가자, 암라 추출복합물(FBF-TC)의 분화된 3T3-L1 세포와 고지방식이로 유도된 비만동물모델에 대한 항비만 효과
박수정․이민희․윤정문․김다경․임현정 경희대학교 임상영양연구소
Anti-Obesity Effects of Combined Extracts of Terminalia chebula and Phyllanthus emblica (FBF-TC) in Matured 3T3-L1 Adipocytes
and High Fat Diet-Induced Obese Mice
Soo-Jeung Park, Minhee Lee, Jeong-Moon Yun, Dakyung Kim, and Hyunjung Lim Research Institute of Medical Nutrition, Kyung Hee University
ABSTRACT Abnormal fat accumulation on the body causes chronic disease known as obesity and is a serious problem in today’s society. We investigated the anti-obesity effects of combined extracts of Terminalia chebula (T) and Phylan- thus emblica (P) (TP; FBF-TC) in vitro and in vivo. 3T3-L1 preadipocytes were differentiated for 9 days. On the last day of differentiation, Oil red O staining, glycerol release, intracellular cAMP levels, and lipolytic enzymes were measured. Compared to the control, lipid accumulation, and mRNA expressions of PDE3B and Perilipin were sig- nificantly decreased in the presence of TP 80 μg/mL. The levels of free glycerol, intracellular cAMP, and mRNA expressions of HSL and ATGL were significantly increased by 139%, 99.5%, 65.6%, and 100.8%, respectively, in the presence of TP 80 μg/mL. In the animal study, C57BL/6J mice were tested for 12 weeks after being separated into eight groups: Normal diet group (ND), high fat diet (HFD) group (C, Control), HFD with 1 g/kg b.w. of catechin group (PC, positive control), HFD with 600 mg/kg b.w. of T or P groups, and HFD with 150, 300, 600 mg/kg b.w.
of TP groups. Feeding of HFD for 12 weeks resulted in significant increases in body weight compared to the ND group. FBF-TC groups showed reduced body weight, weight gain, mesenteric white adipose tissue (WAT) weight, subcutaneous WAT weight, serum total cholesterol, LDL-cholesterol, and triglyceride levels compared to the C group.
In conclusion, this study suggests that TP is a novel anti-obesity candidate through its stimulation of lipolysis.
Key words: Terminalia chebula, Phyllanthus emblica, anti-obesity, 3T3-L1, C57BL/6J
Received 26 October 2018; Accepted 13 December 2018 Corresponding author: Hyunjung Lim, Research Institute of Medi- cal Nutrition, Kyung Hee University, Seoul 02447, Korea E-mail: [email protected], Phone: +82-31-201-2343
Author information: Soo-Jeung Park (Researcher), Minhee Lee (Re- searcher), Jeong-Moon Yun (Researcher), Dakyung Kim (Researcher), Hyunjung Lim (Professor)
서 론
지방세포(adipocyte)는 체내의 지질 및 에너지 저장을 위 해 특화된 세포이며 특정 호르몬을 분비하여 여러 가지 대사 작용에 관여한다(1). 지방세포의 수 증가와 비대는 과도한 지방축적으로 이어져 비만, 제2형 당뇨병, 고혈압 및 관상동 맥성 심장병 등과 같은 비만성 대사질환을 일으킨다(2). 따 라서 최근 세포 내의 지방축적을 억제하거나 축적된 지방의 분해 효능이 있는 다양한 천연물 소재에 대한 연구들이 진행 되고 있다.
체내의 지방축적은 지방세포 내에 존재하는 지방구에서
중성지방(triglyceride; TG)의 합성과 분해를 통하여 조절된 다(3). 지방산과 글리세롤로 이루어진 TG의 분해는 호르몬, 신경전달물질 및 여러 전사인자 등에 의해 조절되는데(4,5), 특히 cyclic AMP-dependent protein kinase A(PKA)를 촉진하는 cAMP 경로가 주요기전으로 알려져 있다. 지방구 의 세포질에 존재하는 hormone sensitive lipase(HSL)는 인슐린(insulin)이나 에피네프린(epinephrine) 등의 호르몬 에 의해 활성화된 PKA에 의해 인산화되어 지방구 표면 위로 전위되고 Perilipin A와 상호 작용하여 유리지방산(free fat- ty acid) 및 글리세롤(glycerol)의 방출을 유도한다(5-7).
Adipose triglyceride lipase(ATGL) 또한 지방조직에서 주 로 발현되며, 대표적 전지방세포(preadipocyte)인 3T3-L1 세포에서 속도제한 지방분해효소로 보고된 바 있다(8,9).
3T3-L1 세포는 분화유도물질 처리 시 지방세포로 변화하 는데 지질대사 조절, 에너지 항상성 유지 및 항비만 활성에 있어서 적용 가능한 모델로 널리 알려져 있다(10).
가자(Terminalia chebula)는 하리타키(Haritaki)라고도 하며 사군자과에 속한 낙엽교목으로, 성숙한 열매의 성상은
긴 원형이고 표면은 황갈색을 띤다. 주로 인도, 대만 등 동남 아시아에서 널리 번식하고 있으며(11), tannins, ellagic acid, punicalagin, flavonoids, gallic acid, chebulinic acid, chebulagic acid 등의 성분들이 함유되어 있다(12). 최근 항암 활성, 항당뇨 활성, 항돌연변이 활성, 항균 활성 등 다양 한 생리활성이 보고된 바 있고(13-16), 특히 비만쥐에서 체 중증가량, 내장지방, 혈중 중성지방 농도 및 콜레스테롤 수 치가 감소하였다는 보고가 있다(17). 암라(Phyllanthus emblica)는 인도뿐만 아니라 미국, 영국, 중국 등 많은 나라 에서 식품이나 건강보조식품의 형태로 사용되고 있고, 비타 민 C를 비롯하여 phenolic compound, tannins, phyllem- belic acid, phyllembelin, rutin, curcuminoids, emblicol 등의 성분들이 함유되어 있는데(18-20), 최근 심혈관계질 환, 고지혈증 등에도 효과가 있으며, 특히 비만쥐에서 내장 지방 개선에 효과가 나타난 것으로 보고된 바 있다(17).
이처럼 가자와 암라의 생리활성에 대해서는 알려진 바가 많고, 특히 체지방감소에도 효능이 있는 것으로 알려져 있으 나, 아직 복합추출물의 항비만 효과에 대해서는 보고된 바가 없다. 또한, 체지방감소 기능성 식품 소재로서의 요구도가 증대되고 있으나, 수입에 의존해야 하는 원료 특성상 단가들 의 경제적인 면에서 제한점이 대두되고 있다. 이에 가자와 암라 복합물에 대한 연구를 통해 효능이 있는 최소 필요량을 파악하고, 그로 인한 경제적인 절감 가능성 파악이 필요하 다. 따라서 본 연구에서는 가자와 암라 복합추출물을 분화된 3T3-L1 지방세포와 고지방식이로 유도된 비만동물모델에 적용하여 항비만 효과에 미치는 영향을 조사하였다.
재료 및 방법
시료의 준비
본 실험에 사용된 가자 열수추출물과 암라 열수추출물은 (주)프롬바이오(Suwon, Korea)로부터 공급받았다. 가자 열 수추출물의 경우 건조분말을 추출, 여과, 분무건조의 과정을 거쳤고, 암라 열수추출물의 경우 분쇄된 시료를 추출, 여과, 농축, 진공건조 과정을 거쳤으며, 수득율은 각각 30%, 20%
였다. 가자추출물과 암라추출물은 기능성 및 안전성을 고려 하고 경제적인 절감을 위한 최소한의 원료 단가를 고려하여 7:3의 혼합비율로 하여 가자, 암라 추출복합물인 FBF-TC (TP)를 제조하여 사용하였다.
재료 및 시약
본 실험에서 사용된 3T3-L1(CL-173) preadipocyte는 Americal Type Cultured Collection(ATCC, Manassas, VA, USA)에서 분양받아 실험하였다. High-glucose Dul- beco’s modified Eagle’s medium(DMEM), phosphate- buffered saline(PBS), fetal bovine serum(FBS), new- born calf serum(NCS), L-glutamine, penicillin-strep- tomycin, HEPES, sodium pyruvate 및 NEAA mixture는
HyClone(Waltham, MA, USA)으로부터 구입하였다. Di- methyl sulfoxide(DMSO), tetrazolium bromide(MTT), 3- isobutyl-1-methylxanthine(IBMX), dexamethasone(DEX), insulin, Oil red O 및 free glycerol reagent는 Sigma- Aldrich Co.(St. Louis, MO, USA)로부터 구입하였고, cAMP ELISA kit은 Cell Biolabs Inc.(San Diego, CA, USA)로부터 구입하여 사용하였으며, Green tea extract catechin은 (주)네이처스토리(Seoul, Korea)에서 구입하여 사용하였다.
3T3-L1 세포배양 및 분화유도
3T3-L1 세포는 10% NCS, 1% L-glutamine, 1% pen- icillin-streptomycin, 1% HEPES, 1% sodium pyruvate, 1% NEAA mixture를 함유한 DMEM을 이용하여 37°C, 5%
CO2의 조건에서 배양하였다(21). 2일마다 배지를 교환하였 고, 세포가 플라스크 바닥에 80% 이상 부착되면 계대배양을 하였다. 이들 세포를 6-well plate에 1×105 cells/well로 균등히 분주하였고, 100% confluent 한 상태인 4일 후부터 기존 배지의 10% NCS를 10% FBS로 교체하였으며, MDI solution(adipogenic cocktail)인 IBMX(0.5 mM), DEX(1 μM), insulin(10 μM)을 첨가하여 분화유도를 시작하였다.
분화 기간은 총 9일간 지속하였으며, 분화 초기 3일간은 MDI solution이 함유된 배지를, 분화 중기 3일간은 insulin (10 μM)만을 함유한 배지를, 분화 후기 3일간은 10% FBS 만 함유하는 배지로 매일 교환해주었다.
세포독성
3T3-L1 세포에 대한 TP의 독성을 평가하기 위해 MTT assay(22)를 수행하였다. MTT solution(5 mg/mL)은 PBS 에 완전히 녹인 후 filtering 하여 사용하였다. 3T3-L1 세포 를 96-well plate에 1×104 cells/well(100 μL)로 분주하였 으며, 배양액은 10% FBS를 함유한 DMEM을 사용하였다.
12시간 동안 배양한 후 TP를 농도별(0, 10, 20, 30, 40, 50, 60, 70, 80, 90 및 100 μg/mL)로 처리하였다. 72시간 후 MTT solution 20 μL를 분주하여 MTT 환원에 있어서 빛의 요인을 최소화하기 위하여 빛을 차단하였고 4시간 동 안 반응시켰다. 각 well의 배양액과 MTT solution을 제거 하고 PBS로 2번 세척한 후, DMSO 200 μL를 분주하여 in- cubator에서 녹였다. 20분 후 iMARKTM Microplate Read- er(Bio-Rad, Hercules, CA, USA)로 560 nm 파장에서 optical density를 측정하였다.
Oil red O 염색
분화된 3T3-L1 세포에 세포독성이 나타나지 않았던 TP 의 농도 범위(<80 μg/mL) 내에서 TP(0, 10, 20, 40, 80 μg/
mL)를 처리하고 72시간 후에 세포의 배지를 제거한 후 PBS 로 2회 세척하였으며, 10% formalin을 분주하여 실온에서 고정했다. 5분 후 10% formalin을 제거한 후 새로운 10%
Table 1. Primer sequences used in real-time PCR quantification of mRNA
Gene Primer sequences PDE3B
PKA Perilipin
HSL ATGL GAPDH
F 5’-AGTGAAAAGGATCGCCTCTTG-3’
R 5’-TTGCTGGGCCATTGATATCTG-3’
F 5’-TTCACTCAGAGCCGCTTAAGG-3’
R 5’-CTCTATCCGAAGTATTGCTGCTACCT-3’
F 5’-GCTCTTCAATACCCTCCAGAAGGA-3’
R 5’-TTCGAAGGCGGGTAGAGATG-3’
F 5’-CACTAGTCCCTCCCCCAGTTT-3’
R 5’-AGCTGGCACAGCAGGTCTGT-3’
F 5’-CATTCTCAGGCGAGAGTGACAT-3’
R 5’-GACGCGAAGCTCGTGGAT-3’
F 5’-CATGGCCTTCCGTGTTCCTA-3’
R 5’-GCGGCACGTCAGATCCA-3’
PDE3B, phosphodiesterase 3B; PKA, cyclic AMP-dependent pro- tein kinase A; HSL, hormone sensitive lipase; ATGL, adipose triglyceride lipase; GAPDH, glyceraldehyde-3-phosphate dehy- drogenase.
formalin을 분주하여 실온에서 고정했다. 2시간 후 10% for- malin을 제거하고 60% isopropanol을 분주하여 곧바로 제 거한 후 완전히 건조하게 했다. 그 후 Oil red O solution 시약을 분주하여 지방구를 염색하였다. 1시간 후 증류수로 4번 세척하였고, 붉은색으로 염색된 세포 내 지방구를 현미 경으로 관찰하였다. 지방의 축적량을 정량하기 위해 증류수 로 세척되었던 well마다 100% isopropanol을 분주하여 Oil red O dye를 용출시킨 후 iMARKTM Microplate Reader (Bio-Rad)로 560 nm 파장에서 optical density를 측정하였 다(21).
TG 함량의 측정
세포 내 TG 함량은 Triglyceride Quantification Col- orimetric Kit(BioVision, Milpitas, CA, USA)을 이용하여 측정하였다(23). TP가 처리된 세포를 5% NP-40으로 균질 화시켜 80~100°C에서 천천히 가열시킨 후 실온에서 냉각 시켰고, TG의 안정화를 위해 1회 더 반복하였다. 2분간 원 심 분리하여 얻어진 상층액과 TG standard를 96-well plate에 분주한 후 TG assay buffer를 정해진 용량대로 분 주하였다. 각 well에 lipase 2 μL씩 분주하여 잘 섞은 후 20분간 방치하였고, TG reaction mix를 각 well에 50 μL씩 분주하였다. 60분 후 iMARKTM Microplate Reader(Bio- Rad)로 570 nm 파장에서 optical density를 측정하였다.
Free glycerol의 측정
분화된 3T3-L1 세포에서 TP 처리가 지방분해산물인 free glycerol 방출량에 미치는 영향을 알아보고자 glycer- ol phosphate oxidase(GPO)-TRINDER 효소반응법(24) 을 적용하였고, free glycerol reagent 시약을 이용하여 9일 동안 분화된 3T3-L1 세포의 배지 내 glycerol 함량을 측정 하였다. Free glycerol reagent 800 μL와 배지 1 mL를 혼 합하여 37°C에서 10분간 반응시켰고 iMARKTM Micro- plate Reader(Bio-Rad)로 560 nm 파장에서 optical den- sity를 측정하였다. Free glycerol 표준시약을 사용하여 표 준곡선을 활용하여 glycerol의 함량을 산출하였다.
세포 내 cAMP level 측정
세포 내 cAMP의 농도는 cAMP ELISA Kit(Cell Biolabs Inc.)을 이용하여 측정하였다(25). TP가 처리된 세포에 Lysis buffer 1 mL를 분주하여 4°C에서 20분간 방치시킨 후 cell scraper로 세포를 모았으며, micropipet을 사용하여 균질 화시켰다. 10분간 최고속도에서 원심 분리한 상층액과 cAMP standard 25 μL를 Goat Anti-Rabbit Antibody Coated Plate의 각 well에 분주하였고, Peroxidase cAMP Tracer Conjugate 25 μL를 분주하였다. Rabbit Anti-cAMP Poly- clonal Antibody 50 μL를 분주한 후 plate 윗부분을 잘 막아 주었다. 2시간 동안 실온에서 방치시킨 후 1× wash buffer 로 5회 세척해주었고, substrate solution 100 μL를 각 well
에 분주한 후 실온에 방치시켰다. 20분 후 stop solution 100 μL를 분주하여 iMARKTM Microplate Reader(Bio-Rad)로 450 nm 파장에서 optical density를 측정하였다.
RNA 추출 및 real-time PCR
분화된 3T3-L1 세포에서 TP 처리가 지방 대사에 관여하 는 효소들의 발현 정도에 미치는 영향을 파악하기 위해 re- al-time PCR을 수행하였다. 분화된 3T3-L1 세포에 TP를 처리하고 72시간 후에 세포를 거두었다. 지방세포로부터 총 RNA 추출은 RNeasy Mini Kit(QIAGEN Sciences, Ger- mantown, MD, USA)을 이용하여 제조사에서 제공하는 방 법에 따라 추출하였다. cDNA 합성은 iScript cDNA Syn- thesis Kit(Bio-Rad)을 이용하였고, SYBR Green(iQ SYBR Green Supermix, Bio-Rad)과 Real-Time PCR(Applied Biosystems, Foster City, CA, USA) 기기를 이용하여 유전 자들의 발현을 측정하였다. 각 유전자에 대한 PCR primer 염기서열은 Table 1에 나타내었다. Real-time PCR 수행 시 2 μL의 cDNA, 10 μL의 2× SYBR mix, 1 μL의 forward primer(100 pmol/μL) 및 1 μL의 reverse primer(100 pmol/μL)를 혼합하여 나머지는 nuclease free water로 채 워 총 20 μL가 되게 하였다. PCR 증폭은 총 40 cycle을 실시하였고, hot start를 위해 95°C에서 10분, 증폭 단계는 denaturation을 95°C에서 15초, annealing을 54°C에서 30 초, extension을 72°C에서 30초를 반복하였다. 모든 cycle 이 완료된 후 melting curve 분석을 하여 primer들의 특이 성을 확인하였다. 결과의 분석은 Applied Biosystems에서 제공하는 프로그램(One step system software v2.1)으로 분석하였다.
실험동물 및 식이
실험동물은 C57BL/6J mice(5 wks, male)를 Charles River Laboratories Inc.(Yokohama, Japan)로부터 분양받
*
*
0 20 40 60 80 100 120
0 10 20 30 40 50 60 70 80 90 100 Concentration (μg/mL)
Cell viability (% of control) .
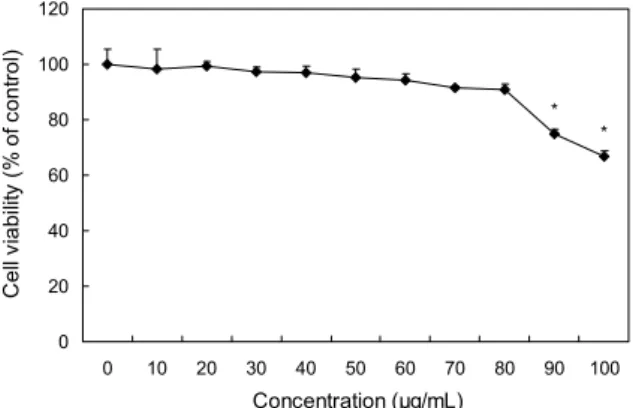
Fig. 1. Effect of combined extract of Terminalia chebula and Phyllanthus emblica (TP) on cell viability in 3T3-L1 preadipo- cytes. The cells were incubated with various concentrations of TP for 72 hours. Cell viability was determined by MTT assay.
Data were calculated as a percent growth rate of cells in pres- ence of TP, compared with untreated control cells. Values are mean±SD (n=3). *Significantly different at P<0.05 compared with 0 μg/mL by the t-test, for each time point.
아 항온항습(22±3°C, 55±10%), 12 h/12 h light/dark cy- cles인 조건으로 사육하였고, 경희대학교 동물실험윤리위 원회의 승인을 받아 수행하였다(승인번호: KHUASP(SE)- 16-013). 동물실험은 1주일간 순화 사육기간을 거친 후 normal diet(ND, AIN93G), high fat diet(HFD)(C, 60%
HFD), positive control(PC, 60% HFD+catechin 1 g/kg b.w.), T600(60% HFD+T 600 mg/kg b.w.), P600(60%
HFD+P 600 mg/kg b.w.), TP150(60% HFD+TP 150 mg/
kg b.w.), TP300(60% HFD+TP 300 mg/kg b.w.), TP600 (60% HFD+TP 600 mg/kg b.w.)과 같이 8개의 군으로 분 류하여 12주간 진행하였으며 주 2회 간격으로 체중과 식이 섭취량을 측정하였다.
혈액분석
실험동물의 실험 종료 시 12시간 절식시킨 후 isofluran, O2 및 N2 가스를 사용하는 호흡마취기기를 이용하여 마취시 키고, capillary tube를 이용하여 안와정맥총에서 채혈을 진 행하였다. 혈액은 원심 분리(16,000 rpm, 20 min, 4°C)하여 혈청을 분리한 후 triglyceride(TG), total cholesterol(TC), LDL/VLDL-cholesterol, HDL-cholesterol은 BioVision assay kit을 사용하여 측정하였으며, adiponectin 및 leptin level은 R&D System Quantikine ELISA kit(Bio-Techne, Minneapolis, MN, USA)을 사용하여 측정하였다.
조직분석
실험동물의 채혈 후 즉시 간 및 지방조직(장간막 지방, 피하지방, 부고환지방, 갈색지방)을 적출하였고, 생리식염수 로 세척하여 여과지로 수분을 제거한 후 중량을 측정하였다.
통계처리
본 실험은 SPSS(Statistical Package for Social Sci- ence) version 22.0 통계프로그램(IBM Corp., New York, NY, USA)을 이용하여 분석하였으며, 모든 측정 결과는 각 실험군의 평균(mean)±표준편차(SD)로 표시하였다. 그룹 간의 통계적 유의성은 Student’s t-test 및 Duncan’s mul- tiple range test를 이용하여 검증하였고, 실험군 간의 유의 성을 P<0.05 수준에서 확인하였다.
결과 및 고찰
세포독성
TP의 3T3-L1 세포에 대한 세포독성을 확인한 결과는 Fig. 1에 나타내었다. 선행연구에서 확인된 TP의 독성범위 인 100 μg/mL의 농도에 기초하여 TP를 0~100 μg/mL의 농도로 처리한 후 세포생존율을 확인한 결과 0~80 μg/mL 농도에서는 통계적 유의성이 나타나지 않았고, 90 μg/mL 이상의 농도에서는 세포생존율이 80% 미만으로 통계적으 로 유의한 세포독성이 나타났다. 따라서 세포생존율이 90.8
%인 80 μg/mL 농도를 최대 농도로 지정하여 추후 본 실험 을 수행하였다. Lee 등(26)은 가자 열매 메탄올추출물을 전 립선암세포 DU145에 0~100 μg/mL 농도로 처리하여 100 μg/mL 농도까지 80% 이상의 세포생존율이 나타났음을 보 고하였고, Lee 등(27)은 가자 열매 열수추출물이 primary cultured hepatocyte에서 0~1,000 μg/mL 농도까지 100%
이상의 생존율을 보였음을 보고하였다. Yahayo 등(28)은 암 라 열수추출물을 인간 섬유육종세포 HT1080에 0~50 μg/
mL 농도로 처리하여 20 μg/mL 농도 이상에서 생존율이 억 제되었고, 노출시간을 72시간까지 증가시키면 IC50 농도가 5 μg/mL로 세포생존력 또한 감소하였음을 보고하였다. 이 처럼 추출용매, 세포의 종류, 그리고 본 실험과 같이 추출물 의 혼합 여부 등에 따라 세포생존율이 달라지는 것을 확인할 수 있었다.
Oil red O 염색 및 lipid accumulation 측정
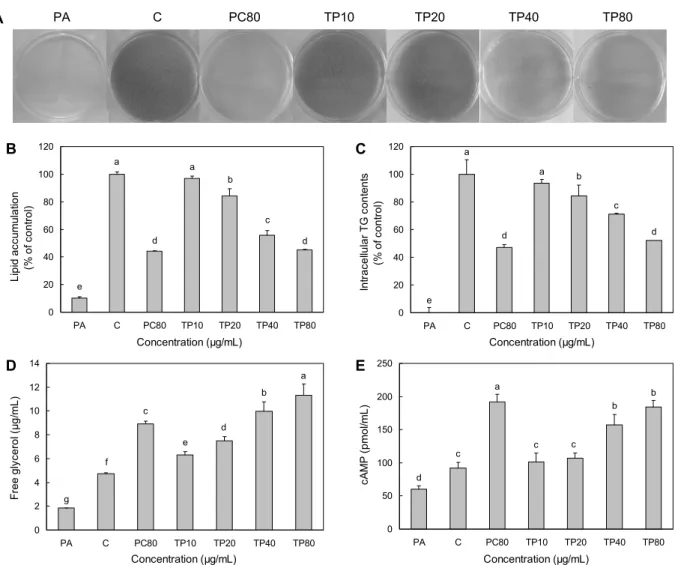
TP 처리에 의한 3T3-L1 세포의 분화 중 지방구의 생성 억제 정도를 확인하기 위하여 Oil red O 염색을 하였고, 그 결과는 Fig. 2A에 나타내었다. 지방구를 염색한 Oil red O dye를 용출하여 지방축적 함량을 측정한 결과(Fig. 2B) preadipocyte군에 비해 지방세포로 분화를 유도한 control 군이 89.8% 증가하여 통계적 유의성을 나타내었다. Con- trol군(100%)과 비교하였을 때 PC80군의 경우 44.27±
0.38%로 지방축적 함량이 감소하였으며 통계적 유의성을 나타내었다. TP를 처리한 군들의 경우 10, 20, 40, 80 μg/
mL 각각 96.78±1.94%, 84.40±4.94%, 55.70±3.57%, 45.18±0.27%로 지방축적 함량이 농도 의존적으로 감소하 였지만, 10 μg/mL 농도로 처리한 군에서는 통계적 유의성 이 나타나지 않았고 20, 40, 80 μg/mL 농도에서는 통계적 유의성이 나타났다. 특히 TP80군이 PC80군과 유사한 정도
A PA C PC80 TP10 TP20 TP40 TP80
d c b a
d a
e 0 20 40 60 80 100 120
PA C PC80 TP10 TP20 TP40 TP80
Concentration (μg/mL) Lipid accumulation . (% of control) .
B
d c a b
d a
e 0 20 40 60 80 100 120
PA C PC80 TP10 TP20 TP40 TP80
Concentration (μg/mL) Intracellular TG contents . (% of control) .
C
g f
c e
d b
a
0 2 4 6 8 10 12 14
PA C PC80 TP10 TP20 TP40 TP80
Concentration (μg/mL)
Free glycerol (μg/mL) .
D
b b
c c a
c d
0 50 100 150 200 250
PA C PC80 TP10 TP20 TP40 TP80
Concentration (μg/mL)
cAMP (pmol/mL) .
E
Fig. 2. Effects of TP on lipid accumulation, TG, glycerol release, and cAMP level in 3T3-L1 adipocytes. The differentiated 3T3-L1 adipocytes were treated with TP for 72 hours. (A) Oil red O staining, (B) lipid accumulation, (C) intracellular TG contents, (D) glycerol release, (E) intracellular cAMP level. PA, preadipocyte; C, differentiated control; PC80, positive control, catechin 80 µg/mL;
TP10, TP 10 μg/mL; TP20, TP 20 μg/mL; TP40, TP 40 μg/mL; TP80, TP 80 μg/mL. Values are mean±SD (n=3). Statistical analyses were performed by Duncan’s multiple range test and differences were considered statistically significant at P<0.05.
로 지방축적을 억제하였다. Kim 등(29) 및 Kim 등(30)은 3T3-L1 preadipocyte를 분화 시 시료의 처리로 지방생성 억제능을 확인하였고, Lee 등(21)은 지방세포가 100% 융 합된 상태에서 잣송이 초임계 추출물을 처리하여 지방분해 에 효과가 있는 것을 확인하였으며, 본 연구에서는 TP를 처 리하여 생성된 지방구의 분해능을 확인할 수 있었다.
세포 내 TG 함량의 측정
TP 처리에 의한 3T3-L1 세포의 분화 중 중성지방의 억제 정도를 보다 세밀하게 확인하기 위하여 세포 내 TG 함량을 측정하였고, 그 결과는 Fig. 2C에 나타내었다. Preadipo- cyte군에 비해 control군의 중성지방이 100% 증가하여 통 계적 유의성을 나타내었고, PC80군의 경우 control군과 비 교하였을 때 47.06±2.36%로 TG 함량이 감소하였으며 통 계적 유의성을 나타내었다. TP를 처리한 군들의 경우 10, 20, 40, 80 μg/mL 각각 93.66±6.23%, 84.45±3.60%, 71.26
±3.37%, 52.19±7.92%로 TG 함량이 농도 의존적으로 감 소하였지만, 10 μg/mL 농도로 처리한 군에서는 통계적 유 의성이 나타나지 않았고 20, 40, 80 μg/mL 농도에서는 통계 적 유의성이 나타났으며, 특히 Oil red O 염색의 결과와 마 찬가지로 TP80군이 PC80군과 유사한 정도로 TG 함량을 억제하였다. Cha 등(31)은 수세미오이 메탄올추출물이 분 화된 3T3-L1 세포에서 중성지방을 억제하였다고 보고하였 고, 본 연구에서도 마찬가지로 TP 처리에 의한 중성지방을 억제 효과를 확인할 수 있었다.
Glycerol release 측정
TP 처리에 의한 3T3-L1 세포의 분화 중 중성지방의 분 해 정도를 간접적으로 확인하기 위하여 유리된 free glyc- erol 함량을 측정하였고, 그 결과는 Fig. 2D에 나타내었다.
Preadipocyte군보다 control군의 free glycerol 함량이 60.8% 증가하여 통계적 유의성을 나타내었고, PC80군의
bc c ab a
c a
c
c b d d
bc
d a
c b a a
c a c
0 0.5 1 1.5 2 2.5
PA C PC80 TP10 TP20 TP40 TP80
Concentration (μg/mL)
mRNA expression of lipolytic genes . PDE3B PKA Perilipin
b a c c
a e d
a
b cd c
ab
de e
0 0.5 1 1.5 2 2.5 3 3.5
PA C PC80 TP10 TP20 TP40 TP80
Concentration (μg/mL)
mRNA expression of lipases . HSL ATGL
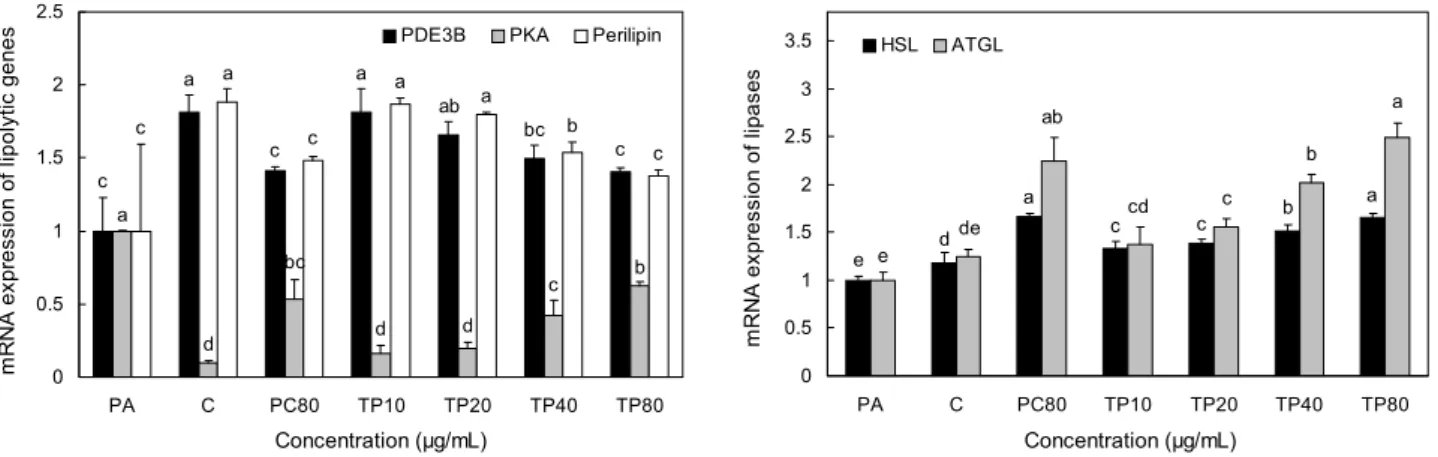
Fig. 3. Effects of TP on mRNA expressions of lipolytic genes in 3T3-L1 adipocytes. The differentiated 3T3-L1 adipocytes were treated with TP for 72 hours. Data were calculated as the relative of lipase mRNA expression versus GAPDH as a internal control.
PA, preadipocyte; C, differentiated control; PC80, positive control, catechin 80 µg/mL; TP10, TP 10 μg/mL; TP20, TP 20 μg/mL;
TP40, TP 40 μg/mL; TP80, TP 80 μg/mL. Values are mean±SD (n=3). Statistical analyses were performed by Duncan’s multiple range test and differences were considered statistically significant at P<0.05.
경우 control군보다 88.2% 증가하여 통계적 유의성을 나타 내었다. TP를 처리한 군들의 경우 control군보다 10, 20, 40, 80 μg/mL 각각 33.0%, 58.5%, 110.9%, 139.0% 증가 하였고 통계적 유의성이 나타났으며, 특히 TP80군이 PC80 군보다 더 많은 양의 free glycerol을 방출하였다. TG의 분 해산물 중 하나인 glycerol은 인체의 경우 혈액으로 유리되 고 간으로 이동하며, 이 분해 작용은 주로 insulin 의존 li- pase인 HSL이 매개가 되어 일어난다(4,32). 본 연구에서는 Lee 등(33)의 연구와 마찬가지로 세포배양액으로 방출된 glycerol의 함량을 측정함으로써 중성지방의 분해 정도를 확인할 수 있었다.
세포 내 cAMP level 측정
TP 처리가 지방세포의 분해과정에서 cAMP level에 미 치는 영향을 Fig. 2E에 나타내었다. Preadipocyte군보다 control군의 cAMP level이 35.1% 증가하여 통계적 유의성 을 나타내었고, PC80군의 경우 control군보다 107.9% 증 가하였으며 통계적 유의성을 나타내었다. TP를 처리한 군 들의 경우 control군보다 10, 20, 40, 80 μg/mL 각각 10.2
%, 15.8%, 70.1%, 99.5% 증가하였으나, 10, 20 μg/mL 농 도로 처리한 군에서는 통계적 유의성이 나타나지 않았고 40, 80 μg/mL 농도에서는 통계적 유의성이 나타났다. 특히 TP80군이 PC80군과 유사한 정도로 cAMP level을 증가시 켰다. cAMP는 PDE3B에 의해 조절되며(34), cAMP의 증가 는 PKA의 활성을 증가시켜 지방분해에 있어 중요한 기전으 로 작용한다(5,6). 본 연구에서는 Ho 등(25)의 연구 결과와 같이 시료의 처리로 인한 cAMP level의 증가를 확인할 수 있었다.
지방분해 관련 유전자의 mRNA 발현
TP 처리가 지방분해 관련 유전자의 mRNA 발현에 미치 는 영향을 조사하기 위하여 PDE3B, PKA, Perilipin, HSL
및 ATGL의 발현량을 real-time PCR을 이용하여 확인하였 고, 그 결과는 Fig. 3에 나타내었다. Phosphodiesterase family(PDE3A, PDE3B) 중 하나로 cAMP를 가수분해 시켜 지방세포에서 항지방분해 활성에 관여하는 것으로 알려진 (26) PDE3B mRNA는 control군에서 preadipocyte군에 비 해 44.8% 증가하였고 통계적 유의성이 나타났으며, PC80 군에서는 control군보다 22.3% 감소하였고 통계적 유의성 이 나타났다. TP를 처리한 군들의 경우 control군에 비해 10, 20, 40, 80 μg/mL 각각 0.0%, 8.7%, 17.5%, 22.7%
감소하였으나, 40, 80 μg/mL 농도에서만 통계적 유의성이 나타났으며, 특히 TP80군은 PC80군과 유사한 정도로 감소 하였다.
cAMP에 의존적이며 HSL의 인산화에 관여하는(5) PKA mRNA는 control군에서 preadipocyte군에 비해 10.1배 감 소하였고 통계적 유의성이 나타났으며, PC80군에서는 con- trol군보다 5.4배 증가하였고 통계적 유의성이 나타났다. TP 를 처리한 군들의 경우 control군보다 10, 20, 40, 80 μg/
mL 각각 1.6배, 2.0배, 4.3배, 6.3배 증가하였으나, 40, 80 μg/mL 농도에서만 통계적 유의성이 나타났으며 PC80군과 유사한 정도로 증가하였다.
지방구의 보호막으로서 HSL의 활성을 조절하는 인자인 (35) Perilipin mRNA는 control군에서 preadipocyte군에 비해 46.8% 증가하였고 통계적 유의성이 나타났으며, PC 80군에서는 control군보다 21.3% 감소하였고 통계적 유의 성이 나타났다. TP를 처리한 군들의 경우 control군에 비해 10, 20, 40, 80 μg/mL 각각 0.7%, 4.4%, 18.0%, 26.9%
감소하였으나, 40, 80 μg/mL 농도에서만 통계적 유의성이 나타났으며, 특히 TP80군은 PC80군과 유사한 정도로 감소 하였다.
지방분해는 PDE3B에 의해 조절되는 cAMP의 세포 내 수준에 결정적으로 의존한다(34). PDE3 gene family는 PDE3A와 PDE3B로 구성되어 있고, cAMP를 5’AMP로 분
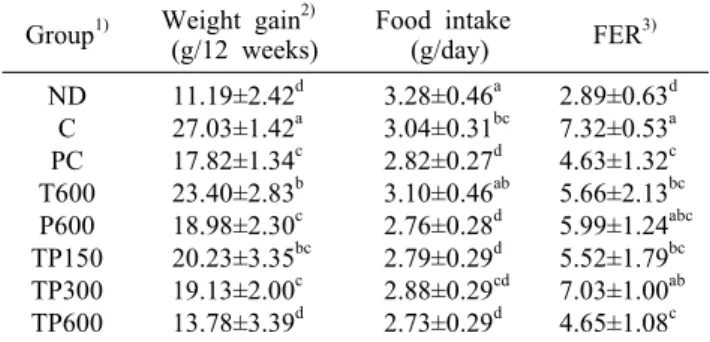
Table 2. The effect of TP on body weight gain and food effi- ciency rate of experimental groups for 12 weeks
Group1) Weight gain2)
(g/12 weeks) Food intake
(g/day) FER3) ND
C PC T600 P600 TP150 TP300 TP600
11.19±2.42d 27.03±1.42a 17.82±1.34c 23.40±2.83b 18.98±2.30c 20.23±3.35bc 19.13±2.00c 13.78±3.39d
3.28±0.46a 3.04±0.31bc 2.82±0.27d 3.10±0.46ab 2.76±0.28d 2.79±0.29d 2.88±0.29cd 2.73±0.29d
2.89±0.63d 7.32±0.53a 4.63±1.32c 5.66±2.13bc 5.99±1.24abc 5.52±1.79bc 7.03±1.00ab 4.65±1.08c
1)ND, normal diet; C, high fat diet (HFD); PC, positive control (HFD+catechin 1 g/kg b.w.); T600, HFD+T600 mg/kg b.w.;
P600, HFD+P600 mg/kg b.w.; TP150, HFD+TP150 mg/kg b.w.; TP300, HFD+TP300 mg/kg b.w.; TP600, HFD+TP600 mg/kg b.w..
2)Weight gain (g)=final body weight (g)-initial body weight (g).
3)FER (food efficiency rate)=gain of body weight (g)/ food in- take calories (kcal).
Values are mean±SD (n=8).
Different superscript letters within a column indicate significant differences by ANOVA with Duncan’s multiple range test at P<0.05.
해하는데 이 중 PDE3B는 간세포 및 췌장의 β 세포를 포함 한 인슐린 감수성 세포에서 발현되며, anti-glycogenolysis, anti-lipolysis 및 인슐린 분비에 핵심적인 역할을 한다(34, 36). PDE3B의 감소는 cAMP의 가수분해를 감소시키고, cAMP에 의존하는 PKA의 활성을 증가시킨다. PKA에 의한 HSL과 Perilipin의 인산화는 지방분해를 효율적으로 증가 시키는데(5,6,37), 세포질에서 지방구로 HSL이 빠르게 이동 하고 지방세포에 저장된 TG의 가수분해가 촉진된다(5,6).
지방세포의 지방구 표면에 위치한 Perilipin은 lipases의 공 격으로부터 TG를 보호하는 역할을 하지만(6,38), 인산화된 Perilipin은 지방구 내의 HSL과 TG의 상호작용을 위해 지방 구의 표면을 변형시킬 수 있다(6,39). 몇몇 연구들은 Perili- pin의 수치 감소가 지방분해를 증가시킨다는 것을 입증했다 (38,39). 본 연구에서는 TP의 처리가 PDE3B를 감소시키고 cAMP와 PKA를 활성화해 지방구 표면 위로 올라온 HSL의 발현을 증가시켰으며, Perilipin의 발현을 감소시킨 것으로 확인되었다. 이는 지방분해 관련 유전자의 발현을 통한 지방 분해 효과를 제시한 Ho 등(25)의 연구 결과와도 일치한다.
HSL의 mRNA는 control군에서 preadipocyte군에 비해 17.8% 증가하였고 통계적 유의성이 나타났으며, PC80군에 서는 control군보다 41.0% 증가하였고 통계적 유의성이 나 타났다. TP를 처리한 군들의 경우 control군에 비해 10, 20, 40, 80 μg/mL에서 각각 12.8%, 17.3%, 28.0%, 40.0% 증 가하였고, 통계적 유의성이 나타났다. 특히 TP80은 PC80 군과 유사한 정도로 증가하였다.
ATGL의 mRNA는 control군에서 preadipocyte군에 비 해 24.0% 증가하였으나 통계적 유의성이 나타나지 않았고, PC80군에서는 control군에 비해 80.9% 증가하였고 통계적 유의성이 나타났다. TP를 처리한 군들의 경우 control군에 비해 10, 20, 40, 80 μg/mL 각각 11.0%, 25.1%, 62.6%, 100.8% 증가하였으나, 10 μg/mL 농도에서는 통계적 유의 성이 나타나지 않았고 20, 40, 80 μg/mL 농도에서는 통계적 유의성이 나타났으며 PC80군과 유사한 정도로 증가하였다.
지방조직에 저장된 TG는 HSL, ATGL 및 monoacylgly- cerol lipase(MAGL)와 같은 lipases에 의해 지방산과 글리 세롤로 가수분해 된다(4,40,41). HSL과 ATGL은 주로 지방 조직에서 발현되는데 HSL은 TG와 diacylglycerol(DG)을 가수분해 하는 효소로서 지방분해의 속도를 제한하는 역할 을 하고(9), ATGL은 TG의 가수분해 시 발현이 되며 DG와 지방산을 생산한다(9,42). 본 연구에서는 TP의 처리가 HSL 과 ATGL의 발현을 증가시켰음을 확인하였고, 이는 지방분 해효소의 유전자 발현을 확인한 Ho 등(25) 및 Lee 등(33)의 연구 결과와 일치하였으며, TP가 TG에서 DG로 가수분해 되는 첫 단계에서 효과적으로 작용한 것으로 생각한다.
실험동물의 체중증가량, 식이섭취효율, 조직 무게의 변화 12주간 실험기간의 체중증가량과 일일 식이섭취량의 변 화는 Table 2에 나타내었다. C군의 체중증가량이 ND군보
다 141.6% 증가하여 비만이 유도되었음을 확인하였으며, PC군의 경우 C군보다 34.1% 감소하였고 통계적 유의성이 나타났다. TP군들(TP150, 300, 600)의 경우 C군에 비해 각각 25.2%, 29.2%, 49.0% 감소하였고 통계적 유의성이 나타났으며, 특히 TP300과 TP600군에서 PC군보다 체중 증가량이 더 많이 감소하였음을 확인하였다.
식이섭취효율의 경우 ND군이 2.89, C군이 7.32로 C군이 ND군보다 높았으며, TP를 섭취한 군들 간의 유의성은 나타 나지 않았다.
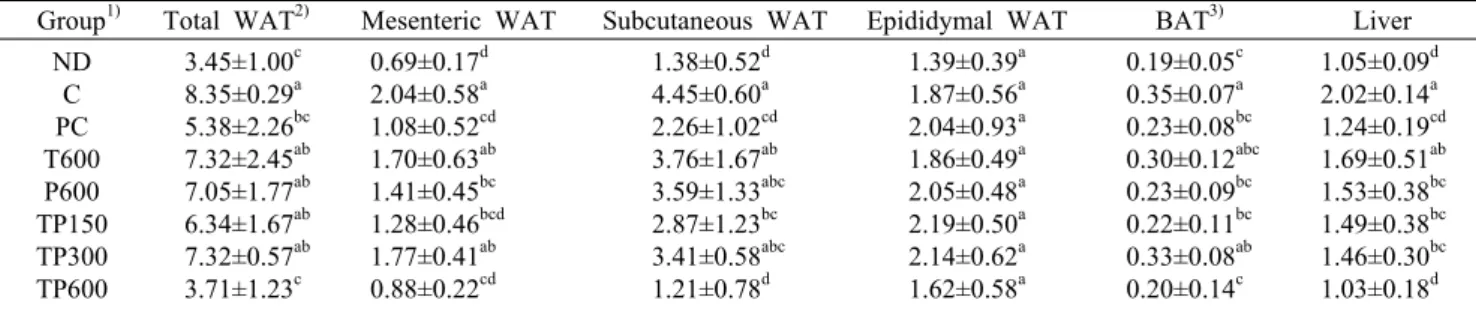
지방조직 무게는 백색지방(장간막 지방, 피하지방, 부고 환지방)과 갈색지방을 각각 측정하였다. 장간막 지방 무게 의 경우 ND군보다 C군에서 197.0% 증가하여 통계적 유의 성이 나타났고, PC군의 경우 C군보다 47.2% 감소하였음을 확인하였다. TP군들(TP150, 300, 600)의 경우 C군보다 각 각 37.2%, 13.3%, 56.9% 감소하였고, 특히 TP600군에서 통계적 유의성을 보여 장간막 지방이 가장 많이 감소하였음 을 확인하였다. 피하지방 무게의 경우 역시 장간막 지방 무 게와 같은 경향이 나타났으나, 부고환지방무게의 경우 모든 군에서 통계적 유의성이 나타나지 않았다(Table 3).
혈청 내 지질(TG, TC, LDL-cholesterol, HDL-choles- terol) 변화
혈청 내 TG, TC, LDL-cholesterol, HDL-cholesterol 의 농도를 측정한 결과는 Table 4에 나타내었다. TG의 경우 고지방식이에 의해 가장 많이 영향을 받는 인자로 ND군보 다 C군의 TG 농도가 98.9% 증가하여 통계적 유의성이 나타 났음을 확인하였으며, PC군의 경우 C군보다 65.3% 감소하 여 통계적인 유의성이 나타났다. TP군들(TP150, 300, 600)
Table 3. Adipose tissue and liver weight of experimental groups for 12 weeks
Group1) Total WAT2) Mesenteric WAT Subcutaneous WAT Epididymal WAT BAT3) Liver ND
C PC T600 P600 TP150 TP300 TP600
3.45±1.00c 8.35±0.29a 5.38±2.26bc 7.32±2.45ab 7.05±1.77ab 6.34±1.67ab 7.32±0.57ab 3.71±1.23c
0.69±0.17d 2.04±0.58a 1.08±0.52cd 1.70±0.63ab 1.41±0.45bc 1.28±0.46bcd 1.77±0.41ab 0.88±0.22cd
1.38±0.52d 4.45±0.60a 2.26±1.02cd 3.76±1.67ab 3.59±1.33abc 2.87±1.23bc 3.41±0.58abc 1.21±0.78d
1.39±0.39a 1.87±0.56a 2.04±0.93a 1.86±0.49a 2.05±0.48a 2.19±0.50a 2.14±0.62a 1.62±0.58a
0.19±0.05c 0.35±0.07a 0.23±0.08bc 0.30±0.12abc 0.23±0.09bc 0.22±0.11bc 0.33±0.08ab 0.20±0.14c
1.05±0.09d 2.02±0.14a 1.24±0.19cd 1.69±0.51ab 1.53±0.38bc 1.49±0.38bc 1.46±0.30bc 1.03±0.18d
1)ND, normal diet; C, high fat diet (HFD); PC, positive control (HFD+catechin 1 g/kg b.w.); T600, HFD+T600 mg/kg b.w.; P600, HFD+P600 mg/kg b.w.; TP150, HFD+TP150 mg/kg b.w.; TP300, HFD+TP300 mg/kg b.w.; TP600, HFD+TP600 mg/kg b.w..
2)WAT: white adipose tissue.
3)BAT: brown adipose tissue.
Values are mean±SD (n=8).
Different superscript letters within a column indicate significant differences by ANOVA with Duncan’s multiple range test at P<0.05.
Table 4. Serum lipid profile, leptin, and adiponectin of ND and C57BL/6J mice fed high fat diet with TP for 12 weeks Group1) Triglyceride
(mM) Total cholesterol
(μg/μL) LDL-cholesterol
(μg/μL) HDL-cholesterol (μg/μL)
LDL-/HDL- cholesterol
(μg/μL)
Adiponectin
(ng/μL) Leptin (pg/μL) ND
C PC T600 P600 TP150 TP300 TP600
4.75±1.01bc 9.45±1.42a 3.27±1.24cd 5.22±2.32b 4.27±1.31bc 4.59±0.43bc 3.46±0.54cd 2.21±0.52d
4.98±0.16ab 5.30±0.39a 4.50±0.33cd 5.09±0.06a 4.21±0.50d 4.70±0.17bc 4.53±0.11cd 4.30±0.16d
1.33±0.05c 1.99±0.06a 1.20±0.07d 1.71±0.06b 1.41±0.15c 1.43±0.11c 1.33±0.08c 1.19±0.07d
2.83±0.14a 1.39±0.14e 2.79±0.24a 2.30±0.16c 1.83±0.11d 2.36±0.12c 2.57±0.09b 2.78±0.15a
0.47±0.03de 1.45±0.17a 0.43±0.04e 0.75±0.06b 0.77±0.06b 0.61±0.04c 0.52±0.04d 0.43±0.05e
8.94±0.99a 4.46±0.40e 7.68±0.22b 4.77±0.16e 4.69±0.12e 4.87±0.15e 5.43±0.13d 6.49±0.56c
873.64±406.92d 3,534.00±118.08a 1,256.50±1006.24cd 2,462.33±689.84b 2,568.17±614.82b 2,658.17±372.30b 1,838.17±107.86bc 804.00±570.39d
1)ND, normal diet; C, high fat diet (HFD); PC, positive control (HFD+catechin 1 g/kg b.w.); T600, HFD+T600 mg/kg b.w.; P600, HFD+P600 mg/kg b.w.; TP150, HFD+TP150 mg/kg b.w.; TP300, HFD+TP300 mg/kg b.w.; TP600, HFD+TP600 mg/kg b.w..
Values are mean±SD (n=8).
Different superscript letters within a column indicate significant differences by ANOVA with Duncan’s multiple range test at P<0.05.
의 경우 C군에 비해 각각 농도 의존적으로 51.5%, 63.4%, 76.6% 감소하였고, 통계적 유의성을 나타냈다.
TC의 경우 ND군보다 C군의 농도가 6.4% 증가하는 경향 을 보였으나 유의성이 나타나지는 않았고, PC군의 경우 C군 에 비해 15.1% 감소하였고, 통계적 유의성이 나타났다. TP 군들(TP150, 300, 600)의 경우 C군에 비해 각각 농도 의존 적으로 11.2%, 14.6%, 18.8% 감소하였고, 통계적 유의성 을 나타냈다.
LDL-cholesterol의 경우 C군의 농도가 ND군보다 49.1
% 증가하여 통계적 유의성이 나타났고, PC군의 경우 C군보 다 39.8% 감소하여 통계적 유의성이 나타났다. TP군들 (TP150, 300, 600)의 경우 C군에 비해 각각 농도 의존적으 로 28.1%, 32.9%, 40.4% 감소하였고 통계적 유의성이 나타 났으며, 특히 TP600군의 경우 PC군과 유사한 정도의 효과 를 나타내었다.
HDL-cholesterol의 경우 ND군보다 C군의 농도가 50.9
% 감소하여 통계적 유의성이 나타났고, PC군의 경우 C군보 다 101.2% 증가하여 통계적 유의성이 나타났다. TP군들 (TP150, 300, 600)의 경우 C군에 비해 각각 농도 의존적으 로 70.0%, 84.8%, 100.1% 증가하였고, 통계적 유의성이
나타났다.
LDL-cholesterol은 주로 혈관벽에 붙어 동맥경화나 심 장질환의 원인이 되는 인자로서 혈중 농도가 높아질수록 위 험성이 커지며, HDL-cholesterol은 주로 혈관벽에 축적된 콜레스테롤을 제거하고 간조직으로 운반하여 조직이나 혈 청의 콜레스테롤 농도를 재조정함으로써 혈중 콜레스테롤 의 농도를 저하하는 역할을 하는데, 본 실험의 결과는 고지 방식이가 정상군보다 TG, TC, LDL-cholesterol의 농도를 증가시킨 다른 연구 결과들과도 일치한다(43,44).
혈청 내 adiponectin 및 leptin 농도의 변화
혈청 내 adiponectin 및 leptin의 농도를 측정한 결과는 Table 4에 나타내었다. Adiponectin의 경우 ND군보다 C군 의 농도가 50.2% 감소하여 통계적 유의성이 나타났고, PC 군의 경우 C군보다 72.4% 증가하여 통계적 유의성이 나타 났다. TP군들(TP150, 300, 600)의 경우 C군에 비해 각각 농도 의존적으로 9.2%, 21.8%, 45.5% 증가하였으나, TP 300군과 TP600군에서만 통계적 유의성이 나타났다. Adi- ponectin의 농도는 비만인에서 정상인보다 낮게 나타나고 염증반응 및 공복 인슐린 농도와 음의 상관관계에 있는 것으
로 알려져 있으며(45), 본 연구에서도 같은 결과를 확인하였 다.
Leptin의 경우 ND군보다 C군의 농도가 304.5% 증가하 여 통계적 유의성이 나타났고, PC군의 경우 C군보다 64.4%
감소하여 통계적 유의성이 나타났다. TP군들(TP150, 300, 600)의 경우 C군에 비해 각각 농도 의존적으로 24.8%, 48.0%, 77.2% 감소하였고, 모든 군에서 통계적 유의성이 나타났다. Leptin은 지방세포에서 분비되는 호르몬으로 식 욕 억제와 포만감을 느끼게 하여 식욕 및 에너지 대사를 조 절하는 것으로 알려져 있다. Galic 등(46) 및 Friedman과 Halaas(47)는 비만환자뿐만 아니라 고지방 섭취가 실험동 물의 leptin 농도를 증가시켰다고 보고하였으며, 본 연구에 서도 같은 결과를 확인할 수 있었다.
요 약
본 연구에서는 가자와 암라 복합추출물(TP)이 분화된 3T3- L1 세포와 고지방식이로 유도된 비만동물모델에 미치는 항 비만 효과를 관찰하였다. 분화된 3T3-L1 세포에 TP 0~80 μg/mL 농도를 처리하여 lipid accumulation, TG, glycerol, cAMP, 그리고 지방분해 관련 유전자들을 측정한 결과, 40 μg/mL 이상의 농도에서 세포 내 축적된 지방구들이 분해되 어 glycerol 방출량을 증가시켰고, PDE3B의 발현을 감소시 켜 cAMP가 가수분해 되지 않게 조절하여 PKA, HSL, 그리 고 ATGL의 발현을 증가시켰으며, 지방구를 둘러싸고 있는 Perilipin의 발현을 감소시킨 것으로 보아 PKA를 촉진하는 cAMP 경로의 지방분해 기전이 작용하였음을 확인하였다.
고지방식이로 비만을 유도한 C57BL/6J 마우스를 이용하여 항비만 활성을 관찰한 결과, TP의 섭취가 체중증가량, 장간 막 지방, 피하지방, 그리고 혈액 내 TG, TC, LDL-choles- terol, leptin의 함량을 대조군에 비해 농도 의존적으로 감소 시켰고, HDL-cholesterol 및 adiponectin의 함량을 유의적 으로 증가시킨 것으로 보아 TP가 비만 관련 호르몬 및 지방 분해 효소들을 조절한 것으로 여겨진다. 특히 동물모델에서 단독추출물보다 복합추출물일 때 지방분해에 있어 시너지 효과가 나타난 것을 확인하였다. 따라서 TP가 지방분해 기 전인 cAMP 경로를 활성화하고 관련 유전자와 효소들을 조 절하여 최종적으로 체중감소에 도움이 되는 원료로 기능성 이 입증되어 항비만 활성에 대한 기초자료로의 활용을 기대 할 수 있다.
감사의 글
본 연구는 산업통상자원부와 한국산업기술진흥원의 “사업 화연계기술개발사업”(과제번호: N0002384, 2017)의 지원 에 의해 수행되었습니다.
REFERENCES
1. Spiegelman BM, Flier JS. 1996. Adipogenesis and obesity;
rounding out the big picture. Cell 87: 377-389.
2. Tateya S, Kim F, Tamori Y. 2013. Recent advances in obe- sity-induced inflammation and insulin resistance. Front En- docrinol 4: 93.
3. Frayn KN, Karpe F, Fielding BA, Macdonald IA, Coppack SW. 2003. Integrative physiology of human adipose tissue.
Int J Obes Relat Metab Disord 27: 875-888.
4. Ahmadian M, Wang Y, Sul HS. 2010. Lipolysis in adipocytes.
Int J Biochem Cell Biol 42: 555-559.
5. Holm C. 2003. Molecular mechanisms regulating hormone- sensitive lipase and lipolysis. Biochem Soc Trans 31: 1120- 1124.
6. Carmen GY, Victor SM. 2006. Signaling mechanisms regu- lating lipolysis. Cell Signal 18: 401-408.
7. Rayalam S, Della-Fera MA, Baile CA. 2008. Phytochemicals and regulation of the adipocyte life cycle. J Nutr Biochem 19: 717-726.
8. Haemmerle G, Lass A, Zimmermann R, Gorkiewicz G, Meyer C, Rozman J, Heldmaier G, Maier R, Theussl C, Eder S, Kratky D, Wagner EF, Klingenspor M, Hoefler G, Zechner R. 2006. Defective lipolysis and altered energy metabolism in mice lacking adipose triglyceride lipase. Science 312:
734-737.
9. Zechner R, Strauss JG, Haemmerle G, Lass A, Zimmermann R. 2005. Lipolysis: pathway under construction. Curr Opin Lipidol 16: 333-340.
10. Ntambi JM, Kim YC. 2000. Adipocyte differentiation and gene expression. J Nutr 130: 3122S-3126S.
11. Juang LJ, Sheu SJ. 2005. Chemical identification of the sources of commercial Fructus Chebulae. Phytochem Anal 16: 246-251.
12. SuryaPrakash DV, Sree Satya N, Sumanjali A, Meena V.
2012. Pharmacological review on Terminalia chebula. Inter J Res Pharm Biomed Sci 3: 679-683.
13. Saleem A, Husheem M, Harkonen P, Pihlaja K. 2002. Inhi- bition of cancer cell growth by crude extract and the phe- nolics of Terminalia chebula Retz. fruit. J Ethnopharmacol 81: 327-336.
14. Sabu MC, Kuttan R. 2002. Anti-diabetic activity of medici- nal plants and its relationship with their antioxidant property.
J Ethnopharmacol 81: 155-160.
15. Kaur S, Arora S, Kaur K, Kumar S. 2002. The in vitro anti- mutagenic activity of Triphala-an Indian herbal drug. Food Chem Toxicol 40: 527-534.
16. Malekzadeh F, Ehsanifar H, Shahamat M, Levin M, Colwell RR. 2001. Antibacterial activity of black myrobalan (Termi- nalia chebula Retz) against Helicobacter pylori. Int J Anti- microb Agents 18: 85-88.
17. Gurjar S, Pal A, Kapur S. 2012. Triphala and its constituents ameliorate visceral adiposity from a high-fat diet in mice with diet-induced obesity. Altern Ther Health Med 18(6):
38-45.
18. Dasaroju S, Gottumukkala KM. 2014. Current trends in the research of Emblica officinalis (Amla): A pharmacological perspective. Int J Pharm Sci Rev Res 24: 150-159.
19. Kumar KPS, Bhowmik D, Dutta A, Pd.Yadav A, Paswan S, Srivastava S, Deb L. 2012. Recent trends in potential tra- ditional indian herbs Emblica officinalis and its medicinal importance. J Pharmacogn Phytochem 1: 24-32.
20. Krishnaveni M, Mirunalini S. 2010. Therapeutic potential of Phyllanthus emblica (amla): the ayurvedic wonder. J Basic
Clin Physiol Pharmacol 21: 93-105.
21. Lee M, Nam DE, Kim OK, Heo SH, Lee J. 2014. Lipolytic effect of supercritical extraction from pine cone (Pinus kor- aiensis) in mature 3T3-L1 adipocytes. J Korean Soc Food Sci Nutr 43: 1342-1348.
22. Berridge MV, Tan AS. 1993. Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetra- zolium bromide (MTT): subcellular localization, substrate dependence, and involvement of mitochondrial electron trans- port in MTT reduction. Arch Biochem Biophys 303: 474- 482.
23. Gao CL, Zhu JG, Zhao YP, Chen XH, Ji CB, Zhang CM, Zhu C, Xia ZK, Peng YZ, Guo XR. 2010. Mitochondrial dysfunction is induced by the overexpression of UCP4 in 3T3-L1 adipocytes. Int J Mol Med 25: 71-80.
24. Trinder P. 1969. Determination of blood glucose using an oxidase-peroxidase system with a non-carcinogenic chromo- gen. J Clin Pathol 22: 158-161.
25. Ho JN, Kim OK, Nam DE, Jun W, Lee J. 2014. Pycnogenol supplementation promotes lipolysis via activation of cAMP- dependent PKA in ob/ob mice and primary-cultured adipo- cytes. J Nutr Sci Vitaminol 60: 429-435.
26. Lee YH. 2013. The inhibitory effect in androgen receptor- dependent prostate cancer cell growth by anti-histone ace- tyltransferase activity from Terminalia chebula Retz. fruit methanol extract. J Korean Soc Food Sci Nutr 42: 1539- 1543.
27. Lee HS, Won NH, Kim KH, Lee H, Jun W, Lee KW. 2005.
Antioxidant effects of aqueous extract of Terminalia cheb- ula in vivo and in vitro. Biol Pharm Bull 28: 1639-1644.
28. Yahayo W, Supabphol A, Supabphol R. 2013. Suppression of human fibrosarcoma cell metastasis by Phyllanthus embl- ica extract in vitro. Asian Pac J Cancer Prev 14: 6863-6867.
29. Kim J, Park J, Jun W. 2014. Anti-obesity effect of ethyl ace- tate fraction from 50% ethanol extract of fermented Curcu- ma longa L. 3T3-L1 cells. J Korean Soc Food Sci Nutr 43:
1681-1687.
30. Kim SO, Kim MR, Hwang KA, Park NJ, Jeong JS. 2017.
Inhibition of differentiation and anti-adipogenetic effect of the Salvia plebeia R. Br. ethanol extract in murine adipo- cytes, 3T3-L1 cells. J Korean Soc Food Sci Nutr 46: 401- 408.
31. Cha SY, Jang JY, Lee YH, Lee G, Lee HJ, Hwang KT, Kim Y, Jun W, Lee J. 2010. Lipolytic effect of methanol extracts from Luffa cylindrica in mature 3T3-L1 adipocytes. J Kore- an Soc Food Sci Nutr 39: 813-819.
32. Degerman E, Belfrage P, Manganiello VC. 1997. Structure, localization, and regulation of cGMP-inhibited phosphodies- terase (PDE3). J Biol Chem 272: 6823-6826.
33. Lee J, Yoon HG, Lee YH, Park J, You Y, Kim K, Jang JY, Yang JW, Jun W. 2010. The potential effects of ethyl acetate fraction from Curcuma longa L. on lipolysis in dif-
ferentiated 3T3-L1 adipocytes. J Med Food 13: 364-370.
34. Rahn Landstrom T, Mei J, Karlsson M, Manganiello V, Degerman E. 2000. Down-regulation of cyclic-nucleotide phosphodiesterase 3B in 3T3-L1 adipocytes induced by tu- mor necrosis factor α and cAMP. Biochem J 346: 337-343.
35. Greenberg AS, Egan JJ, Wek SA, Garty NB, Blanchette- Mackie EJ, Londos C. 1991. Perilipin, a major hormonally regulated adipocyte-specific phosphoprotein associated with the periphery of lipid storage droplets. J Biol Chem 266:
11341-11346.
36. Zmuda-Trzebiatowska E, Oknianska A, Manganiello V, De- german E. 2006. Role of PDE3B in insulin-induced glucose uptake, GLUT-4 translocation and lipogenesis in primary rat adipocytes. Cell Signal 18: 382-390.
37. Jaworski K, Sarkadi-Nagy E, Duncan RE, Ahmadian M, Sul HS. 2007. Regulation of triglyceride metabolism. Ⅳ. Hor- monal regulation of lipolysis in adipose tissue. Am J Physiol Gastrointest Liver Physiol 293: G1-G4.
38. Akter MH, Yamaguchi T, Hirose F, Osumi T. 2008. Perilipin, a critical regulator of fat storage and breakdown, is a target gene of estrogen receptor-related receptor α. Biochem Bio- phys Res Commun 368: 563-568.
39. Miyoshi H, Souza SC, Zhang HH, Strissel KJ, Christoffolete MA, Kovsan J, Rudich A, Kraemer FB, Bianco AC, Obin MS, Greenberg AS. 2006. Perilipin promotes hormone-sen- sitive lipase-mediated adipocyte lipolysis via phosphorylat- ion-dependent and -independent mechanisms. J Biol Chem 281: 15837-15844.
40. Gaidhu MP, Anthony NM, Patel P, Hawke TJ, Ceddia RB.
2010. Dysregulation of lipolysis and lipid metabolism in vis- ceral and subcutaneous adipocytes by high-fat diet: role of ATGL, HSL, and AMPK. Am J Physiol Cell Physiol 298:
C961-C971.
41. Ducharme NA, Bickel PE. 2008. Lipid droplets in lipogenesis and lipolysis. Endocrinology 149: 942-949.
42. Bamba V, Rader DJ. 2007. Obesity and atherogenic dyslip- idemia. Gastroenterology 132: 2181-2190.
43. Bae GJ, Son JH, Lee JH, Jeong NO, Ha BJ. 2014. Effects of Cladosiphon okamuranus dietary fiber on cholesterol in high fat diet-fed rats. J Food Hyg Safety 29: 370-375.
44. Kim SW, Kim NS, Oh MJ, Kim HR, Kim MS, Lee DY, Yoon SH, Jung MY, Kim HJ, Lee CH, Oh CH. 2017. Anti- obesity effects of fermented soybean oils in 3T3-L1 pre-adi- pocytes and high fat diet-fed C57BL/6J mice. J Korean Soc Food Sci Nutr 46: 279-288.
45. Matsuzawa Y, Funahashi T, Kihara S, Shimomura I. 2004.
Adiponectin and metabolic syndrome. Arterioscler Thromb Vasc Biol 24: 29-33.
46. Galic S, Oakhill JS, Steinberg GR. 2010. Adipose tissue as an endocrine organ. Mol Cell Endocrinol 316: 129-139.
47. Friedman JM, Halaas JL. 1998. Leptin and the regulation of body weight in mammals. Nature 395: 763-770.