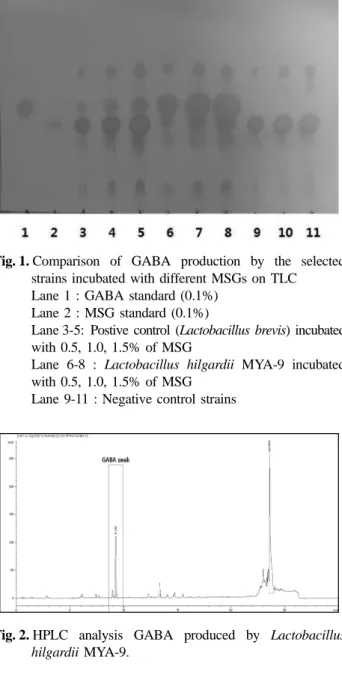
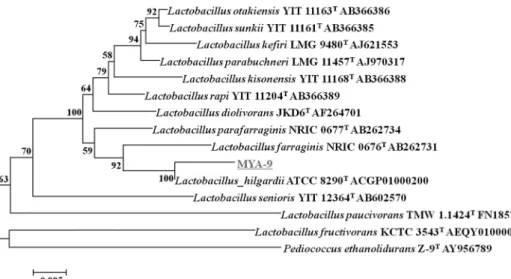
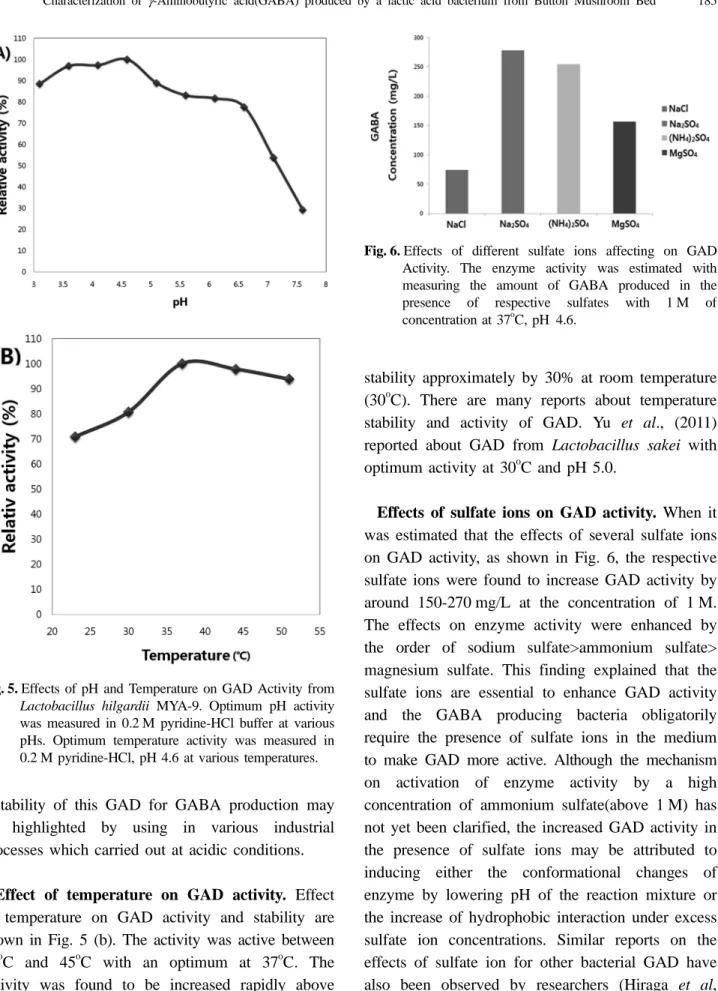
관련 문서
CBS73 strain isolated from domestic Korean traditional food able to produce antimicrobial peptide having high antibacterial effect, low molecular weight, stable over wide
This study takes about 2-3 days to analyze, which is much shorter than the existing fuming nitric acid method, so it is easy to analyze a large amount of samples and will be
In the third stage, strictly anaerobic methane fermentation, biogas was produced by methanogenic bacterium using organic acids including acetic acid.. Biogas consisted
- A novel lactic acid bacterium for the production of high purity l-lactic acid, Lactobacillus paracasei subspL. helveticus - Production of Lactic Acid from Water Hyacinth
The purpose and necessity of this study was to investigate the effects of lactic acid fatigue and body oxidation through myofascial relaxation exercise using small tools
Probiotic properties of lactic acid bacteria isolated from mukeunji, a long-term ripened kimchi. Journal of Food
Effect of amino acid addition in culture media on the mycelial growth and g-aminobutyric acid (GABA) production of Sparassis latifolia..
It was disclosed from the results that the color extracted from eucalyptus leaves was reddish yellow tone that could be manifested in a wide spectrum of