46(3) : 195∼ 202 (2015)
195
벌사상자 Ethyl Acetate 분획물의 항염증활성연구
강옥화#·김상영#·권동렬* 원광대학교 약학대학 한약학과 한약연구소
Ethyl Acetate Fraction of Cnidium monnieri(L). Cussion Suppresses PAM plus A23187-induced Inflammation Reaction through Blockade of NF- κB
and MAPK activation
Ok-Hwa Kang#, Sang-Young Kim# and Dong-Yeul Kwon*
Department of Oriental Pharmacy, College of Pharmacy and Wonkwang-Oriental Medicines Research Institute, Wonkwang University, Iksan, Jeonbuk 570-749, Korea
Abstract − Cnidium monnieri (L). Cussion is used as a tonic agent in traditional oriental medicine. However, the molecular mechanism of mast cell-mediated anti-inflammatory modulation has not been fully understood. The aim of the present study was to demonstrate the effects of Cnidium monnieri (L). Cussion eathyl acetate fraction on the expression of pro-inflammatory cytokines, as well as to elucidate its mechanism of action in the human mast cell line (HMC-1). Cells were stimulated with phorbol 12-myristate 13-acetate (PMA) plus A23187 in the presence or absence of Cnidium monnieri (L). Cussion eathyl ace- tate fraction. Cnidium monnieri (L). Cussion eathyl acetate fraction significantly inhibited the PMA plus A23187-induction of inflammatory cytokines such as tumor necrosis factor (TNF)-α, interleukin (IL)-6 and IL-8. Moreover, EtOAc fraction atten- uated cyclooxygenase (COX)-2 expression. In activated HMC-1 cells, phosphorylation of extra-signal response kinase (ERK) 1/2 decreased after treatment with EtOAc fraction. Moreover EtOAc fraction inhibited PMA plus A23187-induced nuclear fac- tor (NF)-κB activation, IκB degradation. EtOAc fraction suppressed the expression of TNF-α, IL-6, IL-8 through a decrease in the ERK 1/2, as well as activation of NF-κB. These results indicated that Cnidium monnieri (L). Cussion EtOAc fraction exerted a regulatory effect on inflammatory reactions mediated by mast cells.
Key words − Cnidium monnieri (L). Cussion, Mast cell, Anti-inflammatory effect, MAPKs, NF-κB
사상자(Cnidii Fructus)는 벌사상자 Cnidium monnieri(L).
Cussion 또는 사상자 Torilis japonica Decandolle의 열매 로 산형과( 形科; Umbelliferae)에 속한 일년생 초본인 벌사상자(뱀도랏)의 성숙한 과실을 건조한 것으로, 여름과 가을에 과실이 성숙할 때 채취하여 이물을 제거하고 햇볕 에 말려 사용한다.1)
벌사상자는 한방에서 성미가 신온(辛溫)하여 산한(散寒) 하고 조양(助陽)하며, 고조(苦燥)하여 제습시키므로 지양시 키는 효능이 있고, 내복하면 온신조양(溫腎助陽)시키는 효 능이 있어 양위(陽 )와 궁냉(宮冷)의 병증을 치료하고, 외 용하면 조습살충(燥濕殺蟲)시키는 효능이 있어 음양대하(陰
痒帶下)를 치료한다.2) 벌사상자의 유용성분으로 coumarin 계열의 화합물인 osthole과 xanthotoxol이 그 중 가장 많은 활성이 보고되고 있으며 약리작용으로는 면역기능개선작용 과 천식억제작용, 골다공증 예방 및 건망증을 개선하고, 파 동적 피부과민반응과 접촉성 피부염을 억제 및 백혈병 세 포에 대해서 세포독성을 나타낸다고 보고되었다.3)
염증반응은 병원균에 감염되거나 자극을 받았을 때 면 역체계가 일으키는 최초의 반응으로서 홍반, 발열, 통증 및 장기의 기능장애를 동반하는 선천적 면역반응의 일종이다.
따라서 자극이 가해지면 국소적으로 histamine, serotonine, prostaglandins, leukotriene과 같은 혈관 활성 물질이 유리 되어 혈관 투과성이 증대되면서 염증을 유발한다. 그러나 지속적인 염증반응은 도리어 점막손상을 촉진하고, 그 결 과 일부에서는 암이 유발되기도 한다.4)비만세포는 염증성
#These two authors contributed equally to this work.
*교신저자(E-mail):[email protected] (Tel):+82-63-850-6802
질환에 작용하여 각종 사이토카인을 분비하는데, 그 중에 서도 IL-6, IL-8과 Tumor necrosis factor(TNF)-α 등의 증 가는 다양한 염증성 질환을 악화시키는 요인으로 잘 알려 져 있다. IL-6는 T와 B림프구의 증식과 활성자극 분화 및 급성기 단백 합성을 하며, 특히, 외상에 대한 염증 유발이 강하다.5) IL-8의 발현은 비만세포, 섬유아세포, 내피 및 외 피 세포 등의 다른 조직이나 세포뿐 아니라 IL-1α, β 또는 TNF-α와 같은 전염증성 chemokine 또는 lipopolysaccharide 와 같은 전염증성 제제에 감응하여 빠르게 유도된다는 것 이 밝혀졌다.6) 또 다른 보고는 피부병, 관절염, 사구체 신 염에 있어 IL-8 중화항체가 유익한 효과가 있음을 나타냈 다. TNF-α는 여러 면역촉진제에 반응하여 비만세포에 의 해 발생하는 cytokine이다. cytokine을 동물이나 인체에 투 여하면 염증, 발열, 심장혈관계 효과, 출혈, 혈액 응고, 급 성 감염, 쇼크 상태에서 나타나는 반응과 유사한 급성단계 반응을 유발한다. 과도하거나 조절되지 않은 TNF-α의 생 성은 다수의 질병상태와 관련되어 있다. TNF-α는 전 염 증 활성을 지니고 있는데, 이것은 TNF-α의 염증 초기단 계와 함께 심근경색, 발작, 순환계 쇼크를 포함하는 몇몇 심각한 질병에서 조직 손상의 중개자로 작용하는 것으로 보인다. 세포 간 부착 분자나 내피 백혈구 부착 분자와 같 은 부착 분자의 TNF-α의 유도성 발현이 특히 중요한 것 으로 보인다.7) 그러므로 TNF-α의 감소는 많은 염증성, 감 염성, 면역성 악성 질환들의 치료에 대한 유용한 전략이 된다. NF-κB는 iNOS, COX-2, cytokine, chemokine 등의 분비와 관련된 유전자의 전사를 유도할 수 있다.8) 대부분 의 세포에서 NF-κB의 형태는 p50과 p65로 구성된 heterodimer이다. 보통 NF-κB는 내생성 억제분자인 IκB 단백질과 결합한 상태로 세포질에서 존재한다. NF-κB가 활성화됐을 때 세포핵을 전위시키고, 복잡한 염증성 유전 자를 활성화 한다.9) MAPKs는 세포외 자극을 세포막에서 부터 세포내 핵까지 전달하는 대표적인 신호전달 경로로 잘 알려져 있다. MAPKs는 성장호르몬, cytokine, stress 등 의 수용체로부터 활성화된 신호를 세포내로 전달하여 세 포의 증식, 분화, 사멸 등 다양한 기능을 담당한다. MAPK 는 ERK 1/2, JNK 1/2, p38 MAPKs 세 가지로 분류를 할 수가 있다. ERK 1/2는 세포질에 위치하며, 인산화반응에 의해 활성화된다. 성장호르몬의 신호전달에 주로 관여하며 세포전사활동, 세포증식 및 분화에 중요한 역할을 한다. 반 면에 stress kinase로 분류되는 p38과 JNK 1/2는 이름 그 대로 세포 외부의 스트레스성 자극에 의해 활성화 되며 염 증반응, 세포사멸 등을 매개한다고 알려져 있다.10)
사상자는 항염증,11) 항균,12)피부주름 예방13)과 멜라닌 생성 억제14) 관련 효과가 강조되어 보고되고 있으며, 특히 벌사상자는 온신조양(溫腎助陽)의 효능이 있어서 남자의 양 위(陽 )와 음낭습양(陰囊濕痒), 여자의 대하음양(帶下陰痒)
과 한냉불잉(寒冷不孕) 등을 치료함으로써15)성호르몬 조 절,16)발기부전17) 등과 같은 불임치료에 사용되어 불임관 련 효과에 대한 연구는 많은 보고가 있었으나, 항염증과 관련된 연구는 진행되지 않았다. 이에 본 연구에서는 비만 세포에서의 벌사상자 에틸아세테이트 분획물에서 염증성 cytokine의 억제를 통해 항염증효과와 그 메커니즘을 알아 보고자 한다.
재료 및 방법
약재 − 본 실험에 사용된 벌사상자는 (주)백제허브(대전, 한국)에서 구입하고, 원광대 한약학과(권동렬 교수)에서 감 정한 것을 사용하였으며, 표준품은 원광대학교 한약학과 본초학실험실의 표본실에 보관하였다.
시약 및 기기 − 세포배양에서 필요한 Iscove’s Modified Dulbecco’s Media(IMDM) 배지는 Welgene(Daegu, Korea) 제품을, Fetal bovine serum(FBS)는 Hyclone(New zealand) 제품을 구입해 사용하였다. Phorbol 12-myristate 13- acetate(PMA), Calcium Ionophore A23187, Penicillin- Streptomycin, Bovine Serum Albumin(BSA)는 Sigma(St.
Louis, Mo, USA)에서, CellTiter 96® AQueous One Solution Cell Proliferation Assay(MTS)는 Promega(Madison, Wisconsin, USA)에서, Human IL-6, IL-8 ELISA Set는 BD Biosciences(San Diego, USA)에서, Western blot 실험 에서 필요한 Protein Extraction Solution은 iNtRON Biotechnology(Gyeonggi-do, Korea)에서, Ponceau S solution은 Sigma(St. Louis, Mo, USA), EzReprobe은 ATTO(Tokyo, Japan)에서 구입하였다. 항체인 ERK 1/2, JNK 1/2, p38, COX-2, β-actin, NF-κB, IκBα, p-IκBα, anti- rabbit은 Santa Cruz Biotechnology(Santacruz, CA, US)에서, anti-mouse IgG은 Life-Technonlogies와 anti-goat IgG은 kirkegaard &Perry Laboratories Inc.에서, Easy Blue total RNA Extraction kit는 iNtRON Biotechnology(Gyeonggi-do, Korea), Reverse Transcription kit 는 Giagen (Valencia, CA, USA), Sensi 2 × PCR premix은 Lugen, Gel red, 6 × loading buffer는 Dynebio(Gyeonggi-do, Korea), Primer는 Bioneer(Daejeon, Korea)에서, Nuclear and Cytoplasmic Extraction Reagents는 Thermo Scientifeic(MA, USA)에서 구입해 사용하였다.
세포배양 − 실험에 사용한 사람 비만세포주 HMC-1는 항생제(100 U/ml Penicillin, 100 U/ml Streptomycin)와 10% FBS가 첨가된 IMDM 배지에서 37oC, 5% CO2 incubator에 적응시켜 배양하였다. 계대 배양은 이틀에 한 번씩 하였다.
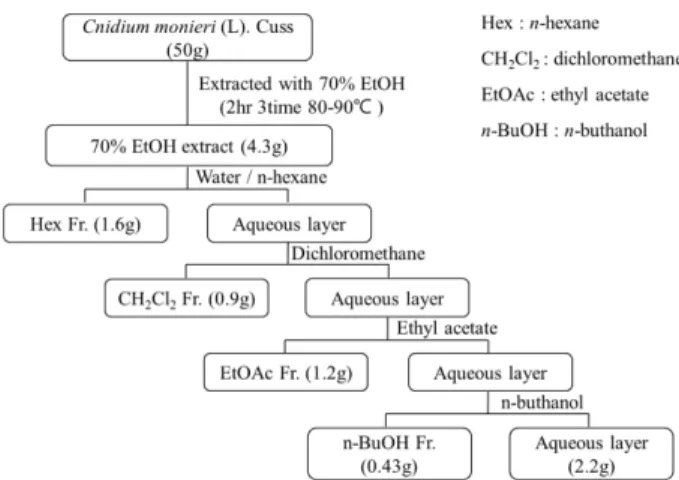
추출물의 제조 − 벌사상자 50 g과 70% EtOH 500 ml를 넣어 2시간씩 3회 추출한 다음 감압 농축하여 동결 건조
한 후, 벌사상자 70% EtOH 추출물(4.3 g)을 얻었다. 추출물 을 물에 현탁시킨 후, 극성이 낮은 용매부터 극성이 높은 용 매 순으로 용매분획을 각각 3회씩 반복 수행하여 감압농축, 동결 건조 한 후, n-hexane(1.6 g), Dichloromethane(0.9 g), EtOAc(1.2 g), n-BuOH(0.43 g) 분획물을 얻었다. 시료는 실 험에 사용하기 전 밀봉하여 냉동보관(-20oC) 하였다(Scheme 1).
MTS assay − 세포독성은 MTS assay를 이용하여 확인 하였다. HMC-1을 96 well plate에 5×104cells/well 로 분 주한 후, 30분 뒤 시료를 농도별로 처리하였다. 24시간 동 안 배양한 후, MTS solution을 20 µl씩 첨가한 후, 37oC, 5% CO2 incubator에서 2시간 배양하였다. Microplate reader를 이용하여 490 nm에서 흡광도를 측정하였다.
ELISA assay- HMC-1을 48 well plate에 5×105cells/
well 로 분주하였고, 30분 뒤 시료를 각 각 처리한 후, 1 시간 뒤 PMA(50 nM)와 A23187(1 µM)를 처리 하고 8시 간 뒤, 세포를 가라앉게 한 후, 배양 상층액을 수거하였다.
4oC에 하룻밤 동안 있던 1차 항체 coating plate를 다음날, 30분 실온 상태로 둔 뒤, 3번 wash buffer로 세척 후, Assay buffer를 200 µl 분주하였다. 1시간 뒤 3번 세척하
였다. sample과 standard를 100 µl 분주 후, 2시간 실온 상 태로 두었다. 5번 세척하고, working Detector(2차 항체 + SA-HRP) 100µl 분주 후, 1시간 실온 상태로 두었다. 7번 세척 후, Substrate Solution을 100 µl 분주하고, 암실에서 최대 30분 기다렸다. 마지막으로, Stop Solution(1M H3PO4)을 50 µl 분주한 후, Microplate reader를 이용하여 450 nm에서 흡광도를 측정하였다.
RNA 분리 및 RT-PCR − HMC-1을 6 well plate에 3×106cells/well 로 분주하였고, 30분 뒤 시료를 각 각 50, 100 250µg/ml 처리한 후, 1시간 뒤 PMA(50 nM), A23187(1µM)로 자극하고 6시간 뒤, 세포를 수집하였다.
PBS로 세척하여 Easy Blue 1 ml을 넣고 용해시킨 뒤, Chloroform을 400 µl 첨가하고 vortexing 후, 원심 분리 (13,000 rpm, 10분)하였다. 상층액 400 µL을 새로운 tube 에 옮기고, isopropanol을 동량 넣어주었다. tube을 2회 inverting 하고 실온에 10분간 두었다. 원심분리(13,000 rpm, 5분)하였다. 상층액을 버리고, 70% EtOH 1 ml을 넣고 2 회 세척한 후, 건조시켜 DEPC가 처리된 증류수에 용해시 킨 후, Nano Drop 기계로 A260/A280 nm의 비율이 1.6~2.0 범위 내의 값을 갖는 RNA 시료를 실험에 사용하 였다.
cDNA 합성은 Reverse Transcription kit를 사용하였다.
cDNA에 IL-6, IL-8, GAPDH, COX-2 Primer를 넣고 유 전자 증폭기를 이용하여 증폭시켰다. 2% Agarose Gel에 Gel red를 1:10,000 희석시켜 전기영동 한 후, UV 검출기 로 발현 정도를 확인하였다. RT-PCR을 위해 여러 개의 primer를 사용하였다. Primer의 구성은 다음과 같다(Table 1).
핵과 세포질 추출 및 Western blot analysis- 핵과 세포 질 추출은 Nuclear and Cytoplasmic Extraction Reagents 를 사용하였다. cell volume이 20 µl 일 때, CER 을 200µl 넣고 pellet을 15초 동안 최고로 세게 vortexing 하 였다. 그리고 얼음에 10분 두고, CER 를 11 µl 넣고 5초 Scheme 1. Fractionation of Cnidium monnieri (L). Cussion.
Table I. Primer sequences for RT-PCR
cDNA Primer sequences
IL-6 Forward 5'-GATGGATGCTTCCAATCTGGAT-3' Reverse 5'-AGTTCTCCATAGAGAACAACATA-3' IL-8 Forward 5'-TGTGCTCTCCAAATTTTTTTTACTG-3'
Reverse 5'-CTCTCTTTCCTCTTTAATGTCCAGC-3' TNF-a Forward 5'-CACCAGCTGGTTATCTCTCAGCTC-3’
Reverse 5'-CGGGACGTGGAGCTGGCCGAGGAG-3’
COX-2 Forward 5'-TTCAAATGAGATTGTGGGAAAATTGCT-3' Reverse 5'-AGATCATCTCTCTCCTGAGTATCTT-3' GAPDH Forward 5'-CCATGTTCGTCATGGGTGTGAACCA-3'
Reverse 5'-GCCAGTAGAGGCAGGGATGATGTTC-3' Primer sequences for RT-PCR
동안 최고로 vortexing 하였다. 얼음에 1분간 둔 뒤, 5초 동안 최고로 vortexing 한 후, 원심분리(13,000 rpm, 5분) 하였다. 즉시, 상층액(세포질 추출)을 새로운 tube에 옮겨 담았다. 남아있는 pellet에 NER을 100 µl 넣고, 15초 동안 최고로 vortexing 하였다. 얼음에 sample을 40분간에 두는 데, 10분마다 15초 동안 vortexing 하였다. 40분 뒤, 원심 분리(13,000 rpm, 10분)하였다. 즉시 상층액(핵 추출)을 새 로운 tube에 옮겨 담았다.
HMC-1를 6 well plate에 5×106cells/well로 분주하고, 30분 뒤 시료를 50, 100, 250 µg/mL 처리한 후, 1시간 뒤 PMA(50 nM), A23187(1µM)로 자극하고 1시간 뒤, 세포 를 수집하였다. PBS로 세척 한 뒤, lysis buffer를 200 µl 넣고, 20분 동안 lysis 한 후, 원심 분리(13,000 rpm, 5분) 하였다. 단백질 정량은 BSA와 Bio-Radprotein Assay Kit 를 사용하였고, microplate reader를 이용하여 595 nm에서 흡광도를 측정하고 정량하였다. 만들어진 단백질을 12%
SDS-PAGE로 변성 분리하여, 이를 PVDF (polyvinylidene difluoride) membrane에 15 V에서 1시간 동안 Transfer하 였다. 그리고 Ponceau S solution으로 염색 한 후, Transfer 가 잘 되었는지 확인하였다. 염색을 TBS-T(TBS + 0.05%
Tween 20)용액으로 세척한 후, membrane을 5% skim milk가 첨가된 TBS-T 용액에 상온에서 1시간 동안 blocking 하였다. 1차 항체 반응은 1차 항체를 5% skim milk에 희석하여 4oC에서 하룻밤 동안 수행하였다. TBS- T로 3회 15분씩 세척하였다. 2차 항체 반응은 HRP(horse radish peroxidase)가 결합된 anti-mouse IgG, anti-rabbit IgG를 1:2,000으로 희석하여 상온에서 2시간 반응시킨 뒤, TBS-T로 3회 15분씩 세척하여 ECL Substrate에 반응 후, ImageQuantTM LAS 4000 mini을 이용하여 단백질 발현량 을 분석하였다.
통계분석 − 모든 실험은 3회 이상 반복으로 이루어졌으
며, 실험결과는 각 항목에 따라 평균치±표준편차(SEM)를 구하여 그 유의성은 Student’s t-test(SPSS. ver.19) 분석법 을 이용하여 신뢰수준 95%( p<0.05)에서 통계적 유의차를 평가하였다.
결과 및 고찰
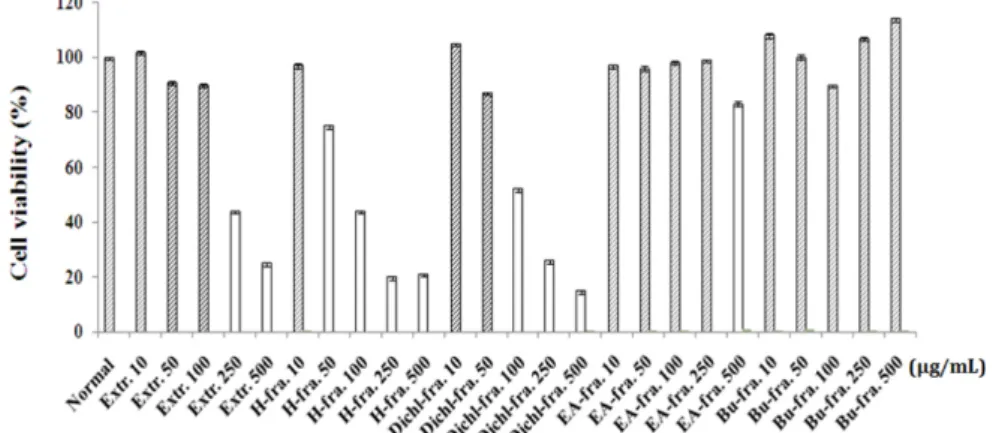
벌사상자가 세포 생존에 미치는 영향을 측정하기 위해, MTS assay를 통해 확인하였다. 실험 결과, 추출물(10, 50, 100µg/ml), n-hexane 분획물(10 µg/ml), Dichloromethane 분획물(10, 50 µg/ml), EtOAc 분획물(10, 50, 100, 250 µg/ml), n-BuOH 분획물(10, 50, 100, 250, 500 µg/ml)에서 85~100%
의 생존율을 보여 세포 독성을 나타내지 않음을 확인하였 다(Fig. 1). 인간 비만세포주인 HMC-1 세포를 PMA(phorbol 12-myristate 13-acetate)와 A23187(calcium ionophore)로 자극 하였을 때 in vitro 실험모델에서, 염증반응 매개인자 를 연구하는데 잘 알려져 있다.18) PMA와 A23187는 세포 질 내의 Ca2농도 상승에 의해 각종 Ca2감수성 조절인자 에 작용하여 일련의 비만세포 활성화 반응에 관여한다. 염 증반응에서 관여하는 세포들 간의 복합적 상호작용은 cytokines 이라고 불리는 단백군에 의해 매개된다. 특히, IL-6은 비만세포에서 생성되어, 조혈작용과 염증을 조절하 는데 관여하는 다양한 기능을 하는 것으로 알려져 있다.5) IL-8은 호산구 화학주성인자이며, 염증 반응에 있어서 중 요한 역할을 하는 chemokines이다.6)본 연구에서, PMA와 A23187로 활성화된 HMC-1에서 벌사상자 추출물과 분획 물을 처리하여 전염증성 사이토카인의 생성양을 측정해본 결과, EA 분획물에서 IL-6, IL-8과 TNF-α의 생성을 모두 억제하였다(Fig. 2). 특히, EA 분획물 250 µg/ml에서 현저 하게 억제되었음을 확인하였다. 따라서 전염증성 사이토카 인의 생성억제 효과를 보인 벌사상자 EA 분획물을 이용
Fig. 1. Cell viability of HMC-1 cells. Cell viability was measured by MTS assay. HMC-1 cells were treated with various con- centrations of Extration and Fraction for 24 hours. Date represent the mean±SD values of triplicate determinations from three separate experiment.
하여 IL-6, IL-8, TNF-α의 mRNA 발현정도를 본 결과, IL-6, IL-8, TNF-α mRNA 발현이 농도 의존적으로 억제 되었으며, 250 µg/ml에서 현저한 억제양상을 확인하였다 (Fig. 3). 한편, COX-2는 염증과정 및 종양 발생에서 중요 한 역할을 하며, COX-2의 발현은 염증반응에서 중요한 지 표로 알려져 있다. 본 연구에서는, PMA와 A23187로 활 성화된 HMC-1에서 벌사상자 EA 분획물을 처리하였을 때, COX-2 mRNA 발현이 농도 의존적으로 억제되었음을 확
인하였다(Fig. 4). 그리고 염증반응은 주로 MAPKs 분자 들의 활성화로 인한 NF-κB 같은 전사조절인자들의 가동 으로 시작된다. MAPKs의 subfamily에는 대표적으로 ERK, JNK, p38가 있으며, 이들의 신호전달과정은 염증물질의 합 성을 유도하는 점에서 염증반응과정에서 중요한 역할을 한
다.19,20)실험 결과, PMA와 A23187을 처리한 HMC-1 세
포에서 ERK경로를 경유하며, 벌사상자 EA 분획물을 함 께 처리한 HMC-1에서는 ERK 인산화를 억제시켰음을 확 Fig. 2. Effects of Extration and Fraction on IL-6, IL8 and TNF-α Production in PMA (50 nM)+A23187 (1 µM)-stimulated HMC-1 cells. TNF-α Production was measured by ELISA assay. HMC-1 cells were treated with IMDM or various concentrations in the presence of PMA+A23187 for 8 hours. Date are mean±SD values of triplicate determinations from three separate experiment.
*P<0.05
인하였다(Fig. 5).
또한, NF-κB는 염증성 cytokines 이나 면역세포 이동에 중요한 접합 분자들, chemokines 등의 유전자를 발현시키
기 때문에 초기 염증반응 조절에 중요한 전사인자이다.
NF-κB 전사인자들 중에, IκBα는 phosphorylation이 되면 ubiquitin-proteasome 체계에 의해 분해되어 IκB 농도가 감 Fig. 3. Effect of EtOAc Fraction on Cytokines mRNA in PMA (50 nM)+A23187 (1 µM)-stimulated HMC-1 cells. Expression of mRNA were analyzed by RT-RCR. HMC-1 cells were treated with IMDM or various concentrations (50, 100 and 250µg/ml) of EtOAc Fra. in the presence of PMA+A23187 for 6 hours. The experiment was repeated three times, and similar results were ascer- tained. (A) cytokines gene expression (B) Gene expression ratio.
Fig. 4. Effect of EtOAc Fraction on Cytokines mRNA in PMA (50 nM)+A23187 (1 µM)-stimulated HMC-1 cells. Expression of mRNA were analyzed by RT-RCR. HMC-1 cells were treated with IMDM or various concentrations (50, 100 and 250µg/ml) of EtOAc Fra. in the presence of PMA+A23187 for 6 hours. The experiment was repeated three times, and similar results were ascer- tained. (A) COX-2 gene expression (B) Gene expression ratio.
Fig. 5. Effect of EtOAc Fraction on the Phosphorylation of MAPKs in PMA (50 nM)+A23187 (1 µM)-stimulated HMC-1 cells.
Phosphorylation of MAPKs were analyzed by western blot analysis. HMC-1 cells were treated with IMDM or various concen- trations (50, 100 and 250µg/ml) of EtOAc Fra. in the presence of PMA+A23187 for 1 hour. The experiment was repeated three times, and similar results were ascertained. (A) protein expression (B) protein expression ratio.
소하게 된다. 그 결과, IκBα의 phosphorylation으로 NF- κB가 활성화가 되어, 염증매개 인자들의 합성과 생성에 영 향을 미치게 된다고 보고되어 있다.21,22) 실험 결과, 벌사상 자 EA 분획물은 PMA와 A23187로 활성화된 HMC-1의 세 포질 내의 IκBα의 phosphorylation과 IκBα의 degradation이 억제하였다. 따라서 벌사상자 EA 분획물이 핵 내의 NF-κB 의 신호전달 경로를 차단함을 확인하였다(Fig. 6).
본 연구결과를 종합하면, 벌사상자 EA 분획물은 PMA 와 A23187로 활성화된 인간비만세포에서 신호전달경로인 MAPKs에서의 ERK의 phosphorylation과 NF-κB 경로를 저해함으로써, IL-6, IL-8, TNF-α, COX-2 발현에 영향을 미치는 것으로 관찰되었다.
결 론
인간 비만세포주인 HMC-1 세포를 PMA와 A23187로 자극 하였을 때 벌사상자 EA 분획물의 항염증 효과를 조 사하여 다음과 같은 결론을 얻었다.
1. HMC-1 세포를 PMA와 A23187로 자극하여 벌사상 자 추출물과 분획물을 처리하였을 때, EA 분획물에서 IL- 6, IL-8, TNF-α은 250 µg/ml에서 현저하게 억제되었다.
2. PMA와 A23187로 활성화된 HMC-1에서 벌사상자 EA 분획물을 처리하였을 때, COX-2 mRNA 발현이 농도 의존적으로 억제되었다.
3. PMA와 A23187을 처리한 HMC-1 세포에서 벌사상 자 EA 분획물을 처리하였을 때 ERK 인산화를 억제시켰 음을 확인하였다.
4. 벌사상자 EA 분획물은 PMA와 A23187로 활성화된 HMC-1의 세포질 내의 IκBα의 phosphorylation과 IκBα의 degradation이 억제 하였으며, 핵 내의 NF-κB의 신호전달 경로를 차단하였다.
이와 같은 결과로 보아 벌사상자 EA 분획물은 인간비
만세포에 작용하여 신호전달경로인 ERK의 phosphorylation 과 NF-κB 경로를 저해함으로써, IL-6, IL-8, TNF-α, COX-2 발현에 영향을 미치는 것으로 관찰되었으며, 이는 비만세포를 매개로한 염증 질환 치료제로서의 가능성이 있 음을 알 수 있다.
사 사
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2013R1A1A2064673), by the Korea government (MSIP) (2008-0062484).
인용문헌
1. Ministry of food and drug safety (2012) Publication of English Version of National Standard of Traditional Medic- inal(Herbal and Botanical) Materials. fifth edition. 173.
2. Hwang, E. I., Lee, S. K., So, S. H., Koo, B. S., Han, G. H. and Kim, N. M. (2007) Isolation and development of quantitative determination of Torilin from the Torilis Fructus. Kor. J.
Pharmacogn. 38: 321-326.
3. Pharmacognosy teaching materials Compilation Committee (2008) Pharmacognosy, 324-325. Dong Myeong Publishers, Seoul.
4. Willoughby, D. A. (1975) Heberden Oration, 1974. Human arthritis applied to animal models. Towards a better therapy.
Ann. Rheum. Dis. 34: 471-478.
5. Hirano, T., Akira, S., Taga, T. and Kishimoto, T. (1990) Bio- logical and clinical aspects of interleukin 6. Immunol. Today 11: 443-449.
6. Mitsuyama, K., Toyonaga, A., Sasaki, E., Watanabe, K., Tateishi, H., Nishiyama, T., Saiki, T., Ikeda, H., Tsuruta, O.
Fig. 6. Effect of EtOAc Fra. on the activation of NF-κB in PMA (50 nM)+A23187 (1 µM)-stimulated HMC-1 cells. Activation of NF-κB were analyzed by western blot analysis. HMC-1 cells were treated with IMDM or various concentrations (50, 100 and 250µg/ml) of EtOAc Fra. in the presence of PMA+A23187 for 1 hour (cytoplasm) or 2 hours (nuclear). The experiment was repeated three times, and similar results were ascertained. (A) protein expression (B) protein expression ratio.
and Tanikawa, K. (1994) IL-8 as an important chmoattractant for neutrophils in ulcerative colitis and Crohn's disease. Clin.
Exp. Immunol. 96: 432-436.
7. Munro, J. M., Pober, J. S. and Cotran, R. S. (1989) Tumor necrosis factor and interferon-gamma induce distinct patterns of endothelial activation and associated leukocyte accumu- lation in skin of Papio anubis. Am. J. Pathol. 135: 121-133.
8. Barnes, P. J. and Adcock, I. M. (1997) NF-kappa B: a pivotal role in asthma and a new target for therapy. Trends. Phar- macol. Sci. 18: 46-50.
9. Barnes, P. J. and Karin, M. (1997) Nuclear factor-kappa B: a pivotal transcription factor in chronic inflammatory diseases.
New. Engl. J. Med. 336: 1066-1071.
10. Caivano, M. (1999) Role of MAP kinase cascades in induc- ing arginine transporters and nitric oxide synthetase in RAW 264.7 macrophages. FEBS. Lett. 429: 249-253.
11. Lee, E. B., Kim, S. M. and Kim, T. H. (1998) Anti-inflam- matory activities of Torilis japonica fruit. Kor. J. Pharma- cogn. 29: 384-390.
12. Cho, W. I., Choi, J. B., Lee, K., Chung, M. S. and Pyun, Y.
R. (2008) Antimicrobial activity of torilin isolated from Tori- lis jponica fruit against Bacillus subtilis. J. Food. Sci. 73:
M37-46.
13. Koo, B. S., Hwang, E. I., So, S. H., Lee, S. K., Han, G. H. and Kim, N. M. (2007) Effect of Torilis Fructus on procollagen biosynthesis and activity of Matrix Methalloproteinase- I(MMP-1) in human dermal fibroblast. Kor. J. Pharmacogn.
38: 349-353.
14. Yun, C. Y., Kim, D., Lee, W. H., Park, Y. M., Lee, S. H., Na, M., Jahng, Y., Hwang, B. Y., Lee, M. K., Han, S. B. and Kim, Y. (2009) Torilin from Torilis japonica inhibits melanin pro-
duction in alpha-melanocyte stimulating hormone-activated B16 melanoma cells. Planta Med. 5: 1505-1508.
15. Herbology Editorial Committee of Korean Medicine Schools.
(1998) Herbology[Boncho-hak], 576-577. Younglim Press, Seoul.
16. Yan, F., Liang, Z., Jianna, C., Zhengtao, W., Losahan, X. and Zhengxing, Z. (2001) Analysis of Cnidium monnieri fruits in different regions of China. Talanta 53: 1155-1162.
17. Chen, C. Y. (2009) Computational screening and design of traditional Chinese medicine(TCM) to block phosphodi- esterase-5. J. Mol. Graph. Model. 28: 261-269.
18. Hosoda, M., Yamaya, M., Suzuki, T., Yamada, N., Kamanaka, M., Sekizawa, K., Butterfield, J. H., Watanabe, T., Nishimura, H. and Sasaki, H. (2002) Effects of rhinovirus infection on histamine and cytokine production by cell lines from human mast cells and basophils. J. Immunol. 169: 1482- 1491.
19. Shakoory, B., Fitzgerald, S. M., Lee, S. A., Chi, D. S. and Krishnaswamy, G. (2004) The role of human mast cell- derived cytokines in eosinophil biology. J. Interferon Cytok- ine Res. 24: 271-281.
20. Crossthwaite, A. J., Hasan, S. and Williams, R. J. (2002) Hydrogen peroxide-mediated phosphorylation of ERK1/2, Akt/PKB and JNK in cortical neurones: dependence on Ca(2+) and PI3-kinase. J. Neurochem. 80: 24-35.
21. Klemm, S. and Ruland, J. (2006) Inflammatory signal trans- duction from the Fc epsilon RI to NF-kappa B. Immuno- biology 211: 815-820.
22. Kriete, A. and Mayo, K. L. (2009) Atypical pathways of NF- κB activation and aging. Exp. Gerontol. 44: 250-255.
(2015. 8. 13 접수; 2015. 9. 14 심사; 2015. 9. 21 게재확정)