Oplegnathus fasciatus by Manipulating Environmental Condition. Korean J. Ichthy- ol., 12: 46-53.(in Korean)
Kwun, H.J. and J.K. Kim. 2010. Occurrence of Natural Hybrid between Oplegnathus fasciatus and Oplegnathus punctatus from the South Sea of Korea. Korean J. Ichthyol., 22: 201- 205.(in Korean)
Lee, C.K., S.B. Hur, S. Park and B.G. Kim. 1997. Qualities of spawned eggs during the spawning period in red spotted grouper, Epinephelus akaare. Kor. J. Aquacult., 10: 463- 472.(in Korean)
Lee, W.O., K.H. Kim, J.M. Back and M.Y. Song. 2013. Egg devel- opment and early life history of Zacco koreanus(Pisces:
Cyprinidae). Korean J. Ichthyol., 25: 200-207.(in Korean) Park, J.M., J.K. Cho, K.H. Han, N.R. Kim, H.K. Hwang, K.M.
Kim, J.I. Myeong and M.H. Son. 2014. Early life history of the sevenband grouper, Epinephelus septemfasciatus from Korea. Dev. Reprod., 18: 13-23.(in Korean)