HaCaT 인간 각질세포에서 산화적 스트레스에 대한 머루근 수용성 추출물의 세포 보호 효과
최은옥1,2․권다혜1,2․황보현1,2․정진우1,2․남원석3․이연희3 양혜경3․황혜진2,4․김병우2,5․이승훈3․최영현1,2
1동의대학교 한의과대학 생화학교실 및 근・골격계 질환제어 융합연구실
2동의대학교 항노화연구소 및 블루바이오소재개발 및 실용화지원센터
3(주)카나, 4동의대학교 의료・보건・생활대학 식품영양학과
5동의대학교 바이오응용공학부 바이오의약공학전공
Cytoprotective Effect of Water Extract of Vitis amurensis Root against Oxidative Stress in HaCaT Human Keratinocytes
Eun Ok Choi1,2, Da Hye Kwon1,2, Hyun Hwang-Bo1,2, Jin-Woo Jeong1,2, Won Sok Nam3, Youn Hee Lee3, Hye Kyung Yang3, Hye Jin Hwang2,4, Byung Woo Kim2,5,
Seung Hun Lee3, and Yung Hyun Choi1,2
1Open Laboratory for Muscular & Skeletal Disease Control and Department of Biochemistry, Dongeui University College of Korean Medicine
2Anti-Aging Research Center and Blue Bio Industry RIC, 4Department of Food and Nutrition, College of Nursing, Healthcare Sciences & Human Ecology, and
5Biopharmaceutical Engineering Major, Division of Applied Bioengineering, College of Engineering, Dongeui University
3CANA Co., Ltd.
ABSTRACT Root of Vitis amurensis Rupr., a wild-growing grape species, has been used as a traditional medicine for the treatment of various diseases in Asian countries, including Korea. Although several studies on the antioxidant effects of V. amurensis root extracts have been carried out, the underlying molecular mechanisms remain largely unresolved. In the present study, we investigated the cytoprotective efficacies of water extract of V. amurensis root (WEVA) against oxidative stress (hydrogen peroxide, H2O2) in HaCaT human keratinocytes. Our data show that treat- ment with WEVA prior to H2O2 exposure significantly increased HaCaT cell viability, indicating that exposure of HaCaT cells to WEVA conferred a protective effect against oxidative stress. WEVA preconditioning also effectively inhibited H2O2-induced apoptotic cell death by blocking degradation of poly (ADP-ribose) polymerase and down-regulation of pro-caspases-9 and -3. The anti-apoptotic effects of WEVA were associated with attenuation of mitochondrial dysfunc- tion and normalization of expression changes of pro-apoptotic Bax and anti-apoptotic Bcl-2 in H2O2-stimulated HaCaT cells. In addition, WEVA abrogated H2O2-induced intracellular reactive oxygen species (ROS) accumulation and phos- phorylation of histone H2AX at serine 139, a widely used marker of DNA damage. Moreover, WEVA increased the levels of hemeoxygenase-1 and NAD(P)H-quinone oxidoreductase-1, potent antioxidant enzymes associated with in- duction of nuclear transcription factor erythroid-2-like factor 2 (Nrf2). Thus, the present study demonstrates that WEVA protects human keratinocytes against oxidative stress-induced apoptosis and DNA damage, possibly through prevention of ROS generation and activation of Nrf2 signaling. Our data indicate that WEVA may potentially serve as an agent for treatment and prevention of skin disorders caused by oxidative stress.
Key words: Vitis amurensis, antioxidant, apoptosis, ROS, DNA damage
Received 27 September 2017; Accepted 21 November 2017 Corresponding author: Yung Hyun Choi, Open Laboratory for Mus- cular & Skeletal Disease Control and Department of Biochemistry, Dongeui University College of Korean Medicine, Busan 47227, Korea
E-mail: [email protected], Phone: +82-51-850-7413
서 론
햇빛으로부터 받는 자외선(ultra violet)에 과도하게 노출 될 경우 나타나는 피부 손상의 주요 위험 요인이 비정상적인 활성산소종(reactive oxygen species, ROS)의 생성이다 (1,2). 비록 일정 수준의 ROS는 산화・환원 신호에서 생리학
적으로 주요한 세포 내 메신저로 작용하지만, ROS의 통제되 지 않은 과다 생성은 지질, 단백질 및 DNA 등의 손상을 유도 하여 많은 인간 피부질환의 발병 기전에 관여한다(3,4). 결 과적으로 손상된 DNA는 피부세포의 세포사멸(apoptosis) 개시에 기여하며, 피부 상피조직의 파괴를 야기한다(5,6).
따라서 자외선 노출에 대한 보다 안전하고 효과적인 피부 보호를 위한 항산화 물질의 발굴은 산화적 스트레스에 의한 피부질환의 예방과 제어를 위한 중요한 관심사에 해당한다 (7,8).
왕머루(Vitis amurensis Rupr.)는 주로 우리나라와 일본, 중국 등지에 분포하는 Vitaceae family에 속하는 야생 포도 의 일종이다. 특히 뿌리와 줄기는 한국과 일본을 포함한 동 아시아 지역에서 다양한 질환의 예방과 치료 목적으로 오랫 동안 사용되어 왔다(9,10). 최근 연구에서 왕머루의 뿌리(머 루근) 추출물 또는 resveratrol, heyneanol 등의 유효성분 들이 항염증(11), 항종양(12), 노화 방지(13), 신경보호 효 과(14) 등과 같은 다양한 약리적 효능이 있는 것으로 알려져 오고 있다. 특히 β-amyloid로 유도된 산화적 스트레스 차단 효과가 보고됨으로써 알츠하이머 병(Alzheimer’s disease) 에 대한 보호 효과도 가질 수 있는 실험적 근거가 제시되기 도 하였다(15). 아울러 왕머루의 종자 추출물은 산화적 스트 레스에 대한 세포 방어에서 중요한 조절 인자 중 하나인 nu- clear transcription factor erythroid-2-like factor 2 (Nrf2)의 발현 증가와 함께(16), inducible nitric oxide syn- thase와 cyclooxygenase 2를 포함한 염증성 인자들의 발 현을 억제하여 항산화 효능과 함께 강력한 항염증 효능이 있음이 보고된 바 있다(17). 그러나 머루근 추출물에 대한 radical scavenging 효능 및 항산화 작용에 대해서는 상대 적으로 그 효능에 대한 연구가 미비한 실정이다. 본 연구에 서는 피부세포 보호 효과를 가지는 효과적인 기능성 소재의 발굴을 위한 연구의 일환으로 머루근 수용성 추출물(water extract of V. amurensis root, WEVA)의 총 페놀 함량(to- tal phenolic content)을 분석하였고, ferric reducing anti- oxidant power(FRAP) 활성능, 2,2’-diphenyl-1-picryl- hydrazyl(DPPH) 및 2,2’-azino-bis(3-ethylbenzothiazo- line-6-sulfonate) cationic(ABTS・+) 라디칼 소거능을 이 용한 항산화 효능 평가와 산화적 스트레스(hydrogen per- oxide, H2O2)에 노출된 HaCaT 인간 피부 각질세포(kera- tinocyte)에서 apoptosis 보호 효과와 관련 항산화 효능을 평가하였다.
재료 및 방법
세포배양 및 WEVA의 추출과 처리
본 실험에 사용된 HaCaT 인간 피부 각질세포는 제주대 학교 의과대학 생화학교실 현진원 교수에게서 분양받았으 며, 10% fetal bovine serum(FBS), 100 U/mL의 penicillin
및 100 mg/mL의 streptomycin이 함유된 Dulbecco modi- fied Eagle medium(DMEM, Gibco-BRL, Grand Island, NY, USA)을 이용하여 37°C 및 5% CO2의 조건에서 배양하 였다. 머루근은 (주)카나(Busan, Korea)에서 제공받았으며, WEVA를 얻기 위하여 머루근 무게의 10배에 해당하는 증류 수로 100°C에서 3시간 동안 추출하였다. 추출물은 여과지 (Whatman No. 3 filter paper, Whatman International Ltd., Maidstone, UK)로 여과한 후 농축하였으며, 농축물은 동결 건조하여 분말화시켰고 획득 수율은 약 13% 정도였다. 이후 추출물을 멸균 증류수에 100 mg/mL(stock solution)의 농 도로 녹이고, Minisart® Syringe filter(0.2 μm, Sartorius AG, Weender Landstr., Germany)로 거른 후 사용 전까지 는 -20°C에서 보관하였으며, 실험 방법에 따라 적절하게 희석하여 사용하였다.
총 페놀 함량
WEVA에 함유된 총 페놀 함량은 Folin-Ciocalteu 방법 (18)에 따라 측정하였으며, gallic acid(Sigma-Aldrich Co., St. Louis, MO, USA)를 표준물질로 사용하였다. 이를 위하 여 적정 실험 농도로 희석한 WEVA 50 μL에 2%의 Na2CO3
1 mL를 첨가하고 2분간 실온에 방치한 후 50% Folin- Ciocalteu reagent(Sigma-Aldrich Co.) 50 μL를 가하고 실온에서 30분간 반응시킨 다음, enzyme-linked immuno- sorbent assay(ELISA) reader(Molecular Devices, Sun- nyvale, CA, USA)를 사용하여 750 nm에서 흡광도를 측정 하였다. Gallic acid를 이용하여 검량선을 작성하였으며, 총 페놀 함량은 mg gallic acid equivalents(GAE)/g of sam- ple dry weight(DW)로 나타내었다.
FRAP 활성 측정
이 방법은 산성 pH 영역에서 ferric tripyridyltriazine (Fe3+-TPTZ) 복합체가 환원성 물질에 의해 청색의 fer- rous tripyridyltriazine(Fe2+-TPTZ)으로 환원되어 593 nm에서 흡광도가 증가하는 원리(19)를 이용한 것으로 대부 분의 항산화제가 환원력을 가지고 있다는 점에 착안하여 고 안된 실험방법이다. FRAP에 의한 WEVA의 항산화력을 측 정하기 위하여 300 mM acetate buffer(pH 3.6), 40 mM HCl에 용해한 10 mM 2,4,6-tripyridyl-s-triazine(TPTZ, Sigma-Aldrich Co.) 및 20 mM FeCl3・6H2O를 각각 10:1:
1(v/v/v)의 비율로 혼합하여 FRAP 시약을 제조하였다. 이 어서 WEVA를 적정 농도로 희석된 시료액 10 μL와 240 μL의 FRAP 시약을 혼합하고 37°C에서 5분간 반응시킨 후 593 nm에서 흡광도를 측정하였다. 양성 대조군(positive control)은 Trolox 및 2,3-butyl-4-hydroxyanisole(BHA, Sigma-Aldrich Co.)을 사용하여 검량선을 작성하였으며, 결과는 mg Trolox/g of sample dry weight(DW)로 나타내 었다.
DPPH 라디칼 소거능
DPPH 라디칼에 전자를 공여함으로써 라디칼을 소거하는 효과를 측정하는 방법인 DPPH 라디칼 소거능은 Guo 등 (20)의 방법에 따라 조사하였다. 이를 위하여 0.4 mM의 DPPH(Sigma-Aldrich Co.) 용액 150 μL에 WEVA가 적정 농도로 희석된 시료 100 μL를 첨가하고 30분간 37°C에서 반응시킨 후 ELISA reader로 518 nm에서 흡광도를 측정하 였다. 그리고 반응물에 대한 흡광도 값을 대조군에 대한 WEVA의 DPPH 라디칼 소거 활성으로 항산화 활성도를 나 타내었으며, Trolox를 양성 대조군으로 비교하였고, 음성 대조군으로는 0.4 mM DPPH 용액 대신 메탄올을 이용한 아래 식에 따라 산출하였다.
DPPH radical scavenging activity (%)=
(
1- O.D. of sample)
×100O.D. of control
ABTS・+ radical cation 소거능
Potassium persulfate와의 반응에 의해 생성된 ABTS 유 리 라디칼이 제거되어 라디칼 특유의 색인 청록색으로 탈색 되는 원리에 바탕을 둔 ABTS radical cation 소거능을 이용 한 WEVA의 항산화능 측정은 Re 등(21)의 방법에 따라 조 사하였다. 즉 7 mM의 ABTS(Sigma-Aldrich Co.)와 2.45 mM potassium persulfate를 1:1로 혼합하여 실온, 암하에 서 24시간 동안 반응시켜 라디칼을 형성한 다음 실험 직전 에 ABTS 용액을 734 nm에서 흡광도가 0.7±0.02가 되도 록 phosphate buffer saline(PBS, pH 7.4)으로 희석하였 다. ABTS radical cation solution 190 μL에 실험 농도로 희석한 시료 10 μL를 가하여 실온에서 6분간 반응시켜 734 nm에서 흡광도를 측정하였으며 Trolox를 양성 대조군으로 하여 아래 식에 따라 항산화 효능을 비교하였다.
ABTS radical scavenging activity (%)=
(
1- O.D. of sample)
×100O.D. of control
세포 생존율 측정
산화적 스트레스에 의한 세포독성에 미치는 WEVA의 영 향을 조사하기 위하여 HaCaT 세포에 적정 농도의 H2O2 (Sigma-Aldrich Co.)를 24시간 처리하거나 WEVA를 1시 간 전처리 후 H2O2를 24시간 처리하였다. 처리가 끝난 세포 에 3시간 동안 0.5 mg/mL의 3-(4,5-dimethylthiazol-2- yl)-2,5-diphenyltetrazolium bromide(MTT, Sigma-Al- drich Co.)를 처리하여 형성된 formazan을 dimethyl sulf- oxide(DMSO)에 용해시킨 후 ELISA reader를 이용하여 540 nm에서 흡광도 변화를 측정하여 대조군에 대한 세포 생존율을 백분율로 표시하였으며, 대조군으로는 N-acetyl cysteine(NAC, Sigma-Aldrich Co.)을 사용하였다.
Apoptosis 유발 여부의 형태적 관찰
산화적 스트레스에 의해 apoptosis가 유발된 세포에서 특이적으로 나타나는 핵의 형태적 변화를 관찰하기 위하여 다양한 조건에서 배양된 HaCaT 세포를 PBS로 2회 수세 후 0.1 M phosphate buffer(pH 7.2)에 희석된 3.7% para- formaldehyde 고정액(Sigma-Aldrich Co.)을 1시간 처리 하였다. 고정된 세포를 수세한 후 0.05 M phosphate buffer (pH 7.2)에 2.5 μg/mL의 농도로 희석된 4,6-diamidino-2- phenylindole(DAPI, Sigma-Aldrich Co.) 용액으로 상온 암 하에서 20분 염색하였다. 이를 다시 PBS로 수세하여 건조시 킨 후 형광현미경(Fluorescence Microscopy, Carl Zeiss, Oberkochen, Germany) 하에서 핵의 형태를 비교하였다.
DNA flow cytometry에 의한 apoptosis 유도의 정량적 분석 산화적 스트레스에 의한 HaCaT 세포의 apoptosis 유발 에 미치는 WEVA의 영향을 정량적으로 분석하기 위하여 Cycle TEST PLUS DNA REAGENT Kit(Becton Dickin- son, San Jose, CA, USA)을 사용하였다. 준비된 세포를 protocol에 따라 고정 후 4°C, 암실에서 30분 동안 propi- dium iodide(PI) 용액에 반응시켰다. 이들 세포를 35-mm mesh를 이용하여 단일세포로 분리하고 실험군당 최소 10,000개 이상의 세포를 flow cytometer(Becton Dickin- son)에 적용시켜 세포 내 DNA 함량에 따른 histogram을 대상으로 sub-G1기에 속하는 세포를 apoptosis가 유발된 세포로 산출하였다.
Mitochondrial membrane potential(MMP, Δψm) 변화의 분석
산화적 스트레스에 의한 미토콘드리아 기능 손상의 여부 를 확인하기 위하여 MMP 값 변화 정도를 측정하였다. 이를 위하여 다양한 조건에서 배양된 HaCaT 세포에 10 μM의 5,5’,6,6’-tetrachloro-1,1’,3,3’-tetraethyl-imidacar- bocyanine iodide(JC-1, Sigma-Aldrich Co.) 용액을 처리 하여 암하, 상온에서 20분 동안 반응시켰다. 반응이 끝난 후 상층액을 제거하고 PBS를 첨가하여 세포를 부유시킨 다 음 flow cytometer에 적용시켜 MMP의 변화를 측정하였다.
ROS 생성 변화 측정
산화적 스트레스에 의한 HaCaT 세포 내 ROS의 생성과 WEVA에 의한 차단 효과의 정도를 확인하기 위하여 준비된 세포들을 fluorescent probe인 10 μM의 2’,7’-di-chloro- dihydrofluorescein diacetate(DCF-DA, Molecular Probes, Leiden, Netherlands)로 20분간 염색 후 형광현미경(Fluo- rescence Microscopy, Carl Zeiss) 하에서 ROS의 생성 변 화 여부를 조사하였다. 아울러 동일한 조건에서 준비된 세포 들에서 ROS 생성에 대한 정량적인 비교를 위하여 반응이 끝난 세포들을 flow cytometer를 적용시켜 ROS 값의 변화 를 분석하였다.
Table 1. Total phenolics contents and ferric reducing antioxidant power of water extract of Vitis amurensis Rupr. root (WEVA)
Total phenolic content (mg GAE/g)
FRAP (mg Trolox/g) WEVA 56.37±6.57 WEVA
BHA
49.03±5.61 878.61±128.03 Values are presented as the mean±SD.
A
B
Fig. 1. Effects of WEVA on DPPH and ABTS radical scaveng- ing activities. Trolox (T, 0.1 μg/mL) and BHA (B, 0.1 μg/mL) were used as the positive controls. Each point represents the mean±SD of three independent experiments.
단백질의 분리 및 western blot analysis
WEVA의 산화적 스트레스 보호에 따른 특정 유전자들의 발현 변화 관찰을 위하여 준비된 세포에 적당량의 lysis buf- fer[25 mM Tris-Cl(pH 7.5), 250 mM NaCl, 5 mM ethyl- enediaminetetraacetic acid, 1% Nonidet-P40, 1 mM phenylmethylsulfonyl fluoride, 5 mM dithiothreitol]를 첨가하여 4°C에서 1시간 이상 반응시켰다. 상층액에 있는 단백질을 분리한 후 동량의 단백질을 sodium dodecyl sul- fate(SDS)-polyacrylamide gel을 이용하여 전기영동을 실 시하였다. Gel에 함유된 단백질을 polyvinylidene difluor- ide membrane(Schleicher & Schuell Bioscience Inc., Keene, NH, USA)으로 전이시키고 membrane을 5% skim milk 용액에 1시간 처리하여 비특이적인 단백질들에 대한 blocking을 실시하였다. 그리고 Santa Cruz Biotechnol- ogy Inc.(Dallas, TX, USA) 및 Cell Signaling Technol- ogy Inc.(Danvers, MA, USA)에서 구입한 적정 1차 항체를 처리하여 상온에서 2시간 이상 반응시킨 후, PBS-T 용액 (PBS with Tween 20)으로 세척하고 2차 항체를 상온에서 1시간 정도 반응시켰다. 반응이 끝난 후 암실에서 enhanced chemiluminescence(ECL) solution(Santa Cruz Biotech- nology Inc.)을 적용시킨 다음 X-ray film에 감광시켜 특정 단백질의 발현 변화를 분석하였다.
통계 분석
각각의 평가 분석은 3회 이상 반복실험 하였고 실험 결과 들의 유의성을 검정하기 위하여 분산분석(ANOVA)을 실시 한 후 P<0.05 수준에서 Duncan’s multiple range tests를 실시하였으며, 그 결과는 평균(mean)±표준편차(standard deviation, SD)로 표시하였다.
결과 및 고찰
WEVA의 총 페놀 함량과 FRAP 활성능, DPPH 및 ABTS・+ 라디칼 소거능에 측정
일반적으로 식물유래 생리활성 성분의 많은 부분은 2차 대사산물로서 phenolic hydroxyl기를 가지는 방향족 화합 물의 총칭인 페놀 화합물(phenolic compounds)이다(22, 23). 따라서 특정 식물 추출물의 항산화능을 비교하기 위한 비교 지표로서 페놀 화합물의 양적 평가는 가장 기본적으로 수행되는 과정이다. 본 연구에서 사용된 WEVA의 표준화를 위한 기본적인 방법으로 준비된 WEVA에 함유된 총 페놀 함량을 조사하였으며, gallic acid를 표준용액으로 선정하여 작성한 검정곡선으로부터 얻은 값은 56.37 mg GAE/g으로 산출되었다(Table 1). 아울러 FRAP법에 의한 WEVA의 환 원력은 49.03 mg Trolox/g으로 나타났으며, DPPH 및 ABTS 라디칼 소거능을 조사한 결과는 Fig. 1에 나타낸 바 와 같다. DPPH 라디칼 소거능의 IC50은 머루근이 2.18 mg/mL, BHA는 16.39 μg/mL, Trolox는 9.98 μg/mL이며,
ABTS 라디칼 소거능의 IC50은 머루근이 0.29 mg/mL, BHA 는 11.85 μg/mL, Trolox는 12.37 μg/mL로 나타났다. Chung (24)의 연구 결과와 비교해보면 머루와 유사한 외관을 가진 아사이베리의 수용성 추출물의 DPPH 라디칼 소거능은 0.5 mg/mL에서 약 20%, 1 mg/mL에서 약 40% 정도로 나타나 본 연구에서 사용된 머루근과 유사한 DPPH 라디칼 소거 활성이 나타난 것을 확인할 수 있었다. 이러한 결과는 향후 실시할 유기용매별 머루근의 항산화력 비교와 WEVA의 항 산화력을 다른 페놀 화합물과의 비교를 위한 지표로 사용될 것이다.
HaCaT 각질세포에서 H2O2로 유도된 산화적 스트레스에 대 한 WEVA의 보호 효과
HaCaT 세포를 이용한 in vitro 항산화력의 평가를 위한 실험 조건의 설정을 위하여 H2O2 및 WEVA가 HaCaT 세포 의 생존에 미치는 영향을 조사하였다. 이를 위하여 HaCaT 세포를 다양한 농도의 H2O2(10~500 μM) 및 WEVA(0.25~
2.0 mg/mL)가 처리된 조건에서 24시간 배양 후 MTT as- say를 실시한 결과, H2O2 처리 농도의 증가에 따라 HaCaT
A
B
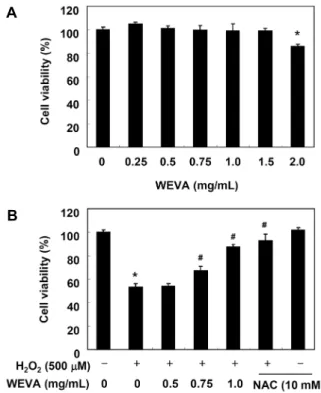
Fig. 2. Effect of WEVA on cell viability in H2O2-treated HaCaT human keratinocytes. HaCaT cells were incubated with the in- dicated concentrations of WEVA for 24 h (A) or WEVA (0.5∼
1.0 mg/mL) and NAC (10 mM) in the presence or absence of 500 μM H2O2 for 24 h (B). The cell viability was assessed by the MTT reduction assay, and the results are expressed as the percentage of surviving cells over the control cells (no addition of WEVA and H2O2). Values represent the mean±SD of three independent experiments. *P<0.05 compared to the untreated control. #P<0.05 compared to the H2O2 treated group.
세포의 생존율이 유의적으로 감소하여 500 μM 처리군에서 약 50% 정도의 생존력으로 보였다(data not shown). WEVA 의 경우 1.5 mg/mL 이하의 농도에서 유의적인 생존율 변화 를 보이지는 않았지만, 2.0 mg/mL 처리군에서 대조군 대비 약 85% 정도의 생존율을 나타내었다(Fig. 2A). 따라서 500 μM의 H2O2를 산화적 스트레스 조건으로 설정하였으며, 이 에 따른 WEVA의 세포독성 보호 효과를 조사하였다. Fig.
2B에 나타낸 바와 같이 H2O2를 처리하기 1시간 전에 WEVA 를 노출시켰을 경우 H2O2 단독 처리군보다 WEVA 처리군에 서 농도 의존적으로 세포 생존율이 증가(0.75 mg/mL 및 1.0 mg/mL의 WEVA 처리군에서 각각 약 67% 및 87%)하 여 WEVA는 H2O2 처리에 의한 세포 생존율 저하를 향상시 킬 수 있었음을 알 수 있었다. 대조군으로 사용된 10 mM의 NAC 전처리군에서는 WEVA 처리 농도의 1.0 mg/mL와 유 사한 효능을 나타내었다. 따라서 모든 후속 실험은 1.0 mg/
mL의 WEVA로 1시간 전처리한 후 500 μM의 H2O2에 24시 간 노출시킨 HaCaT 세포를 대상으로 수행했다.
산화적 스트레스에 의한 HaCaT 각질세포의 apoptosis 유발에 미치는 WEVA의 영향
이상에서 관찰된 산화적 스트레스에 대한 HaCaT 세포의 WEVA 보호 효과가 apoptosis 유도 억제와 연관성이 있는 지를 조사하였다. 이를 위하여 apoptosis가 일어난 세포의 핵에서 특이적으로 관찰되는 염색질의 응축(chromatin con- densation)에 따른 apoptotic body의 형성에 미치는 영향 을 DAPI 염색을 통하여 조사하였다. Fig. 3A에 나타낸 바와 같이 H2O2만이 처리된 HaCaT 세포에서는 apoptosis가 유 발되었음을 의미하는 전형적인 핵의 형태 변화가 관찰되었 으나, WEVA 전처리군에서는 이러한 현상을 거의 관찰할 수 없었다.
한편 산화적 스트레스에 의한 세포의 사멸은 caspase cascade의 활성과 연관되어 있으며(25,26), caspase 활성 의존적인 apoptosis 유발에서는 effector caspase의 활성 에 따른 poly(ADP-ribose) polymerase(PARP)와 같은 기 질 단백질의 분해가 동반된다(26,27). 따라서 WEVA의 산 화적 스트레스 억제 효능이 caspase 활성 의존적 apopto- sis 유발의 억제에 의한 것인지를 확인하기 위하여 PARP의 발현 변화를 관찰한 결과, H2O2만이 단독 처리된 조건에서 배양된 HaCaT 세포에서는 전형적인 PARP 단편화 현상이 관찰되었으며, 이러한 현상은 WEVA가 전처리된 조건에서 완벽하게 억제되었다(Fig. 3B). 아울러 PARP의 단편화와 연관된 대표적인 effector caspase인 caspase-3과 cas- pase-3의 상위 initiator caspase인 caspase-9(28,29)의 불활성형 발현이 H2O2 단독 처리군에서는 뚜렷한 감소 경향 을 보였으나, WEVA의 전처리군에서는 대조군 수준으로 유 지되었다(Fig. 3B). 이러한 WEVA의 산화적 스트레스에 의 한 HaCaT 세포의 apoptosis 유도 억제 여부를 정량적으로 분석하기 위하여 flow cytometry 분석을 수행한 결과, H2O2가 단독 처리된 세포에서는 apoptosis 유발군에 해당 하는 sub-G1기에 속하는 세포의 빈도가 약 28.7% 정도임 에 비하여 WEVA의 전처리군에서는 약 6.3% 정도로 나타 났다(Fig. 3C). 따라서 산화적 스트레스에 의하여 유도되는 HaCaT 세포의 생존율 저하는 apoptosis 유도와 연관성이 있으면 WEVA는 apoptosis 유도에 핵심이 되는 caspase 활성 경로를 차단함으로써 이를 효과적으로 억제하였음을 알 수 있었다.
산화적 스트레스에 의한 HaCaT 각질세포의 미토콘드리아 기능 손상에 미치는 WEVA의 영향
선행 연구들에 의하면 산화적 스트레스에 따른 apopto- sis 유도는 미토콘드리아의 기능적 손상에 따른 intrinsic apoptosis 경로의 활성과 직접적인 관련이 있는 것으로 알 려져 있다(3,24). 이러한 미토콘드리아 기능 손상과 연계된 apoptosis 유도 조절에 가장 중요하게 작용하는 유전자군이 Bcl-2 family이며, Bcl-2 family에 속하는 단백질은 apop- tosis를 유도하거나 억제하는 인자들로 구성되어 있다(29, 30). 현재 가장 대표적인 미토콘드리아의 기능 측정 방법은 MMP 값의 변화 여부를 조사하는 것이며, MMP 값의 저하
A B
C
Fig. 3. Protection against H2O2-induced apopto- sis by WEVA in HaCaT human keratinocytes.
The cells were pre-treated with 1.0 mg/mL WEVA for 1 h and then incubated without or with 500 μM H2O2 for 24 h. (A) The cells were fixed and stained with DAPI solution. The stained nu- clei were observed under a fluorescence micro- scope (×400 original magnification). (B) Cellu- lar proteins were separated on SDS-polyacryla- mide gels and were subsequently transferred on- to membranes. The membranes were probed with an anti-bodies against PARP, caspase-9, and caspase-3. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (C) To quantify the degree of apoptosis, the cell cycle analysis was performed using flow cytometry, and the cell percentages of the sub- G1 peaks in the cell cycle were analyzed. Data are presented as mean of two independent ex- periments.
A
B
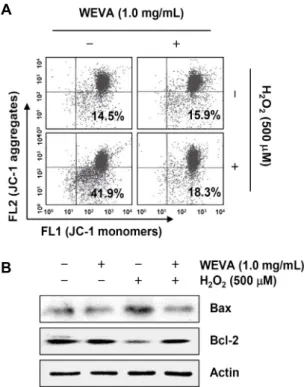
Fig. 4. Effects of WEVA on the MMP values and the expression of Bax and Bcl-2 in H2O2-treated HaCaT human keratinocytes.
The cells were pre-treated with 1.0 mg/mL WEVA for 1 h and then stimulated with and without 500 μM H2O2 for 24 h. (A) The cells were collected and incubated with 10 μM JC-1 for 20 min at 37°C in the dark. The cells were washed with PBS and analyzed using a flow cytometer. Data are presented as mean of two independent experiments. (B) The cells were lysed, and the aliquots containing the total proteins were subjected to SDS-polyacrylamide gels followed by immunoblot analysis with anti-Bax and anti-Bcl-2 antibodies. Actin was used as an internal control.
는 미토콘드리아의 기능이 손상되었음을 나타내는 지표이다 (31). 따라서 산화적 스트레스에 의한 HaCaT 세포의 apop- tosis 유도에 미토콘드리아 기능 손상이 연계되어 있는지, 그리고 WEVA가 이를 차단할 수 있는지를 조사하였다. Fig.
4A에 나타낸 바와 같이 대조군과 WEVA 단독 처리군에 비 하여 H2O2가 함유된 배지에서 배양된 HaCaT 세포에서의 MMP의 소실 정도는 41.9%로 나타나 대조군보다 약 2.9배 증가하였지만, WEVA 전처리 조건군에서는 18.3% 정도를 보였다. 아울러 H2O2의 처리에 의하여 증가하였던 미토콘드 리아 기능 손상을 유발하는 pro-apoptotic Bax의 발현이 증가한 반면, anti-apoptotic Bcl-2의 발현은 감소하였으나 WEVA의 전처리에 의해서는 두 Bcl-2 family 단백질들의 발현이 대조군 수준으로 유지되었다(Fig. 4B). 이러한 결과 들은 WEVA가 H2O2에 의한 Bcl-2 family 인자들의 발현 변화를 억제함으로써 미토콘드리아 기능 손상을 차단시켜 apoptosis 유도를 억제하였음을 보여주는 것이다.
산화적 스트레스에 의한 HaCaT 각질세포의 DNA 손상에 미치는 WEVA의 영향
다음은 이상에서 관찰된 WEVA의 산화적 스트레스에 따 른 apoptosis 방어 효과가 ROS의 생성 및 DNA 손상 억제와 직접적인 연관성이 있는지를 조사하였다. 이러한 조사를 실 시한 이유는 세포 내에서 생산되는 ROS 대부분은 미토콘드 리아 기능 손상과 연관되며, 이로 인한 apoptosis 유발은 DNA 손상과 연계되기 때문이다(3,24). 이를 위하여 H2O2가 처리된 HaCaT 세포를 대상으로 DCF-DA 염색을 통하여 ROS의 생성 여부를 조사한 결과, H2O2 처리 30분 이내에 ROS의 생성이 증가하기 시작하여 6시간 경과 후 최고치를
A B
C
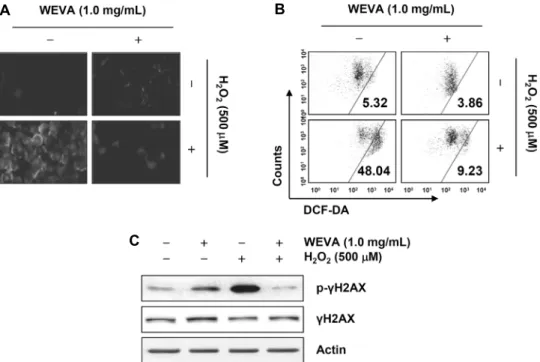
Fig. 5. Protection of H2O2-induced ROS generation and DNA damage by WEVA in HaCaT human keratinocytes. The cells were pre-treated with 1.0 mg/mL WEVA for 1 h and then stimulated with and without 500 μM H2O2 for 6 h (A and B) or 24 h (C).
(A) In order to monitor ROS production, the cells were incubated at 37°C in the dark for 20 min with culture medium containing 10 μM DCF-DA. Images were obtained using a fluorescence microscope, and those presented are from one experiment, and are representative of at least three independent experiments. (B) ROS generation was measured by flow cytometry. Each point represents the mean of two independent experiments. (C) The cells were lysed and then equal amounts of cell lysates were separated on SDS- polyacrylamide gels and transferred to membranes. The membranes were probed with specific antibodies against p-γH2AX, γH2AX, and actin as an internal control, and the proteins were visualized using an ECL detection system.
Fig. 6. Induction of Nrf2, HO-1, and NQO-1 expression by WEVA in HaCaT human keratinocytes. HaCaT cells were in- cubated with various concentrations of WEVA for 24 h. Total cellular proteins were separated on SDS-polyacrylamide gels and then transferred onto membranes. The membranes were pro- bed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
나타낸 후 다시 감소하였다(data not shown). 그러나 WEVA 를 1시간 전처리 후 H2O2를 6시간 처리하였을 경우 대조군 수준까지는 아니지만 현저하게 감소하여 WEVA가 강력한 항산화 효능을 가지고 있음을 형광현미경적 관찰과 flow cytometry 분석을 통하여 확인할 수 있었다(Fig. 5A, 5B).
따라서 WEVA의 항산화 효능이 산화적 스트레스에 의한 DNA 손상 차단과의 연관성이 있는지를 확인하기 위하여 DNA 이중 나선 손상 지표인 γH2AX 단백질의 인산화(serine 139)(32)에 미치는 WEVA의 영향을 조사하였다. Fig. 5C의 immunoblotting 결과에 의하면 H2O2에 노출된 HaCaT 세 포에서 γH2AX 단백질의 전체적인 발현에는 큰 변화 없이 인산화가 매우 증가하였으나, WEVA 전처리된 세포에서는 이러한 현상을 관찰할 수 없었다. 따라서 WEVA에 의한 DNA 손상의 방어 효과는 ROS 생성의 차단과 직접적인 연 관성이 있음을 알 수 있었다.
HaCaT 각질세포의 Nrf2 및 Nrf2 관련 유전자 발현에 미치 는 WEVA의 영향
Nrf2는 산화-환원 감수성 전사인자(redox-sensitive transcription factor)로서 다양한 항산화 효소 발현을 조절 하는 것으로 알려져 있다(33,34). 산화적 방어 신호가 존재 하지 않을 경우 Nrf2는 보통 세포질에서 Nrf2의 억제제인 Kelch-like ECH-associated protein 1(Keap1)과 복합체
를 이루고 있지만, Nrf2의 활성 신호에 의하여 Keap1과 분 리되면 핵 속으로 이동하여 해당 유전자 promoter의 항산 화 반응인자(antioxidant response element, ARE) 자리에 결합하여 heme oxygenase-1(HO-1) 및 NAD(P)H-qui- none oxidoreductase-1(NQO-1)을 포함한 다양한 항산화 효소들의 전사 활성을 증가시킨다(33,34). 따라서 이 신호 계의 활성 유도를 위한 신규 물질의 발굴은 산화적 피부 손
상뿐만 아니라 산화적 스트레스와 연관된 다양한 퇴행성 및 대사성 질환의 예방과 치료에 유용하게 적용될 수 있다(35, 36). 본 연구에서 관찰된 WEVA의 산화적 스트레스에 대한 방어 작용이 이러한 Nrf2 신호 전달계의 활성과 연관성이 있는지를 조사하기 위하여 WEVA가 첨가된 배지에서 배양 된 HaCaT 세포를 대상으로 Nrf2, HO-1 및 NQO-1의 발현 증가 여부를 조사하였다. Fig. 6에 제시된 western blot analysis의 결과에서 알 수 있듯이 WEVA 처리에 따라 Keap1의 발현 감소와 함께 Nrf2의 발현이 증가하였으며, 그에 따른 HO-1 및 NQO-1의 발현도 증가하였다. 따라서 HaCaT 세포에서 WEVA의 항산화 활성에는 최소한 Nrf2 경로가 관여하였음을 알 수 있었다.
요 약
왕머루(Vitis amurensis Rupr.)의 뿌리(머루근)는 한국을 비롯한 아시아 각국에서 다양한 질환의 예방과 치료의 목적 으로 오랫동안 사용되어 왔다. 비록 머루근 추출물의 항산화 효과에 대한 몇몇 보고가 있었지만, 근본적인 분자생물학적 기전 연구는 아직 완전히 이해되지 못하고 있다. 본 연구에 서는 HaCaT 각질세포에서 산화적 스트레스(H2O2)에 대한 머루근 열수 추출물(WEVA)의 세포 보호 효과를 조사하였 다. 본 연구의 결과에 의하면 WEVA는 H2O2에 의한 HaCaT 각질세포의 생존력 저하를 현저하게 억제시켜 WEVA가 산화 적 스트레스에 대한 보호 효과가 있음을 보여주었다. WEVA 는 또한 H2O2에 의한 PARP 분해와 pro-caspase-9와 -3 의 발현 억제를 차단함으로써 H2O2에 의한 apoptosis를 효 과적으로 억제시켰다. 이러한 WEVA의 apoptosis 억제 효 과는 H2O2에 자극받은 HaCaT 세포에서 미토콘드리아 기능 손상의 감소 및 pro-apoptotic Bax와 anti-apoptotic Bcl- 2의 발현 변화의 차단과 관련이 있었다. 또한, WEVA는 H2O2에 의해 유도된 세포 내 ROS 축적과 DNA 손상을 억제 하였고, Nrf2의 발현 증가와 관련된 강력한 항산화 효소인 HO-1과 NQO-1의 발현을 유도하였다. 이러한 결과는 WEVA가 ROS의 생성 억제 및 Nrf2 신호 전달계 활성을 통하여 각질세포를 산화적 스트레스에 의한 apoptosis 및 DNA 손상으로부터 보호하였음을 의미한다. 따라서 본 연구 의 결과는 WEVA가 산화적 스트레스로 인한 피부질환의 치 료 및 예방을 위한 기능성 소재로 유용성이 있음을 보여주었 다.
감사의 글
본 결과물은 산업통상자원부・부산광역시 지원 지역혁신센 터사업 동의대학교 블루바이오 소재개발 및 실용화 지원센 터(RIC08-06-07) 및 한국연구재단의 지원에 의해 수행되 었음(2015R1A2A2A01004633).
REFERENCES
1. Kammeyer A, Luiten RM. 2015. Oxidation events and skin aging. Ageing Res Rev 21: 16-29.
2. Karran P, Brem R. 2016. Protein oxidation, UVA and human DNA repair. DNA Repair (Amst) 44: 178-185.
3. Birch-Machin MA, Russell EV, Latimer JA. 2013. Mito- chondrial DNA damage as a biomarker for ultraviolet radia- tion exposure and oxidative stress. Br J Dermatol 169: 9-14.
4. Katiyar SK. 2016. Dietary proanthocyanidins inhibit UV ra- diation-induced skin tumor development through functional activation of the immune system. Mol Nutr Food Res 60:
1374-1382.
5. Brunelle JK, Chandel NS. 2002. Oxygen deprivation induced cell death: An update. Apoptosis 7: 475-482.
6. Cadet J, Douki T, Ravanat JL. 2015. Oxidatively generated damage to cellular DNA by UVB and UVA radiation. Pho- tochem Photobiol 91: 140-155.
7. Oresajo C, Pillai S, Manco M, Yatskayer M, McDaniel D.
2012. Antioxidants and the skin: understanding formulation and efficacy. Dermatol Ther. 25: 252-259.
8. Tundis R, Loizzo MR, Bonesi M, Menichini F. 2015. Poten- tial role of natural compounds against skin aging. Curr Med Chem 22: 1515-1538.
9. Huang KS, Lin M. 1999. Oligostilbenes from the roots of Vitis amurensis. J Asian Nat Prod Res 2: 21-28.
10. Nassiri-Asl M, Hosseinzadeh H. 2009. Review of the phar- macological effects of Vitis vinifera (Grape) and its bioac- tive compounds. Phytother Res 23: 1197-1204.
11. Huang KS, Lin M, Cheng GF. 2001. Anti-inflammatory tet- ramers of resveratrol from the roots of Vitis amurensis and the conformations of the seven-membered ring in some oligostilbenes. Phytochemistry 58: 357-362.
12. Lee EO, Lee HJ, Hwang HS, Ahn KS, Chae C, Kang KS, Lu J, Kim SH. 2006. Potent inhibition of Lewis lung cancer growth by heyneanol A from the roots of Vitis amurensis through apoptotic and anti-angiogenic activities. Carcino- genesis 27: 2059-2069.
13. de la Lastra CA, Villegas I. 2005. Resveratrol as an anti-in- flammatory and anti-aging agent: mechanisms and clinical implications. Mol Nutr Food Res 49: 405-430.
14. Huang G, Cao X, Zhang X, Chang H, Yang Y, Du W, Wilson JX. 2009. Effects of soybean isoflavone on the notch signal pathway of the brain in rats with cerebral ischemia. J Nutr Sci Vitaminol (Tokyo) 55: 326-331.
15. Jang MH, Piao XL, Kim HY, Cho EJ, Baek SH, Kwon SW, Park JH. 2007. Resveratrol oligomers from Vitis amurensis attenuate β-amyloid-induced oxidative stress in PC12 cells.
Biol Pharm Bull 30: 1130-1134.
16. Bak MJ, Jun M, Jeong WS. 2012. Procyanidins from wild grape (Vitis amurensis) seeds regulate ARE-mediated en- zyme expression via Nrf2 coupled with p38 and PI3K/Akt pathway in HepG2 cells. Int J Mol Sci 13: 801-818.
17. Bak MJ, Truong VL, Kang HS, Jun M, Jeong WS. 2013.
Anti-inflammatory effect of procyanidins from wild grape (Vitis amurensis) seeds in LPS-induced RAW 264.7 cells.
Oxid Med Cell Longev 2013: 409321.
18. Slupphaug G, Kavli B, Krokan HE. 2003. The interacting pathways for prevention and repair of oxidative DNA dam- age. Mutat Res 531: 231-251.
19. Benzie IF, Strain JJ. 1996. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal Biochem 239: 70-76.
20. Guo XY, Wang J, Wang NL, Kitanaka S, Yao XS. 2007.
9,10-Dihydrophenanthrene derivatives from Pholidota yun- nanensis and scavenging activity on DPPH free radical. J Asian Nat Prod Res 9: 165-174.
21. Re R, Pellegrini N, Proteggente A, Pannala A, Yang M, Rice-Evans C. 1999. Antioxidant activity applying an im- proved ABTS radical cation decolorization assay. Free Radic Biol Med 26: 1231-1237.
22. Zhu YZ, Huang SH, Tan BK, Sun J, Whiteman M, Zhu YC. 2004. Antioxidants in Chinese herbal medicines: a bio- chemical perspective. Nat Prod Rep 21: 478-489.
23. Gülçin İ. 2012. Antioxidant activity of food constituents: an overview. Arch Toxicol 86: 345-391.
24. Chung HJ. 2012. Physiological activity of acai berry (Euterpe oleracea Mart.) extracted with different solvents. Korean J Food Cult 27: 75-81.
25. Fleury C, Mignotte B, Vayssière JL. 2002. Mitochondrial re- active oxygen species in cell death signaling. Biochimie 84:
131-141.
26. MacKenzie SH, Clark AC. 2008. Targeting cell death in tu- mors by activating caspases. Curr Cancer Drug Targets 8:
98-109.
27. Decker P, Muller S. 2002. Modulating poly (ADP-ribose) polymerase activity: potential for the prevention and therapy of pathogenic situations involving DNA damage and oxida- tive stress. Curr Pharm Biotechnol 3: 275-283.
28. Kaufmann T, Strasser A, Jost PJ. 2012. Fas death receptor
signalling: roles of Bid and XIAP. Cell Death Differ 19:
42-50.
29. Tummers B, Green DR. 2017. Caspase-8: regulating life and death. Immunol Rev 277: 76-89.
30. Fulda S, Debatin KM. 2006. Extrinsic versus intrinsic apop- tosis pathways in anticancer chemotherapy. Oncogene 25:
4798-4811.
31. Kroemer G, Galluzzi L, Brenner C. 2007. Mitochondrial membrane permeabilization in cell death. Physiol Rev 87:
99-163.
32. Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM.
1998. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J Biol Chem 273: 5858-5868.
33. Satoh T, McKercher SR, Lipton SA. 2013. Nrf2/ARE-medi- ated antioxidant actions of pro-electrophilic drugs. Free Radic Biol Med 65: 645-657.
34. Loboda A, Damulewicz M, Pyza E, Jozkowicz A, Dulak J.
2016. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: an evolutionarily conserved mechanism. Cell Mol Life Sci 73: 3221-3247.
35. Gęgotek A, Skrzydlewska E. 2015. The role of transcription factor Nrf2 in skin cells metabolism. Arch Dermatol Res 307: 385-396.
36. Schäfer M, Werner S. 2015. Nrf2-A regulator of keratinocyte redox signaling. Free Radic Biol Med 88: 243-252.