https://doi.org/10.20307/nps.2018.24.3.171
171
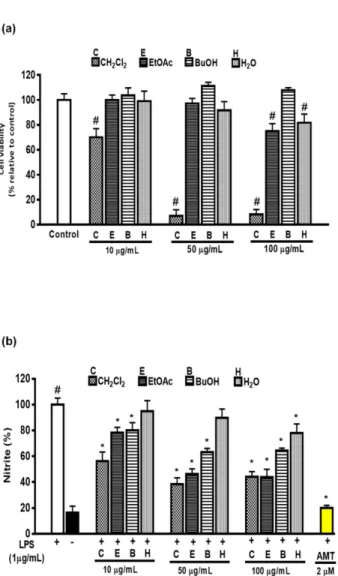
Anti-inflammatory Potential of Artemisia capillaris and Its Constituents in LPS-induced RAW264.7 Cells
Qudeer Ahmed Abdul
1, Su Hui Seong
1, Bo Ra Ahn
1, Md Nurul Islam
2, Hyun Ah Jung
3,*, and Jae Sue Choi
1,*
1
Department of Food and Life Science, Pukyong National University, Busan 48513, Republic of Korea
2
Department of Pharmacy, Mawlana Bhashani Science and Technology University, Santosh, Tangail 1902, Bangladesh
3