세리포리아 락세라타 균사체 배양물이 고지방식이로 급이된 당뇨병 마우스의 인슐린 저항성에 미치는 영향
최진우1*․신은지1*․이세진1․김용환1․김소라1․지유미1․김나연1․안창훈2․이인한2․김윤수1
1(주)퓨젠바이오파마
2(주)퓨젠셀텍
Effects of Submerged Culture of Ceriporia lacerata Mycelium on High Fat Diet-Induced Diabetic Mice with Insulin Resistance
Jin Woo Choi1*, Eun Ji Shin1*, Sae Jin Lee1, Yong Hwan Kim1, So Ra Kim1, Yu Mi Ji1, Na Yeon Kim1, Chang Hun An2, In Han Lee2, and Yoon Soo Kim1
1FugenbioPharma Co., Ltd.
2FugenCelltech Co., Ltd.
ABSTRACT We investigated the effects and regulatory mechanism of submerged culture of Ceriporia lacerata myce- lium (FBS0P) on high fat diet-induced diabetic mice with insulin resistance and C2C12 myotubes. In order to examine the in vivo effects of treatment with FBS0P, FBS0P (500 mg/kg/d) was orally administered to high fat-fed male mice for 4 weeks. An oral glucose tolerance test was performed by measuring blood glucose levels following an oral glucose load. In the results, the FBS0P-treated mice group showed improved insulin sensitivity while FBS0P treatment reduced serum insulin and c-peptide levels, compared to the high fat diet group. High fat diet-induced mice with FBS0P showed increasing adenosine monophosphate-activated protein kinase (AMPK) phosphorylation and glucose transporter 4 (GLUT4) expression levels in gastrocnemius muscles. To determine the down-stream mechanism, C2C12 myotubes were incubated with and without FBS0P. Treatment with FBS0P increased AMPK phosphorylation and glucose uptake by inducing GLUT4 translocation in C2C12 myotubes. Furthermore, FBS0P improved insulin sensitivity in tumor necrosis factor α-treated myotubes. The present investigation demonstrates that FBS0P from submerged culture of C. lacerata mycelium exhibits antihyperglycemic efficacy both in a cell culture system and in a high fat-fed diabetic mice model by overcoming insulin resistance.
Key words: Ceriporia lacerata, diabetes, high fat diet-induced diabetic mice, insulin resistance, natural products
Received 29 September 2017; Accepted 16 November 2017 Corresponding author: Yoon Soo Kim, FugenbioPharma Co., Ltd., Seoul 06746, Korea
E-mail: [email protected], Phone: +82-2-3463-5402
*These authors contributed equally to this work.
서 론
인슐린 저항성(insulin resistance)은 인슐린이 정상적으 로 분비되거나 초과로 분비되지만 양에 비해 인슐린의 작용 이 효과적으로 이루어지지 않는 상태를 나타낸다. 비만일 경우 인슐린 저항성에 의해 포도당 흡수가 저해되고 결국 제2형 당뇨병으로 진행되게 된다. 인슐린 저항성의 발생기 전은 인슐린 신호 전달 과정, adenosine monophosphate activated protein kinase(AMPK) 인산화 과정 등의 이상에 의한 것이며, 이는 제2형 당뇨병의 내당능장애에 중요한 요 소가 된다(1-3).
포도당이 체내에 유입되면 췌장의 베타세포에서 인슐린이
분비되어 세포막의 insulin receptor(IR)와 결합하여 세포 내 tyrosine kinase가 활성화되고, insulin receptor sub- strate 1(IRS1), insulin receptor substrate 2(IRS2)가 인 산화된다. 인산화된 IRS1은 phosphatidylinositol 3-kin- ase(PI3K), phosphatidylinositol 3,4,5-trisphosphate, protein kinase B(Akt)를 순차적으로 인산화시키는데, 이때 Akt의 인산화로 인하여 포도당 수송체인 glucose trans- porter 4(GLUT4)가 세포질에서 세포막으로 이동(GLUT4 translocation)하여 포도당 흡수를 증가시키게 된다(4-6).
인슐린 신호 전달 과정에서 IRS1, Akt 등에 이상이 생겨 GLUT4 translocation에 장애가 생기면 고혈당이 유발되어 당 독성이 발생하고 이로 인해 인슐린 저항성이 초래되어 고인슐린혈증을 야기하는 것으로 보고되고 있다(7,8).
AMPK는 인슐린 신호 전달 경로와 별개로 인산화를 통해 GLUT4 translocation을 증가시켜 세포 내 당 유입을 증가 시키는 것으로 알려져 있으며(9), 세포 내 에너지 대사를 조 절하는 역할을 한다(10). 세포 내 ATP가 고갈되어 AMP/
Table 1. Diet compositions in experimental groups (%)
Ingredients CD1) HFD2)
Starch Sucrose Cellulose Proteins Fat
Vitamin and mineral mixture
44.9 3.2 4.1 15.2 2.9 3.9
8.4 20.1 5.8 20.0 24.0 6.4
1)Fed control diet group.
2)Fed high fat diet group.
ATP 비율이 증가하게 되면 AMPK가 활성화되어 지방산의 산화와 해당과정을 촉진하고, 이를 통해 미토콘드리아 내 ATP 생산을 증가시킨다(11).
2003년 일본에서 처음 발견되어 보고된(12) Ceriporia lacerata는 백색부후균 중 하나로 목재의 셀룰로오스, 헤미 셀룰로오스, 리그닌 등을 분해하여 생물학적 환원의 역할을 한다(13). 한편 C. lacerata의 secondary metabolites는 현 재까지 많이 연구가 되어 있지는 않지만 새로운 tremulane sesquiterpenoids(14), lanostane triterpenes(15), 그리 고 flavonoid(16) 등의 구조들이 보고되어 있다. C. lacer- ata의 의약학적 연구로 제1형 당뇨병 쥐에서의 항당뇨 효과 (17), 인슐린 분비 세포 보호 효과(18), 제2형 당뇨에서의 항당뇨 효과와 인슐린 신호 전달상의 기전 연구(19) 등이 보고되어 있다.
본 논문에서는 유전자 변형으로 당뇨병을 유도한 동물 모 델보다 실제 사람에서의 제2형 당뇨병 발병 원인과 직접 관 계가 있는 동물 모델인 고지방식이로 비만 및 고혈당을 유도 한 마우스에서 세리포리아 락세라타 균사체 배양물이 인슐 린 저항성에 미치는 영향을 연구하며, 근육 세포에서 혈당 강하에 관련된 기전을 규명하고자 하였다.
재료 및 방법
세리포리아 락세라타 균사체 배양물 제조
본 실험에 사용한 균주는 (주)퓨젠셀텍(Sangju, Korea)에 서 보유한 Ceriporia lacerata 균사체를 potato dextrose agar(PDA, Difco Laboratories Inc., Detroit, MI, USA) 배지에 접종하여 25°C에서 9일간 배양하여 사용하였다. C. lacerata 균주를 Shin 등(19)의 방법으로 세리포리아 락세 라타 균사체 배양물을 제조하였다. 전분 4 g/L, 포도당 20 g/L, 정제수 600 mL를 혼합하여 만든 액체 배지에서 pH 5, 온도 23°C, 회전속도 300 rpm으로 10일간 전배양하였 다. 전배양이 종료된 균사체 배양액을 설탕 12.5 g/L, 탈지 대두분 2.5 g/L, 소포제 0.125 g/L, 정제수 400 L를 혼합하 여 만든 액체 배지에 옮겨 pH 5, 온도 23°C, 공기주입량 1.0 kgf/cm2, 회전속도 100 rpm으로 9일간 배양하였다. 배 양한 세리포리아 락세라타 균사체 배양물을 농축 건조 및 분쇄(FBS0P)하여 건조 중량을 기준으로 각 실험군의 용량 에 맞게 사용하였다.
실험동물의 사육
실험동물은 5주령된 수컷 C57BL/6JJmsSlc를 중앙실험 동물(주)(Seoul, Korea)로부터 구입하여 7일간 일반사료 (SAFE+40RMM, SAFE Diets, Augy, France) 급이로 순 화시킨 후 실험에 사용하였으며 물과 사료는 자유롭게 섭취 하도록 하였다. 사육실의 온도는 22±2°C, 상대습도 50±5
% 및 12시간 명암주기로 조절되는 환경에서 사육하였다.
실험군은 일반식이 대조군(CD), 고지방식이 대조군(HFD), 양
성대조군(Met), 실험군(S0P)으로 무작위 배치하였으며, 실 험에 사용된 식이 조성은 Table 1에 나타내었다. HFD, Met, S0P 군은 실험이 진행되는 18주 동안 고지방식이(D12451, Research Diets, Inc., New Brunswick, NJ, USA)를 자유 급이하였고, CD 군은 일반사료로 자유 급이 하였다. CD 군 을 제외한 군은 14주간 고지방식이를 급이한 후 혈당을 측 정하여 평상 시 혈당이 300 mg/dL 이상인 고혈당 마우스를 선별하여 평균 체중이 약 40±5 g이 되도록 각각 10마리씩 각 군에 재배치하였다. 실험 15주차부터 종료 시까지 4주간 매일 동일한 시간에 시료를 phosphate buffer saline(PBS, Welgene Inc., Gyeongsan, Korea)에 녹여 경구 투여하였 다. Met 군은 metformin hydrochloride(Sigma-Aldrich Co., St. Louis, MO, USA)를 150 mg/kg 농도로 투여하였 고, S0P 군은 FBS0P를 500 mg/kg 농도로 투여하였으며, CD 군과 HFD 군은 PBS를 투여하였다. 식이량과 체중은 시료를 투여하는 동안 일주일에 한 번 일정한 시간대에 측정 하였으며, 본 동물실험은 (주)퓨젠바이오파마 동물실험윤리 위원회 승인(승인번호 FBPs-17-001) 하에 진행되었다.
경구 당 부하 검사(oral glucose tolerance test, OGTT) 실험 4주차에 실험동물을 16시간 절식시킨 후 경구 당 부하 검사를 수행하였다. 미정맥에서 ACCU-CHEK Per- forma 혈당기(Roche Ltd., Mannheim, Germany)로 측정한 최초 혈당을 0분 혈당으로 하고, glucose solution을 1 g/kg 으로 경구 투여한 후 각각 15, 30, 60, 90, 120분에 혈당을 측정하였다. 경구 당 부하 검사 결과는 시간에 따른 혈당 그래프 및 area under the curve(AUC)로 나타내었다.
혈액 분석
18주간 사육 후 최종일에 실험동물을 6시간 절식시키고 미정맥에서 혈액을 채취하여 공복혈당을 측정하였다. 공복 혈당 측정 후 Alfaxan과 Rompun을 각각 100 mg/kg, 5 mg/
kg으로 혼합한 마취제를 복강 내 주입으로 마취시켜 심장 채혈 하였다. 채혈된 혈액은 3,000 rpm에서 5분간 원심분리 로 혈장을 분리하여 -70°C에 보관하고 분석 시에 사용하였다.
혈장 인슐린 농도와 c-peptide 농도는 각각 insulin(mouse) ultrasensitive EIA kit(ALPCO, Salem, NH, USA), c-pep- tide(mouse) ELISA kit(ALPCO)을 사용하여 측정하였다.
C2C12 근육세포 배양
C2C12 근육세포를 American Type Culture Collection (ATCC, Manassas, VA, USA)으로부터 구입하였으며, 10%
fetal bovine serum(FBS, Gibco, Grand Island, NY, USA), 1% penicillin/streptomycin(Gibco)을 첨가한 Dulbecco’s modified Eagle’s medium(DMEM, Gibco)으로 37°C, 5%
CO2 환경에서 배양하였다. C2C12가 well의 80% 정도를 차지했을 때 분화 유도를 위해 2% horse serum(Gibco)과 항생제가 첨가된 DMEM으로 6일 동안 배양하였으며, 2일 마다 새 배지로 교환하였다. 현미경으로 관찰하여 세포가 분화된 것을 확인한 후 실험에 사용하였다. 분화된 세포는 음성대조군(NC), 양성대조군(Met), 농도별 실험군(S0P1, S0P10, S0P100)으로 나누어 실험하였다. 실험군은 혈청을 첨가하지 않은 DMEM 배지에 FBS0P를 각각 1, 10, 100 μg/mL의 농도로 처리하고, NC 및 Met 군은 각각 증류수와 metformin 1 mM을 처리하여 24시간 동안 배양하였다. 인 슐린 신호 전달 과정을 자극시키기 위하여 세포를 파쇄하기 30분 전에 인슐린(Sigma-Aldrich Co.) 1 μM을 처리하였다.
인슐린 저항성 유도를 위하여 분화된 세포에 시료 처리와 동시에 TNFα(Sigma-Aldrich Co.) 10 ng/mL를 24시간 동 안 처리하였고, 음성대조군(Veh)은 DMSO로, 양성대조군 (BAY)은 Bay 11-7085(R&D Systems, Inc., Minneapolis, MN, USA) 10 μM로 처리하였다.
Glucose uptake를 측정하기 위하여 glucose uptake as- say kit(Abcam, Eugene, OR, USA)을 사용하였고, micro- plate reader(Synergy H1, BioTek Instruments, Inc., Winooski, VT, USA)로 흡광도 412 nm에서 측정하였다.
Western blot analysis
실험이 종료된 마우스의 종아리 근육 및 근육세포를 이용하 여 인슐린 신호 전달 관련 단백질 및 GLUT4를 측정하였다.
마우스의 종아리 근육을 적출하여 homogenizer(DAIHAN Scientific Co., Ltd., Wonju, Korea)를 이용해 근육을 잘게 균질화하였고, 근육세포는 배지를 제거하여 준비하였다.
PBS로 2~3회 세척한 후 Halt protease and phospha- tase inhibitor cocktail(Thermo, Rockford, IL, USA)을 첨 가한 RIPA buffer(Sigma-Aldrich Co.)를 사용하여 단백질 을 분리하였다.
GLUT4 translocation 여부를 측정하기 위하여 막 단백 질을 분리하여 사용하였다. 균질화시킨 마우스 종아리 근육 및 C2C12 세포에 isolation buffer(250 mM sucrose, 20 mM Tris-HCl pH 7.4, 1 mM EDTA)를 넣어 세포를 파쇄하 고, 원심분리(3,000 rpm, 5분) 하여 핵을 제거하였다. 상등 액을 다시 원심분리(10,300 rpm, 10분) 하여 상등액을 제거 하고 세포막이 포함된 pellet을 isolation buffer로 2~3회 세척하여 단백질 분리를 진행하였다. 분리한 단백질을 BCA protein assay kit(Thermo)을 이용하여 정량하였다. 정량 한 단백질은 10% SDS-PAGE에서 단백질을 분리한 후 gel
을 polyvinylidene fluoride membrane(PVDF, Bio-Rad Laboratories, Hercules, Italy)으로 전이시켰다. START- RM-Temping block T20 TBS buffer(Thermo)로 2시간 blocking 한 다음 일정 비율로 희석한 primary antibody로 4°C에서 overnight로 반응시킨 후 secondary antibody로 실온에서 2시간 반응시켰다. Antibody를 반응시킨 mem- brane에 ECL 기질(Bio-Rad Laboratories)을 반응시킨 후 단백질 밴드는 Chemi Doc(Bio-Rad Laboratories)을 이용 하여 가시화하였다.
본 실험에는 AMPK-α antibody(Cell Signaling Tech- nology Inc., Danver, MA, USA), pAMPK-α antibody(Cell Signaling Technology Inc.), Akt antibody(Cell Signaling Technology Inc.), pAkt(Cell Signaling Technology Inc.), GLUT4 antibody(IF8)(Santa Cruz Biotechnology, Santa Cruz, CA, USA), m-IgGκ BP-HRP(Santa Cruz Biotech- nology), goat anti-rabbit IgG-HRP(Santa Cruz Biotech- nology)를 사용하였다.
통계 처리
본 실험 결과는 평균±표준오차(mean±standard error (SE))로 표기하였으며, 각 그룹 간의 유의성 검증은 Graph Pad Prism 5(Graph Pad Software, San Diego, CA, USA) 를 이용하여 t-test를 실시하였고, 모든 통계적 유의성은 P<0.05에서 검증하였다.
결과 및 고찰
고지방식이 마우스에서 인슐린 저항성 및 혈중 지표에 미 치는 영향
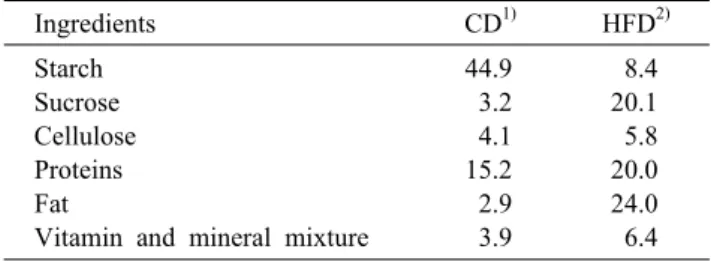
FBS0P를 4주간 투여한 실험동물의 체중 변화, 식이량 및 혈중 지표를(Fig. 1)에 나타내었다. 경구 투여에 의한 FBS0P의 효과를 비교하기 위하여 AMPK activator로 작용 하여 인슐린 저항성을 개선하는 약물인 metformin(20,21) 을 양성대조군으로 투여하였다. 14주간 고지방식이를 급이 하고 CD 군을 제외한 군의 평균 체중이 약 40±5 g이 되도록 재배치하여 0주로 나타내었고, 시료 투여 4주 후 체중은 4주 로 나타내었다(Fig. 1A). 0주차 체중은 CD 군보다 고지방식 이를 급이한 군에서 약 233% 증가하였으며, 시료 투여 4주 후 체중은 모든 실험군에서 약간 증가하였으나 유의적인 차 이는 나타나지 않았다. 식이량(Fig. 1B)은 CD 군보다 고지 방식이를 급이한 군들에서 감소하였는데 이는 Lee 등(22) 의 일반사료를 급이한 군보다 고지방식이를 급이한 군의 식 이량이 유의적으로 감소하였으나 체중 증가량은 현저히 증 가하였다는 보고와 유사하다.
경구 당 부하 검사를 측정한 결과(Fig. 1C), 모든 군에서 포도당 섭취 후 15분 후 혈당이 가장 높았고 시간이 흐를수 록 혈당이 감소하는 것을 확인할 수 있었다. CD 군에서는 초기 혈당 약 98 mg/dL로 시작하여 15분 후 혈당이 약 308
A B
C D
E F G
Fig. 1. Effects of FBS0P on high fat diet-induced diabetic mice with insulin resistance. Mice were divided into four groups (n=10 in each group): Control diet (CD), high fat diet (HFD), high fat diet+metformin at 150 mg/kg/d via oral gavage (Met), and high fat diet+FBS0P at 500 mg/kg/d via oral gavage (S0P). Body weight of the mice during the treatments (A). Average daily food intake of each group was shown as bar graph (B). OGTT was performed 3 weeks after the treatment. Blood glucose level was shown as line graph (C) and area under the curve (AUC) was calculated (D). Blood profile of fasting glucose level (E), serum insulin (F), and serum c-peptide (G). All data represent the mean±SE. *P<0.05 vs CD, #P<0.05 vs HFD.
mg/dL로 상승하였다가 30분 후에 약 214 mg/dL로 감소하 였고, 이후 완만하게 혈당이 감소하다가 120분 후에는 약 133 mg/dL로 측정되었다. 고지방식이를 급이한 군들은 초 기 혈당 및 120분 후 혈당의 수준이 유사하게 측정되었다.
HFD 군에서는 15분 후 혈당이 약 395 mg/dL이고 이후 완 만하게 혈당이 감소하였지만, S0P 군에서는 15분 후 혈당이 약 338 mg/dL에서 30분 후에는 약 234 mg/dL로 급격하게 감소하였다. 이는 HFD 군과 비교하여 유의적인 차이가 있 었다. AUC(Fig. 1D)를 계산한 결과 CD 군보다 HFD 군에서 144.23% 유의적으로 높았고, S0P 군에서 HFD 군보다 25.99% 유의적으로 감소하였다. 실험 종료 후 시료 투여에 따른 공복혈당 측정 결과(Fig. 1E), 일반 사료를 급이한 CD 군에서 혈당이 약 93.37 mg/dL로 측정되었고, 고지방식이 를 한 HFD 군에서는 157.60 mg/dL로 고혈당으로 나타났 다. FBS0P를 투여한 군에서의 혈당은 약 113.80 mg/dL로 Met 군의 114.70 mg/dL와 유사한 정도로 혈당이 유의적으 로 감소하였다. 이 결과를 통해 고지방식이 군에서 인슐린 저항성이 증가한 반면, 추가로 FBS0P를 투여한 군에서는
인슐린 저항성이 감소한 것을 알 수 있었다. 이 결과가 혈액 프로파일에서도 재현되는지 확인하기 위하여 혈장 인슐린 농도를 측정하였다.
혈장 인슐린 농도(Fig. 1F)는 HFD 군에서 약 1.82 ng/mL 로 CD 군보다 약 210.82% 유의적으로 증가하였고, Met 군과 S0P 군에서는 각각 약 1.07 ng/dL, 1.21 ng/dL로 정상 동물과 유사한 정도로 유의하게 감소하였다. 혈장 c-pep- tide 농도(Fig. 1G)는 HFD 군에서 약 781.08 pM로 CD 군 보다 177% 정도 증가하였고, S0P 군에서는 687.08 pM로 HFD 군보다 유의적으로 감소하였다.
고지방식이를 하게 되면 비만으로 인한 인슐린 저항성이 초래되는데(23), 4주간 FBS0P 섭취 후 혈장 인슐린 농도가 유의적으로 감소한 것을 통하여 인슐린 저항성이 개선된 것 을 유추할 수 있었다.
근육 내 GLUT4 발현에 미치는 영향
AMPK의 인산화로 인하여 혈중 포도당이 세포 내로 유입 되는 통로인 GLUT4가 세포질에서 세포막으로 이동하게 되
A B
C D
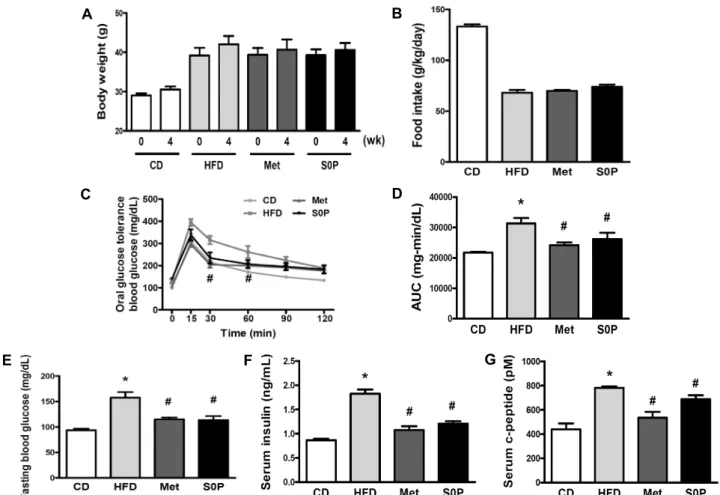
Fig. 2. Effects of FBS0P on AMPK phosphorylation and GLUT4 expression in gastrocnemius muscle of high fat diet induced-diabetic mice. Gastrocnemius muscle tissues were lysed and subjected into western blot analysis (A). The band intensity of pAMPK was normalized to that of AMPK (B). The mean value obtained with tissue lysates from CD was set to 1. Gastrocnemius muscle tissues were divided into whole tissue lysate and plasma membrane fraction. Proteins were subjected to western blot analysis with specific antibodies (C). The band intensity of GLUT4 was normalized to GAPDH (whole tissue lysate) and Caveolin (plasma membrane) (D). The mean value obtained with tissue lysates from CD was set to 1. All data represent the mean±SE. N.S.: not significant,
#P<0.05 vs HFD.
는데, 고지방식이 마우스 모델에서 FBS0P 섭취로 인해 공 복혈당이 감소하고 인슐린 저항성이 개선되는 것이 AMPK 인산화 및 GLUT4 translocation에 의한 것인지 연구하고 자 하였다.
AMPK 인산화 정도를 western blot으로 확인한 결과 (Fig. 2A, 2B), CD 군의 pAMPK/AMPK ratio를 1로 보았을 때 HFD 군에서 AMPK 인산화가 약 32.24% 감소하였는데, 이는 Kuo 등(24)이 고지방식이를 급이한 쥐의 골격근 조직 에서 정상식이를 급이한 쥐보다 pAMPK 단백질 발현량이 감소하였다는 보고와 유사하다. S0P 군에서는 유의적이지 는 않으나 HFD 군에 비하여 약 193.44% 증가하였다.
조직 전체와 세포막을 분리하여 얻은 단백질에서 GLUT4 발현량을 western blot으로 측정한 결과(Fig. 2C, 2D)를 확인하였다. 조직 내 전체 GLUT4 및 세포막 GLUT4 발현 량에서 각각의 CD 군을 기준으로 비교한 결과, HFD 군의 조직 전체 및 세포막에서 GLUT4 발현량이 유의적이지는 않으나 소폭 감소하였고, Met군과 S0P 군에서는 유의적으 로 증가하였다. 이는 Ji 등(25)의 AMPK 인산화가 GLUT4 translocation을 유도하여 혈당 조절에 도움을 줄 수 있다는 보고와 유사하다. 실제로 FBS0P 투여로 인해 마우스에서 당 대사가 효과적으로 이루어진 것은 세포막 GLUT4 발현 증가에 의한 것으로 유추할 수 있다. 동물실험에서의 결과가 세포 수준에서도 확인 가능한지 마우스 골격근 세포에서 in vitro 실험을 진행하였다.
근육세포에서 당 흡수에 미치는 영향
FBS0P가 근육세포에서 AMPK 인산화 정도를 확인한 결 과(Fig. 3A, 3B), Met 군에서의 AMPK 인산화가 약 337.61
% 증가하였다. FBS0P를 10 μg/mL 처리하였을 때 149.69
% 정도 증가하였고, 100 μg/mL 처리하였을 때 301.06%로 유의적으로 증가하였다. 이 결과로 FBS0P가 유의적이지는 않으나 농도 의존적으로 AMPK의 인산화에 효과가 있는 것 을 확인할 수 있었으며, 이는 metformin이 AMPK를 인산화 시키는 것과 유사한 정도임을 확인할 수 있었다.
세포 내 전체 GLUT4 및 세포막 GLUT4 발현량을 비교한 결과(Fig. 3C) 각각 NC 군에 비하여 Met 군의 GLUT4 발현 량이 증가하였고, S0P 군에서는 농도 의존적으로 GLUT4 발현량이 증가한 것을 확인하였다. 이는 마우스 근육조직에 서 발현이 증가한 것과 유사한 결과로 확인할 수 있었다.
더 나아가 근육세포에서 포도당 흡수를 확인한 결과(Fig.
3D), metformin, FBS0P 100 μg/mL를 각각 처리하였을 때 시료를 처리하지 않은 NC 군에 비하여 각각 237.09%, 202.66% 정도로 증가하였다. 추가로 인슐린을 처리한 후 인슐린을 처리하지 않은 군의 결과와 비교하였을 때, NC 군은 약 219.70%, Met, S0P100 군에서는 각각 134.36%, 170.37%로 당 흡수가 증가하였다. 인슐린 처리 없이도 시 료에 의하여 당 흡수가 증가하였고, 체내 반응과 유사 조건 으로 인슐린을 처리하였을 때 당 흡수가 더 효과적으로 증가 한다는 것을 알 수 있었다. 그러므로 FBS0P 투여가 마우스
A B
C D
Fig. 3. Effects of FBS0P on glucose uptake in C2C12 myotubes. C2C12 myotubes were treated with vehicle (NC), metformin (1 mM), and FBS0P (1 to 100 μg/mL) for 24 h. Cell lysates were subjected to western blot analysis with specific antibodies (A). The band intensity of pAMPK was normalized to that of AMPK (B). The mean value obtained with cell lysates from NC was set to 1. C2C12 myotubes were divided into whole cell lysate and plasma membrane fraction. Proteins were subjected to western blot analysis with specific antibodies (C). After insulin (Ins) treatment for 30 min, glucose uptake was measured (D).
All data represent the mean±SE. *P<0.05 vs NC, #P<0.05 vs S0P100 without insulin treatment.
A B
C D
Fig. 4. Effects of FBS0P on insulin signal pathway in C2C12 myotubes. C2C12 myotubes were treated with vehicle (NC), metformin (1 mM), and FBS0P (10 and 100 μg/mL) for 24 h. To stimulate insulin signal pathway, insulin (1 μM) was treated for 30 min.
Cell lysates were subjected to western blot analysis with specific antibodies (A). The band intensity of pAkt was normalized to that of Akt (B). The mean value obtained with tissue lysates from NC was set to 1. TNFα was treated to mimic insulin resistance status. Also, cell lysates were subjected to western blot analysis with specific antibodies (C). The band intensity of pAkt was normalized to that of Akt (D). The mean value obtained with tissue lysates from NC was set to 1. All data represent the mean±SE.
N.S.: not significant.
의 혈당 강하에 직접 도움을 줄 수 있음을 유추할 수 있었다.
근육세포에서 인슐린 신호 전달 과정에 미치는 영향 AMPK 인산화가 인슐린 신호 전달 과정에 영향을 미친다 는 연구 결과(26)를 바탕으로 근육세포에 FBS0P와 인슐린 을 처리하여 Akt의 인산화 정도를 측정하였다(Fig. 4A, 4B).
Akt의 인산화는 인슐린 처리에 의해서만 증가하며, 모든 실 험군에서 Akt의 인산화에 유의적인 차이를 나타내지 않았 다. 따라서 본 연구에 사용한 동물 모델과 유사한 인슐린 저항성 상태를 유도하기 위하여 근육세포에 TNFα를 처리 한 후 Akt 인산화 정도를 측정하였다(Fig. 4C, 4D). TNFα 를 처리하고 시료를 처리하지 않은 Veh 군에서는 인슐린을 처리했음에도 불구하고 Akt의 인산화가 전혀 일어나지 않 았다. TNFα 자극을 억제시키는 Bay 군(NF-κB 저해제)에 서는 132.51%, S0P100 군에서는 142.62% 정도 증가한 것으로 보아 고지방식이 동물모델에서 FBS0P가 인슐린 저 항성을 개선시켰을 것으로 유추할 수 있었다.
요 약
본 연구에서는 세리포리아 락세라타 균사체 배양물(FBS0P) 이 고지방식이를 급이한 마우스에서 인슐린 저항성에 미치 는 영향을 규명하고자 C57BL/6J 마우스에 14주간 고지방 식이를 급이하고 4주간 매일 FBS0P를 500 mg/kg으로 경 구 투여하여 혈액 및 근육조직에서의 단백질 발현을 분석하 였다. FBS0P 군의 식이량과 체중에서 HFD 군과 유의한 차이가 나지 않았으나 OGTT 및 공복혈당을 측정한 결과 HFD 군에 비하여 당 대사가 유의하게 개선된 것을 확인할 수 있었으며, 혈장 인슐린 농도와 c-peptide 농도가 유의하 게 감소하여 인슐린 저항성이 개선된 것을 유추할 수 있었 다. FBS0P가 근육 내 당 수송에 미치는 영향을 확인하기 위하여 마우스 종아리 근육에서 AMPK 인산화 및 GLUT4 발현 정도를 확인한 결과, FBS0P 투여에 의하여 AMPK의 인산화 및 세포막에서의 GLUT4 발현이 증가하였음을 확인 하였다. 동물실험에서의 결과를 뒷받침하기 위해 마우스 골 격근 세포에서 AMPK 인산화 및 GLTU4 발현 정도를 확인 한 결과, 동물실험에서와 마찬가지로 FBS0P가 AMPK의 인 산화와 세포막에서의 GLUT4를 유의적으로 증가시켰으며 농도 의존적으로 작용함을 확인할 수 있었다. 세포에서 포도 당 흡수를 측정한 결과 인슐린 처리 없이도 시료에 의하여 당 흡수가 증가하였고, 체내 반응과 유사하게 인슐린을 처리 하였을 때에 당 흡수가 더 효과적으로 증가한다는 것을 알 수 있었다. 또한, 인슐린에 의해 활성화되는 Akt의 인산화 정도를 확인하였을 때 TNFα로 인슐린 저항성을 유도한 세 포에서 FBS0P가 Akt 인산화를 증가시켰다. 이를 통해 고지 방식이 동물모델에서 FBS0P가 인슐린 저항성을 개선시켰 을 것으로 유추할 수 있었다. 따라서 본 연구를 통하여 FBS0P가 인슐린 저항성이 유발된 제2형 당뇨병 개선에 많
은 도움이 될 것으로 기대된다.
REFERENCES
1. Choi CS, Park SW. 2002. The mechanism of development of insulin resistance in type 2 diabetes mellitus. Korean J Med 63: 613-624.
2. Kim YH, Yoon JH, Kim KC. 2013. Effects of exercise in- tensity on AMPK, GLUT-4, MCT-1 expression and insulin sensitivity in skeletal muscle of type 2 diabetic mice. Kore- an J Physical Education 52: 599-612.
3. Hayashi T, Wojtaszewski JF, Goodyear LJ. 1997. Exercise regulation of glucose transport in skeletal muscle. Am J Physiol 273: E1039-E1051.
4. Thirone AC, Huang C, Klip A. 2006. Tissue-specific roles of IRS proteins in insulin signaling and glucose transport.
Trends Endocrinol Metab 17: 72-78.
5. White MF. 2003. Insulin signaling in health and disease.
Science 302: 1710-1711.
6. Ishikura S, Bilan PJ, Klip A. 2007. Rabs 8A and 14 are tar- gets of the insulin-regulated Rab-GAP AS160 regulating GLUT4 traffic in muscle cells. Biochem Biophys Res Com- mun 353: 1074-1079.
7. Stoll BA. 1999. Western nutrition and the insulin resistance syndrome: a link to breast cancer. Eur J Clin Nutr 53: 83-87.
8. Galgani JE, Moro C, Ravussin E. 2008. Metabolic flexibility and insulin resistance. Am J Physiol Endocrinol Metab 295:
E1009-E1017.
9. Shepherd PR, Kahn BB. 1999. Glucose transporters and in- sulin action: Implications for insulin resistance and diabetes mellitus. N Engl J Med 341: 248-257.
10. Dolinsky VW, Dyck JR. 2006. Role of AMP-activated pro- tein kinase in healthy and diseased hearts. Am J Physiol Heart Circ Physiol 291: H2557-H2569.
11. Hardie DG. 2007. AMP-activated/SNF1 protein kinases: con- served guardians of cellular energy. Nat Rev Mol Cell Biol 8: 774-785.
12. Suhara H, Maekawa N, Kaneko S, Hattori T, Sakai K, Kondo R. 2003. A new species, Ceriporia lacerata, isolated from white-rotted wood. Mycotaxon 86: 335-347.
13. Lee JW, Gwak KS, Park JY, Park MJ, Choi DH, Kwon M, Choi IG. 2007. Biological pretreatment of softwood Pinus densiflora by three white rot fungi. J Microbiol 45: 485-491.
14. Shan WG, Liang DE, Ying YM, Zhan ZJ. 2012. Two new tremulane sesquiterpenoids from Ceriporia lacerate, an en- dophytic fungus of Huperzia serrata. J Chem Res 36: 365- 366.
15. Ying YM, Shan WG, Zhang LW, Chen Y, Zhan ZJ. 2013.
Lanostane triterpenes from Ceriporia lacerate HS-ZJUT- C13A, a fungal endophyte of Huperzia serrata. Helv Chim Acta 96: 2092-2097.
16. Wang J, Yao L, Lu Y. 2013. Ceriporia lacerata DMC1106, a new endophytic fungus: Isolation, identification, and opti- mal medium for 2’,4’-dihydroxy-6’-methoxy-3’,5’-dimeth- ylchalcone production. Biotechnol Bioprocess Eng 18: 669- 678.
17. Kim JE, Kim HJ, Lee SP. 2012. Hyperglycemic effect of submerged culture extract of Ceriporia lacerata in strepto- zotocin-induced diabetic rats. Food Sci Biotechnol 21: 1685- 1693.
18. Kim JH, Park YK, Kim JE, Lee SP, Kim BC, Jang BC.
2013. Crude extract of Ceriporia lacerata has a protective effect on dexamethasone-induced cytotoxicity in INS-1 cells
via the modulation of PI3K/PKB activity. Int J Mol Med 32: 179-186.
19. Shin EJ, Kim JE, Kim JH, Park YM, Yoon SK, Jang BC, Lee SP, Kim BC. 2015. Effect of submerged culture of Ceri- poria lacerata mycelium on GLUT4 protein in db/db mouse.
Korean J Food Preserv 22: 893-900.
20. Zhou G, Myers R, Li Y, Chen Y, Shen X, Fenyk-Melody J, Wu M, Ventre J, Doebber T, Fujii N, Musi N, Hirshman MF, Goodyear LJ, Moller DE. 2001. Role of AMP-activated protein kinase in mechanism of metformin action. J Clin Invest 108: 1167-1174.
21. Musi N, Hirshman MF, Nygren J, Svanfeldt M, Bavenholm P, Rooyackers O, Zhou G, Wiliamson JM, Ljunqvist O, Efendic S, Moller DE, Thorell A, Goodyear LJ. 2002. Met- formin increases AMP-activated protein kinase activity in skeletal muscle of subjects with type 2 diabetes. Diabetes 51: 2074-2081.
22. Lee JS, Lee MK, Ha TY, Bok SH, Park HM, Jeong KS, Woo MN, Do GM, Yeo JY, Choi MS. 2006. Supplementa- tion of whole persimmon leaf improves lipid profiles and
suppresses body weight gain in rats fed high-fat diet. Food Chem Toxicol 44: 1875-1883.
23. Kaushik G, Satya S, Khandelwal RK, Naik SN. 2010. Com- monly consumed Indian plant food materials in the manage- ment of diabetes mellitus. Diabetes Metab Syndr 4: 21-40.
24. Kuo YH, Lin CH, Shih CC. 2015. Ergostatrien-3β-ol from Antrodia camphorata inhibits diabetes and hyperlipidemia in high-fat-diet treated mice via regulation of hepatic related genes, glucose transporter 4, and AMP-activated protein kin- ase phosphorylation. J Agric Food Chem 63: 2479-2489.
25. Ji L, Zhang X, Liu W, Huang Q, Yang W, Fu F, Ma H, Su H, Wang H, Wang J, Zhang H, Gao F. 2013. AMPK- regulated and Akt-dependent enhancement of glucose up- take is essential in ischemic preconditioning-alleviated re- perfusion injury. PLoS One 8: e69910.
26. Chopra I, Li HF, Wang H, Webster KA. 2012. Phosphoryla- tion of the insulin receptor by AMP-activated protein kinase (AMPK) promotes ligand-independent activation of the in- sulin signalling pathway in rodent muscle. Diabetologia 55:
783-794.