Toll-like Receptor 4-mediated Apoptotic Cell Death in Primary Isolated Human Cervical Cancers
Jinyoung Won1,2,3, Yunkyung Hong2,3,4, Sookyoung Park2,3,4, Joo-Heon Kim5* and Yonggeun Hong1,2,3,4*
1Department of Rehabilitation Science, Graduate School of Inje University, Gimhae 50834, Korea
2u-Healthcare & Anti-aging Research Center (u-HARC), Inje University, Gimhae 50834, Korea
3Biohealth Products Research Center (BPRC), Inje University, Gimhae 50834, Korea
4Department of Physical Therapy, College of Healthcare Medical Science & Engineering, Inje University, Gimhae 50834, Korea
5Institute of Animal Medicine, College of Veterinary Medicine, Gyeongsang National University, Jinju 52828, Korea Received January 4, 2018 /Revised March 12, 2018 /Accepted March 17, 2018
Toll-like receptor 4 (TLR4) has been implicated in cell proliferation and apoptosis in several types of cancer. In this study, the impact of TLR4 activation on apoptotic cell death in gynecologic cancers in- duced by lipopolysaccharide (LPS) was investigated. Cervical cancer cell lines were produced from isolated surgical specimens supplied by Paik Hospital. The primary cultures of normal myometrium and gynecologic cancers, including cervical, endometrial, and ovarian cancers, were used to examine the differences in morphological characteristics between normal and cancerous cells. A reverse tran- scription polymerase chain reaction analysis was used to determine the relative expression levels of TLR4 gene involved in apoptosis-associated signaling in cervical cancer cells. The cancer cell colonies showed a tendency to reach high levels of confluency compared with normal cells. In addition, an enhanced growth rate and loss of contact inhibition were observed in gynecologic cancer cells com- pared with normal cells (doubling times of 16.6 hr vs. 26 hr, respectively). The expression level of ITGA5, an alpha-5 integrin marker, was upregulated in normal myometrial cells, but this tendency was not exhibited in cervical cancer cells. Furthermore, p53 tumor suppressor gene expression was upregulated, whereas TLR4 and caspase-3 gene expressions were downregulated in cervical cancer cells. Notably, the expression levels of TLR4 and caspase-3 were increased significantly in LPS-treated cancer cells compared with those in non-LPS-treated cells. These results suggest that the TLR4-mediated caspase-dependent apoptotic signaling pathway could be suggested as a therapeutic target for the treatment of gynecologic cancers, including cervical cancers.
Key words : Apoptosis, gynecologic cancer, lipopolysaccharide, proliferation, Toll-like receptor
*Corresponding authors
*Tel : +82-55-320-3681, Fax : +82-55-329-1678
*E-mail : [email protected] (Yonggeun Hong)
*Tel : +82-55-772-2344, Fax : +82-55-772-2308
*E-mail : [email protected] (Joo-Heon Kim)
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2018 Vol. 28. No. 6. 718~725 DOI : https://doi.org/10.5352/JLS.2018.28.6.718
Introduction
Gynecologic cancers, such as cervical, endometrial, and ovarian cancers, cause serious morbidity and mortality.
Moreover, they are common malignancies in women [31].
There are an estimated 1 million new cases of and 500,000 deaths due to gynecologic cancers annually worldwide [20, 26]. Of all gynecologic cancers, uterine cervix cancer is a ma- jor cause of death in woman, after breast, endometrial, and
ovarian cancer [5]. While gynecologic cancer mortality has decreased due to improvements in cancer therapies, such as chemotherapy, radiation therapy, and surgery, additional research is needed for the development of therapeutic strat- egies to address the risks of infertility and spontaneous abor- tion [4].
Toll-like receptors (TLRs) play important roles in early innate immune defense mechanisms through recognition of pathogen-associated molecular patterns (PAMPs) [32]. TLRs 1-13 have been identified in human and mouse, which are evolutionarily conserved in terms of responses to specific classes of microbes [7]. Most TLRs have been found to trig- ger responses after recognition of PAMPs or microbe-asso- ciated molecular patterns (MAMPs). Damage-associated mo- lecular patterns (DAMPs), produced by death signals, also stimulate TLR-dependent signal transduction [24]. The main function of the TLRs is to promote the innate immune re- sponse and trigger the adaptive immune response. Recent
studies have reported that the activation of TLRs, induced by TLR agonists, promotes tumor-targeting immune re- sponses effectively [14].
TLR-mediated immunotherapy is currently being inves- tigated in the treatment of some cancers. In animal cancer models, the use of TLR agonists, such as TLR1/TLR2 ago- nists (e.g., bacterial lipoprotein; BLP), and TLR7/TLR8 ago- nists (e.g., imiquimod) has induced tumor regression [27, 28, 36]. Additionally, a TLR4 agonist (lipopolysaccharide; LPS) has shown anti-tumor effects via inhibition and delaying tu- mor growth [10].
Recent studies have reported the importance of TLR4 in tumor suppression pathways through induction of pro-in- flammatory cytokine secretion [15]. Activation of TLR4, in- volved in apoptotic signaling, has been shown to protect against the development of cancer. TLR4 signaling is modu- lated via TLR adaptor protein which is composed of a toll/interleukin-1 receptor (TIR) domain. In TLR4 signaling, myeloid differentiation primary response gene 88 (MyD88)- dependent and TIR-domain containing adaptor inducing be- ta interferon (TRIF)-dependent pathways are essential for ac- tivation of transcription factor, nuclear factor kappa B (NF-κ B). Activation of TLR4-induced NF-κB then triggers multiple signal pathways, including those involved in immunity, in- flammation, apoptosis, proliferation, and stress responses [17].
In this study, we established primary cultured cells from gynecologic cancers and normal myometrium tissue to ex- amine the morphology of cancerous and normal cells. We also investigated the role of TLR4 in apoptotic cell signaling in cancer to assess whether the molecular mechanism of TLR4-induced apoptosis might provide therapeutic targets for cancer treatment.
Materials and Methods
Animals
The protocols of animal study were approved by the Institutional Animal Care and Use Committee (IACUC) at Inje University (approval no. 2014-50), and conducted in ac- cordance with the Inje University Animal Care guidelines and the Korean Department of Agriculture. All animals were maintained under a 12 hr light/dark cycle with free access to laboratory chow and drinking water. Sprague–Dawley (SD) rats weighing 180-200 g (6-week-old) were used in this experiment.
Cell culture
Primary cultured cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Lonza, Walkersville, MD, USA), supplemented with 10% fetal bovine serum (FBS;
HyClone, Logan, UT, USA) and penicillin/streptomycin (60 μg/ml; Lonza). Cultured cells were incubated at 37℃ in a humidified atmosphere of 5% CO2, and the culture medium was refreshed 2 or 3 times per week.
Primary culture system
Primary cancer cells were isolated from patients with gy- necologic malignancies. This procedure was approved by our institutional ethics committee. Gynecologic cancer tis- sues, including cervical, endometrial, and ovarian cancers, were obtained from patients with gynecologic conditions be- ing treated at Paik Hospital. The average age of the patients was 45.3 years. These samples were minced into 1 mm3 pieces in Roswell Park Memorial Institute 1640 medium (RPMI 1640 medium; Lonza) using sterilized micro-scissors.
The minced tissue was placed in a conical tube containing RPMI 1640 and 2 mg/ml collagenase type II, and incubated at room temperature for 15 min in a shaking incubator. After digestion, the cancer tissues were centrifuged at low speed (600-800 rpm, 3 min) and the supernatant was removed. The cancer tissues were then subjected to further digestion with 0.025% trypsin-0.02% ethylenediaminetetraacetic acid (EDTA) at room temperature for 10 min in a shaking incubator. Digestion was stopped by washing the tissue 3 times with RPMI 1640. Gynecologic cancer cells were main- tained in RPMI 1640 supplemented with 10% FBS, penicillin (60 μg/ml), and streptomycin sulfate (100 μg/ml).
Myometrial cells isolated from uterine myometrial smooth muscle were used as ‘normal’ cells. Normal my- ometrial smooth muscle was obtained from female SD rats (6-week-old, weighing 180-200 g). Rat uterine tissue was ob- tained and placed in a sterile Petri dish containing cold DMEM. Uterine tissue was separated from the uterine horn to the uterine cervix and minced in DMEM into 1-mm3 pieces using sterilized micro-scissors. The minced tissue was placed in a conical tube containing DMEM and 2 mg/ml collagenase type II at room temperature for 30 min in a shak- ing incubator. After digestion, the myometrial tissue was centrifuged at low speed (600-800 rpm, 3 min) and the super- natant was removed. Myometrial cells were immersed in 0.025% trypsin-0.02% EDTA (room temperature, 20 min, shaking incubator). Myometrial cells were then collected by
Table 1. Sequence-specific primers used for RT-PCR
Gene Primer sequence (5'-3') Product length (bp) GenBank accession No.
ITGA5 F: GGAAGGGACGGAGTCAGTGT
R: ACTGGATAGGACTGGTGCCC 118 NM_001108118
p53 F: CCGCGCCATGGCCATCT
R: GCGCTCATGGTGGGGG 100 NM_030989
TLR4 F: CATTCCTTTAAGTTGCCTGA
R: ATATTTGCCAAGCAATTGAG 200 NM_019178
Caspase-3 F: GGGTCATGGTTCATCCAGTC
R: GTCCAGGGAGAAGGACTCAA 140 NM_012922
Gapdh F: GTATGACTCCACTCACGGCAAA
R: GGTCTCGCTCCTGGAAGATG 100 BC094037
low-speed centrifugation (600-800 rpm, 3 min), and washed 3 times with DMEM. Cultured primary myometrial cells were maintained in DMEM containing 10% FBS, penicillin (60 μg/ml), and streptomycin sulfate (100 μg/ml) [21, 22].
Cell proliferation assay
To assess the growth curve of normal myometrial cells and uterine cervix cancer cells, cell counting was performed with a hemocytometer (Marienfeld Laboratory Glassware, Lauda-Königshofen, Germany). Cells were plated at 4×104 cells per 100 mm plate and incubated for 72 hr. Cell pellets were collected and resuspended in DMEM. Cells were stained with an equal volume of 0.4% trypan blue solution (Sigma-Aldrich) to determine the number of dead cells. Cells were counted with the hemocytometer every 24 hr, trypan blue-excluding and -positive cells.
Chemical treatment
Cells (5×105/ml) were stimulated with 1 μg/ml LPS.
Additionally, the cells were subjected to pretreatment with either vehicle or LPS in DMEM containing 10% FBS and pen- icillin/streptomycin (60 μg/ml). Cell culture medium that contained vehicle or LPS was preincubated at 37℃ in 5%
CO2.
RNA isolation and reverse transcription polymerase chain reaction (RT-PCR) analysis
RNA isolation was performed using the TRIzol® reagent (Sigma-Aldrich); total RNA was extracted according to the manufacturer’s protocol. The concentration of RNA was de- termined with a spectrophotometer (Mecasys, Daejeon, Korea). RNA (5 μg) was reverse-transcribed using reverse transcriptase (Invitrogen, Carlsbad, CA, USA). The primers used are listed in Table 1. RT-PCR was performed using
standard reaction conditions. PCR was conducted in the fol- lowing steps: denaturation at 95℃ for 30 s, annealing at 60
℃ for 1 min, and elongation at 72℃ for 1 min. Up to 40 PCR cycles were performed. The reaction mixture consisted of 0.5 μM of the forward and reverse primers, 1.5 mM MgCl2, 0.2 mM of each deoxynucleotide (dNTP), and 1.25 U of GoTaq® polymerase (Promega, Fitchburg, WI, USA). After amplification, the PCR products were electrophoresed on a 1.2% agarose gel and stained with ethidium bromide. The intensities of the amplified bands were quantified with the Gel Doc System and Quantity One® software (Bio-Rad Laboratories, Richmond, CA, USA). The ImageJ program was used to quantify the density (ver. 1.6; NIH, Bethesda, MD, USA).
Statistical analysis
Data were subjected to statistical analyses using Student’s t-tests or a one-way analysis of variance (ANOVA) followed by post hoc Tukey tests. Data are presented as means ± stand- ard deviation (SD). A p-value less than 0.05 was considered to indicate statistical significance.
Results
Isolation and characterization of primary cells from gynecologic cancers
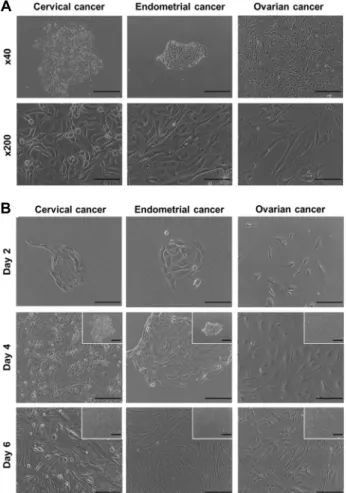
Cervical, endometrial, and ovarian cancers are the most lethal gynecologic malignancies in woman. Cancerous tis- sues were obtained from patients at Paik Hospital; we iso- lated and cultured primary cervical, endometrial, and ovar- ian cancer cells after collagenase type II treatment (Fig. 1A).
Morphological characterizations of primary cultured cells were made with an inverted microscope ~6 d after culture initiation. Colonies of cancer cells attached and started to
A
B
Fig. 1. Primary cultures of gynecologic cancer cells. (A) Primary cells were isolated from gynecologic cancer tissues, sup- plied by Paik Hospital. Scale bars = 500 μm (40×), 100 μm (200×). (B) Comparison of morphological character- ization of primary culture gynecologic cancer cells, in- cluding cervical, endometrial, and ovarian cancer cells.
Scale bars = 500 μm (40×), 200 μm (100×).
spread, with cell proliferation (Fig. 1B). The density of gyne- cologic cancer cells reached 100% confluency more rapidly than did the normal cells (data not shown).
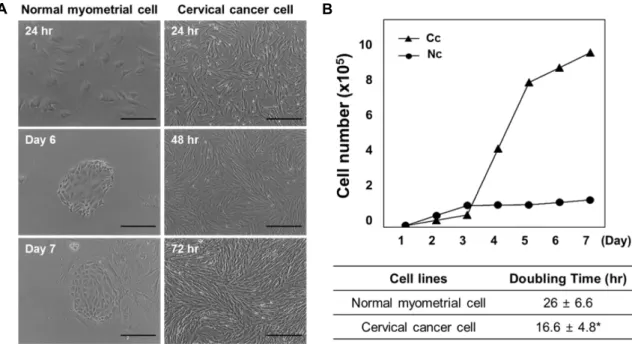
Analysis of the cell growth curve and proliferation of cervix cancer cells
Primary cultured cells were monitored to determine their proliferation after initial seeding density (5×104/ml).
Cervical cancer cells showed active proliferation and loss of contact inhibition, with cellular overlap occurring within 72 hr (Fig. 2A). While the cancer cells rapidly reached high con- fluency within 72 hr, normal cells showed a largely constant basal density level of <50% until Day 7.
To confirm the growth curve and proliferation of normal versus cancer cells, cell counting was performed with a he-
mocytometer every 24 hr. The cell doubling time was calcu- lated from the following formula: N = N0×2t/T, where N0 is the initial concentration of cells, N is the final concen- tration of cells, t is the duration of the culture, and T is the doubling time. Cervical cancer cells showed more rapid growth than normal myometrial cells over 7 d. The cell dou- bling times of cervical cancer and normal myometrial cells were 16.6 hr and 26 hr, respectively (Fig. 2B).
Comparison of the gene expression levels of normal myometrium and cervical cancer
To determine changes in gene expression in normal and cancerous cells, RT-PCR analysis was performed for the in- tegrin alpha 5 (ITGA5), p53, toll-like receptor 4 (TLR4), Caspase-3 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) genes using total RNA samples isolated from cul- tured primary cells (Fig. 3). The expression of ITGA5, an al- pha5 integrin marker, was upregulated in myometrial cells but not in uterine cervix cancer cells. Expression of ITGA5 was observed primarily in smooth muscle, including my- ometrium and endometrium tissues. Additionally, expre- ssion of Caspase-3, related to apoptosis marker, and TLR4 was seen in normal cells but not in cancer cells. However, the expression of p53, related to tumor suppression, was in- creased in cancer cells versus normal cells.
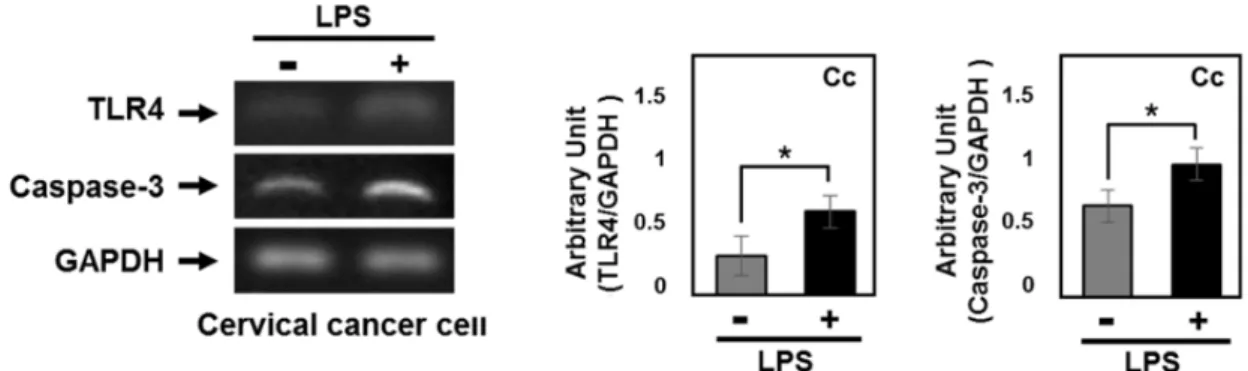
The effect of LPS induced TLR4 activation on apop- totic signaling pathway in cervical cancer
To examine the effect of LPS-mediated TLR4 activation on apoptotic signaling in gynecologic cancer, cervical cancer cells (5×105/ml) were stimulated with LPS (1 μg/ml) or ve- hicle in DMEM containing 10% FBS and pen- icillin/streptomycin (60 μg/ml). Factors associated with tu- mor suppression were differentially expressed after LPS treatment (Fig. 4). Increased TLR4 and Caspase-3 expression was observed in LPS-treated cancer cells versus vehicle treat- ed cancer cells. These results indicated that the activation of TLR4, mediated by LPS, was associated with the cas- pase-dependent apoptotic pathway.
Discussion
The diagnosis and treatment of cancer have been revolu- tionized by recent developments in medicine. Molecular- based targeted therapy is currently under investigation as a potential therapeutic approach in the treatment of cancer.
A B
Fig. 2. Cell growth curves and doubling times of primary culture cells. (A) Normal myometrial cells and cervical cancer cells were assessed with an inverted microscope. Imaging of cancer cells revealed active proliferation, as compared with normal cells.
Scale bars = 200 μm (100×). (B) Doubling times and cell numbers were measured to assess cell proliferation. Nc, normal myometrial cells; Cc, cervical cancer cells. *p<0.05 vs. normal myometrial cells.
Fig. 3. Comparison of the gene expression patterns of normal cells and cervical cancer cells. Reverse transcription polymerase chain reaction (RT-PCR) analysis of the integrin alpha 5 (ITGA5), p53, toll-like receptor 4 (TLR4), Caspase-3 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) genes using total RNA samples isolated from cultured primary cells. Nc, normal my- ometrial cells; Cc, cervical cancer cells. *p<0.05 vs. normal myometrial cells.
Cancer-specific genes, known as oncogenes, are activated in different cancers [6]. Additionally, genetic variations, includ- ing dysfunction of tumor suppressor genes, also increase the risk of some cancers [19]. A major hallmark of cancer is re- sistance to apoptotic cell death induced by activation of on-
cogenic signaling pathways [8]. This also has the potential to trigger certain normal cells to become cancerous, which is then associated with inappropriate proliferation and meta- static spread of cancer cells.
Several recent studies have reported on distinctive clinical
Fig. 4. The effect of LPS treatment on apoptosis in cervical cancer cells. The expression levels of TLR4 and Caspase-3 in cervical cancer cells. Cervical cancer cells were stimulated with lipopolysaccharide (LPS). *p<0.05 vs. non LPS treated group.
and pathological features of gynecologic cancers. Most gyne- cologic cancers show metastatic tendencies and a pro- gressive malignant state. Gene-based comparative analyses are currently being conducted in gynecologic cancer re- search, but gene expression profiling has mostly failed to specifically characterize gynecologic cancers, likely due to their diversity [3, 13, 36].
In this study, we established gynecologic cancer cell line from cancer biopsy specimens including the uterine cervix, endometrium and ovary. Small clusters from cancer speci- mens were observed within 12-24 hr after primary culture initiation. Colonies of cancer cells attached and spread with- in 3-4 d after initial seeding. In normal cells, cell viability and adhesion were similar to that of cancer cells. However, cancer cells displayed higher cell densities with limited con- tact inhibition [12]. Because many cancer cells are not sensi- tive to extracellular growth factors, they are largely un- affected by density-dependent inhibition [34]. Most normal cells, unlike cancer cells, stop proliferating and initiate cell cycle arrest before reaching confluent [1].
We identified characteristic doubling times and growth rates in gynecologic cancer cells. Uncontrolled growth and proliferation occur commonly in cancers when the cells reach confluence. However, the growth of normal cells is arrested upon contact with each other. Previous studies have reported that the cell density at which normal cells stop pro- liferating is influenced by growth factor requirements [9].
Because of the loss of growth factor requirements in cancer cells, they can continue to proliferate independent of cell confluency [35]. Moreover, some cancer cells produce growth factors by autocrine mechanisms, stimulating cell di- vision [30].
We compared gene expression patterns of normal my- ometrial cells with those of cervical cancer cells using RT-
PCR. We focused on the characterization of cervical cancer cell that showed significantly altered cell morphology and accelerated cell proliferation compared with primarily cul- tured normal cells. The colony formation and attachment of cervical cancer cells was persistently observed after 72 hr.
Under normal state, p53 play an important role in the cel- lular response by regulating growth arrest and cell death [2]. The p53 gene is commonly mutated in cancer that devel- ops resistance to apoptosis [18, 23]. In present study, in- creased expression of p53 and accelerated cell proliferation were observed in cervical cancer cells. Although over- expression of p53 was increased in cervical cancer cells, p53-dependent cell cycle arrest and cell death were not oc- curred, suggesting that increased transcriptional activation of p53 may associated with altered structure and function.
While the expression of p53 related to tumor suppression was increased, the expression of TLR4 and Caspase-3 was downregulated in cancer cells compare with normal cells.
Other studies have reported that the expression of p53 and TLR4 is commonly increased in different types of human cancers, but the role of TLR4 is still unclear and controversial [29, 33]. We used cultured primary cancer cells to define the molecular action of TLR4 in the variety of cellular responses.
Expression of TLR4 was higher in LPS treated cancer cells than non LPS treated cells. The present study also demon- strated that the expression of Caspase-3, which is identified as a key mediator of apoptosis involved in cellular DNA damage, was upregulated through LPS-mediated TLR4 activation. The presence of Caspase-3 indicates the induction of apoptotic irreversible cell death [16, 25].
In previous study, we evaluated the cell viability, pro- liferation, and cell cycle using MTT and trypan blue assay and visualized the total nuclei using in vitro BrdU in- corporation assay to investigate the relationship between
apoptosis, autophagy, and senescence in colorectal cancer cells [11]. Based on the previous our study, we measured the doubling time of cancer and normal cells and compared gene expression pattern, which involved in pro-apoptotic cell death such as Caspase-3 and p53, to determine the effect of LPS mediated TLR4 activation on apoptotic cell death.
Therefore, these findings suggest that LPS induced TLR4 ac- tivation is related with Caspase-3 mediated apoptotic cell death, and TLR4 signal pathway may be useful target for anti-cancer drug development.
The present study provides evidence for the involvement of TLR4-mediated apoptosis in gynecologic cancers. Further studies should focus on the role of TLR signaling in cancers, in association with the induction of immunotherapy.
Acknowledgment
This study was supported by the KRIBB Research Initiative Program (KGM4611714 to Y. Hong), Republic of Korea.
References
1. Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K. and Walter, P. 2002. The Cell Cycle and Programmed Cell Death.
Mol. Biol. Cell. pp. 983-1026, 4th ed., Garland Science, NY, USA.
2. Aubrey, B. J., Kelly, G. L., Janic, A., Herold, M. J. and Strasser, A. 2018. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell.
Death. Differ. 25, 104-113.
3. Biewenga, P., Buist, M. R., Moerland, P. D., Ver Loren van Themaat, E., van Kampen, A. H., ten Kate, F. J. and Baas, F. 2008. Gene expression in early stage cervical cancer.
Gynecol. Oncol. 108, 520-526.
4. Chhabra, S. and Kutchi, I. 2013. Fertility preservation in gy- necologic cancers. Clin. Med. Insights Reprod. Health 7, 49-59.
5. Fleck, J. L., Pavel, A. B. and Cassandras, C. G. 2016. Integrat- ing mutation and gene expression cross-sectional data to in- fer cancer progression. BMC. Syst. Biol. 10, 12.
6. Fukata, M., Chen, A., Klepper, A., Krishnareddy, S., Vama- devan, A. S., Thomas, L. S., Xu, R., Inoue, H., Arditi, M., Dannenberg, A. J. and Abreu, M. T. 2006. Cox-2 is regulated by Toll-like receptor-4 (TLR4) signaling: Role in prolifera- tion and apoptosis in the intestine. Gastroenterology 131, 862- 877.
7. Fulda, S. 2013. Regulation of cell death in cancer-possible implications for immunotherapy. Front. Oncol. 3, 29.
8. Goustin, A. S., Leof, E. B., Shipley, G. D. and Moses, H.
L. 1986. Growth factors and cancer. Cancer Res. 46, 1015- 1029.
9. He, W., Liu, Q., Wang, L., Chen, W., Li, N. and Cao, X. 2007.
TLR4 signaling promotes immune escape of human lung cancer cells by inducing immunosuppressive cytokines and apoptosis resistance. Mol. Immunol. 44, 2850-2859.
10. Hanahan, D. and Weinberg, R. A. 2011. Hallmarks of cancer:
the next generation. Cell 144, 646-674.
11. Hong, Y., Won, J., Lee, Y., Lee, S., Park, K., Chang, K. T.
and Hong, Y. 2014. Melatonin treatment induces interplay of apoptosis, autophagy, and senescence in human color- ectal cancer cells. J. Pineal. Res. 56, 264-274.
12. Iguchi, Y. Ito, Y. M., Kataoka, F., Nomura, H., Tanaka, H., Chiyoda, T., Hashimoto, S., Nishimura, S., Takano, M., Yamagami, W., Susumu, N., Aoki, D. and Tsuda, H. 2014.
Simultaneous analysis of the gene expression profiles of can- cer and stromal cells in endometrial cancer. Genes Chromo- somes Cancer 53, 725-737.
13. Iribarren, K., Bloy, N., Buqué, A., Cremer, I., Eggermont, A., Fridman, W. H., Fucikova, J., Galon, J., Špíšek, R., Zitvogel, L., Kroemer, G. and Galluzzi, L. 2015. Trial Watch:
Immunostimulation with Toll-like receptor agonists in can- cer therapy. Oncoimmunology 5, e1088631.
14. Janeway, C. A. Jr. and Medzhitov, R. 2002. Innate immune recognition. Annu. Rev. Immunol. 20, 197-216.
15. Kawai, T. and Akira, S. 2007. Signaling to NF-kappaB by Toll-like receptors. Trends Mol. Med. 13, 460-469.
16. Kiraz, Y., Adan, A., Kartal Yandim, M. and Baran, Y. 2016.
Major apoptotic mechanisms and genes involved in apopto- sis. Tumour. Biol. 37, 8471-8486.
17. Liu, Y., Hu, X., Han, C., Wang, L., Zhang, X., He, X. and Lu, X. 2015. Targeting tumor suppressor genes for cancer therapy. Bioessays 37, 1277-1286.
18. Maxwell, S. A. and Davis, G. E. 2000. Differential gene ex- pression in p53-mediated apoptosis-resistant vs. apopto- sis-sensitive tumor cell lines. Proc. Natl. Acad. Sci. USA. 97, 13009-13014.
19. Minig, L., Padilla-Iserte, P. and Zorrero, C. 2016. The rele- vance of gynecologic oncologists to provide high-quality of care to women with gynecologic cancer. Front. Oncol. 5, 308.
20. Mitchell, J. A., Shynlova, O., Langille, B. L. and Lye, S. J.
2004. Mechanical stretch and progesterone differentially reg- ulate activator protein-1 transcription factors in primary rat myometrial smooth muscle cells. Am. J. Physiol. Endocrinol.
Metab. 287, 439-445.
21. Palmberg, L. and Thyberg, J. 1986. Uterine smooth muscle cells in primary culture. Alterations in fine structure, cy- toskeletal organization and growth characteristics. Cell Tissue Res. 246, 253-262.
22. Piccinini, A. M. and Midwood, K. S. 2010. DAMPening in- flammation by modulating TLR signalling. Mediators In- flamm. 2010, pii: 672395.
23. Pistritto, G., Trisciuoglio, D., Ceci, C., Garufi, A. and D'Orazi, G. 2016. Apoptosis as anticancer mechanism: function and dysfunction of its modulators and targeted therapeutic strategies. Aging 8, 603-619.
24. Preedy, V. R. and Watson, R. R. 2010. Handbook of Disease Burdens and Quality of Life Measures, pp. 803-823, Spring-
초록:부인과질환 특이적 종양의 TLR4 매개성 apoptosis 유발에 관한 연구
원진영1,2,3․홍윤경2,3,4․박수경2,3,4․김주헌5*․홍용근1,2,3,4*
(1인제대학교 대학원 재활과학과, 2인제대학교 u-항노화헬스케어연구소(u-HARC), 3인제대학교 바이오헬스 소재
연구센터(BPRC), 4인제대학교 보건의료융합대학 물리치료학과, 5경상대학교 수의과대학 동물의학연구소)
Toll 유사수용체의 TLR4는 세포자연사(apoptosis)와 관련하여 세포의 생존과 증식에 영향을 미치는 것으로 알 려져 있다. 본 연구에서는 TLR4의 활성이 부인과 질환 특이적 종양세포의 세포사멸기작에 어떠한 영향을 미치는 지 살펴보았다. TLR4의 활성에 의한 세포자연사를 확인하기 위하여 부인암 조직(자궁경부암, 자궁내막암, 난소암) 에서 종양세포를 분리하여 초대배양시스템을 구축하였고, lipopolysaccharide (LPS)에 의한 TLR4의 활성유도 과 정에서 종양세포의 형태학적 변화를 살펴보았다. 또한, TLR4 매개성 세포사멸 기작을 확인하기 위하여 역전사 중합효소 연쇄반응(RT-PCR)을 통해 유전자 분석을 진행하였다. 연구 결과, 부인암의 초대배양세포에서 세포접촉 저지(contact inhibition)현상이 감소되었고, 세포의 배가시간(doubling time)이 단축되어, 종양세포의 성장률 변화 를 확인하였다(p<0.05). 자궁근육층(정상조직)의 초대배양세포에서는 민무늬근육 확인 인자인 ITGA5 (an alpha5 integrin marker)의 유전자 발현이 나타났으나, 자궁경부조직의 초대배양세포에서는 발현변화를 확인할 수 없었 다. 종양세포의 유전자분석 결과에서 p53과 같은 종양억제인자의 발현이 유의적으로 증가한 반면(p<0.05), 세포사 멸 신호기작과 관련하여 TLR4와 Caspase-3의 발현은 감소하였다(Caspase-3, p<0.05). LPS를 처리한 종양세포에서는 LPS 비처리군과 비교 시, TLR4의 발현증가와 함께 Caspase-3의 발현변화가 동반되었다. 이러한 결과들은 TLR4 매개성 apoptosis 유도가 종양세포의 증식억제에 중요한 영향을 미치는 것을 의미하며, TLR4 신호기작을 이용한 종양세포의 새로운 치료적 접근법을 제시할 것으로 기대한다.
er, NY, USA.
25. Radogna, F., Dicato, M. and Diederich, M. 2015. Cancer- type-specific crosstalk between autophagy, necroptosis and apoptosis as a pharmacological target. Biochem. Pharmacol.
94, 1-11.
26. Scott, S. H. and Bahnson, B. J. 2011. Senescence marker pro- tein 30: functional and structural insights to its unknown physiological function. Biomol. Concepts 2, 469-480.
27. Smits, E. L., Ponsaerts, P., Berneman, Z. N. and Van, Tendeloo, V. F. 2008. The use of TLR7 and TLR8 ligands for the enhancement of cancer immunotherapy. Oncologist 13, 859-875.
28. Soussi, T. 2007. p53 alterations in human cancer: more ques- tions than answers. Oncogene 26, 2145-2156.
29. Sporn, M. B. and Roberts, A. B. 1985. Autocrine growth fac- tors and cancer. Nature 313, 745-747.
30. Suh, D. H., Kim, M., Kim, H. J., Lee, K. H. and Kim, J.
W. 2016. Major clinical research advances in gynecologic cancer in 2015. J. Gynecol. Oncol. 27, e53.
31. Tan, R. S., Ho, B., Leung, B. P. and Ding, J. L. 2014. TLR
cross-talk confers specificity to innate immunity. Int. Rev.
Immunol. 33, 443-453.
32. Tyson, J. J. and Novak, B. 2014. Control of cell growth, divi- sion and death: information processing in living cells.
Interface Focus 4, 20130070.
33. Wang, Y., Weng, Y., Shi, Y., Xia, X., Wang, S. and Duan, H. 2014. Expression and functional analysis of Toll-like re- ceptor 4 in human cervical carcinoma. J. Membr. Biol. 247, 591-599.
34. Witsch, E., Sela, M. and Yarden, Y. 2010. Roles for growth factors in cancer progression. Physiology (Bethesda) 25, 85- 101.
35. Yin, J. G., Liu, X. Y., Wang, B., Wang, D. Y., Wei, M., Fang, H. and Xiang, M. 2016. Gene expression profiling analysis of ovarian cancer. Oncol. Lett. 12, 405-412.
36. Zhang, Y., Luo, F., Li, A., Qian, J., Yao, Z., Feng, X. and Chu, Y. 2014. Systemic injection of TLR1/2 agonist improves adoptive antigen-specific T cell therapy in glioma-bearing mice. Clin. Immunol. 154, 26-36.