Jour. Korean For. Soc. Vol. 99, No. 3, pp. 312~322 (2010)
312
JOURNAL OF KOREAN FOREST SOCIETY
한반도 비자나무림의 식생구조
신현철
1·이광수
1·박남창
1·정수영
2*
1
국립산림과학원 남부산림연구소,
2경상대학교 농업생명과학대학 학술림
Vegetation Structure of the Torreya Nucifera Stand in Korea Hyuncheol Shin
1, Kwangsoo Lee
1, Namchang Park
1and Su Young Jung
2*
1
Southern Forest Research Center, Korea Forestry Research Institute, Jinju 660-300, Korea
2
The Research Forests, Gyeongsang National University, Jinju 660-701, Korea
요 약: 한국에서 비자나무는 남해안 및 제주도의 온난 기후대에 주로 분포하고 있으며 , 사찰과 향교 주변 등에서 주로 보호림으로 조성되어 오고 있다 . 본 연구의 주된 목적은 한반도의 대표적 비자나무 임분에서 식생구조분석을 실시함으로서 현재 비자나무 식생군락의 동태를 파악하고 이로부터 적절한 임분 보전대책을 수립하는데 있다 . 비자 나무 임분 내에서 조사된 종수는 총 148 종이었으며 , 교목층 임분에서 출현수종 28 종 , 아교목층 출현수종 38 종 , 그리
고 관목층 출현수종 82 종의 목본 관속식물이 생육하고 있는 것으로 나타났다 . 조사된 비자나무 임분에서 공통적으 로 나타난 종은 쥐똥나무 , 송악 및 마삭줄이었다 . 조사지역 모든 층위에서 비자나무가 우점종 이었으나 , 아교목층 및 관목층에서는 그 우점치가 다소 떨어져 임분 쇠퇴현상이 나타났는데 , 이러한 사실은 교목층 임분의 수관발달 및 아 교목층에서의 생장편의에 의한 결과이다 . 비자나무림의 교목층과 아교목층은 천연발아에 의해 조성된 임분이 대
부분이다 . 따라서 조사된 우리나라 비자나무림에 대한 건강한 임분을 유도하기 위해서는 도태 간벌시업을 응용함으 로서 과도한 수관경쟁의 조정이 필요할 것으로 판단되는 바이다 .
Abstract: In Korea, Torreya nucifera (L.) Siebold & Zuccarini is widely distributed in the warm temperate zone of South coastal area and Jeju island, mainly as preserved forest in the vicinity of the Buddhist temple and Confucian temple. The objective of this study is in order to develop the conservation method and comprehension of vegetation community by current vegetation structure analysis of Torreya nucifera stand. As the results, the number of surveyed species in Torreya nucifera stand were total 148 species with 28 species of tree layer, 38 species of subtree layer, and 82 species of shrub layer. The appearance of the common species were Ligustrum obtusifolium Siebold & Zucc., Hedera rhombea Bean, and Trachelospermum asiaticum Nakai var. asiaticum. All the story of the surveyed region, Torreya nucifera maintained the current dominant species, but the subtree layer and shrub layer was decreased dominant rate because of the development of the crown of tree layer and biased growth of the subtree layer. Most of the tree layer and subtree layer in Torreya nucifera stands are composed of the sprout forest. Therefore, these results suggest that in order to maintain the healthy stand it is demanded for the application of selection thinning method for reducing crown competition in Torreya nucifera forests.
Key words : warm temperature zone, dominant species, similarity index, environmental variable
서 론
한반도의 난대림은 제주도 및 남해안의 해안 도서지역 을 중심으로 주로 분포하고 있다. 하지만 대부분의 난대 수종은 그 임상이 인위적 간섭, 염해 및 태풍 등 자연재해 등에 의해 그 임분이 쇠퇴하고 있으며, 해송 및 낙엽활엽 수 등에 의해 천이과정을 거치면서 그 임분 동태가 복잡
해지고 있는 실정이다. 난대림은 그 자체로도 내륙의 임 상과 다른 독특한 자연경관을 지니고 있을 뿐만 아니라, 해안 지형특성 및 그 환경에 적응한 수종으로 구성되어 있어 생물유전자원 및 천연자원으로서도 그 가치가 높다.
특히 지구온난화로 인한 난대기후대의 북상에 따른 난대 림분포범위의 확대가 예측되며, 이에 따른 붉가시나무, 구 실잣밤나무, 황칠나무, 후박나무 등의 난대 수종에 대한 식물사회학적 연구가 활발히 수행되고 있으며(Shin et al., 1999; Park et al., 2001; Shin et al., 2001; Shin et al.,
*Corresponding author
E-mail: [email protected]
한반도 비자나무림의 식생구조 313
2002), 나아가 이들의 체계적인 관리 및 보전대책이 요구
되는 시점이다 .
난대수종 중 비자나무는 주목과 (Taxaceae) 에 속하는 상
록침엽교목이며 세계적으로 5 종이 분포하는데 중국에 2
종 , 북아메리카에 2 종 , 우리나라를 비롯한 일본에 분포하 는 종은 Torreya nucifera (L.) Siebold & Zuccarini 로 분 포지는 난대와 온대남부지방의 거주지나 사찰 , 향교주변 등의 단목 또는 집단림으로 분포하고 있으며 , 植木은 서
해안에 있어서 북위 36°, 남부는 35°10', 동해안에 있어서 는 36°30' 까지 , 1 월의 평균기온이 0oC 선까지 분포하며 백 양산에서는 산복 이하지역까지 자라고 있다 . 비자나무는 재질이 치밀하고 심재가 황색을 띄고 있는 데다 독특한 향기와 광택이 있어 바둑판 , 조각재 , 의장재 등으로 고가
에 거래되고 있으며 종자는 과거 식용기름 또는 구충제로 사용되기도 했다 . 이와 같이 비자나무는 그 효용가치에 따 른 인간의 이용 및 간섭으로 자연상태의 순림 및 그 식생 을 관찰하기 어려운 실정이다 . 따라서 본 연구는 한반도 에 분포하고 있는 비자나무 임분의 식생구조를 분석하기 위하여 층위별 식생조사를 실시하여 비자나무 숲의 임분 동태를 체계적으로 파악하기 위하여 수행되었다 . 이러한 기초자료는 한반도에서의 생태적으로 안정된 비자나무림 을 조성하고 유도하는데 필요한 정보를 제공할 뿐만 아니 라 나아가 생태적으로 균형을 이룬 비자나무 임분의 지속 적 보전 및 관리를 하는 데에도 효과적으로 응용될 수 있 을 것으로 판단되는 바이다 .
연구내용 및 방법
1. 조사지 개황
우리나라 한반도에 분포되어 있는 6 개 지역의 비자나무 림을 조사지로 선정하였다 . 이들 중 전남 장성군 북하면 낙수리 백양사의 비자림은 생육분포의 북방한계지 ( 北方 限界地 ) 로 그 가치가 인정되어 천연기념물 (153 호 ) 로 지정 되었는데 297.5 ha 의 면적에 5,000 여 그루가 자라고 있으
며 고려 고종 때 각진국사가 심은 것으로 전해진다 . 전남 화순군 춘양면 가동리 개천사의 비자나무림은 천태산 동
남쪽 중턱 1.2 ha 의 면적에 300 본정도가 생육하고 있으며
전남도 기념물 (65 호 ) 로 지정되었다 . 전남 강진군 칠량면 의 비자나무림은 계곡부의 임도변에 위치하고 있으며 , 소
면적이나 자연상태로 잘 보존되고 있고 생육도 양호하다 .
전남 고흥군 포두면 봉림리 금탑사의 비자림은 천등산 중
턱에 9.7 ha 생육하고 있으며 학술연구자원으로 천연기념
물로 지정 (239 호 ) 되어있다 . 전남 장흥군 유치면 보림사의 비자림은 가지산 중턱에 위치해 있으며 사찰림이 대부분 이다 . 북제주군 구좌읍 평대리의 비자림은 학술연구자원
으로 천연기념물 (182-2 호 ) 로 지정되었으며 44.8 ha 의 면
적에 수고 11 m, 평균 흉고직경이 73 cm 내외이며 큰 개 체는 110 cm 에 달한다 .
2. 식생조사 및 통계처리
조사지역을 대상으로 2005 년도 임목의 생육기간동안 비 자나무 군락지에서 10 m×10 m 크기의 총 80 개 방형구를 설치하고 , 식생요인 및 국소지형 , 고도 , 사면향 , 경사도 , 토 양의 화학성 등의 환경요인 자료를 수집하였다 . 식생조사
에서 각 층위별 구분은 일반적으로 수고 8 m 이상을 교목
층 , 2 m 이하를 관목층 , 그 중간범위를 아교목층으로 하
였다 . 비자나무 임분에서의 Classification 및 Ordination 분 석은 다음의 방법에 의하여 수행되었다 .
식생조사 자료로 부터 각 종의 합성치 Xij를 다음과 같 이 구하였다 ; Xij=(d
ij+D
ij)/2. 이때 , X
ij는 j 조사구에서 종 i
=(d
ij+D
ij)/2. 이때 , X
ij는 j 조사구에서 종 i
의 합성치이며 , dij는 상대밀도 , Dij는 상대피도를 나타낸다 .
는 상대피도를 나타낸다 .
합성치 Xij를 가지고 각 조사구에 따른 종조성을 나타내는
식생 자료 matrix 를 작성하였으며 , 또한 야외 조사와 실험
실 측정 결과 얻어진 환경 요인들을 이용하여 환경요인 자료 matrix 를 작성하였다 .
Classification 은 Hill(1979) 의 TWINSPAN(two-way indicator species analysis) 을 이용하였으며 , 얻어진 자료는
0%, 2%, 5%, 10%, 20% 의 cut level 이 사용되었다 . 각 조
사구에서 20% 이상의 중요치를 가지는 종은 그 조사구의
우점종으로 간주되었다 .
Ordination 은 DCA(detrended correspondence analysis)
의 확장인 DCCA(detrended canonical correspondence analysis) 를 사용하였으며 (Hill 1979) Ter Braak(1987) 의
CANOCO program 을 이용하였다 . 결과 및 고찰
1. 비자나무림의 수종구성
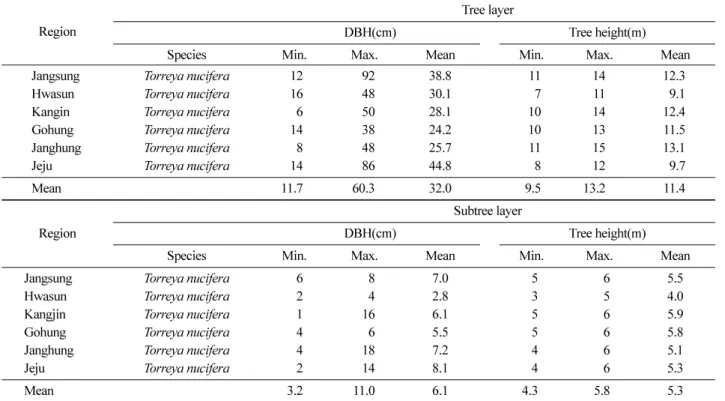
우리나라 비자나무림 중에서 조사지역으로 선정된 임분 의 교목층 및 아교목층 생장량 조사결과는 Table 1 과 같다 .
우리나라 비자나무림을 대상으로 선정된 조사임분에서 교목층의 평균수고는 11.4 m, 평균흉고직경급은 32 cm 로 나타났으며 , 평균수고가 가장 높게 나타난 지역은 장흥지 역의 비자나무림분이었다 (Table 1). 그리고 아교목층 비자 나무림의 평균수고는 11.4 m 이었으며 , 평균흉고직경급은 32 cm 로 조사되었다 .
전남 장성군 북하면 낙수리 백양사의 비자나무림을 포 함한 6 개 지역의 비자나무림의 층위별 식생조사에서 , 출 현된 각 수종에 대한 중요치 (Importance value) 를 요약한 결과는 Table 22 와 같다 .
Table 2 는 비자나무림의 층위에 따른 수종별 중요치를
분석한 결과로서 , 비자나무림의 교목층 중요치는 비자나
314 韓國林學會誌 제 99 권 제 3 호 (2010)
무가 전 지역에서 우점하고 있었다. 특히 장흥지역은 82.4 로 조사된 6개 지역 임분 중 가장 높은 값을 보였는데 이 는 출현종 28종 중 교목층 출현종의 수가 4종으로 가장 단 순하였을 뿐만 아니라 임야 소유주가 보림사 사찰림으로 비자나무를 위주로 집중관리 하였기 때문으로 추정된다.
지역별로 비자나무림에서 중요치가 높은 종은 장성지 역에 내장단풍, 팽나무가 화순지역은 굴참나무, 소나무, 팽나무, 비목 등이 비교적 높은 값을 보였으며 강진 지역 은 굴참나무, 졸참나무, 서어나무가 고흥 지역은 서어나무, 비목, 느티나무가 장흥 지역은 졸참나무가 제주지역은 팽 나무, 예덕나무가 높은 값을 나타내었다. 출현종의 수가 전 지역 모두 다른 종에 비하여 전반적으로 낮았는데 비 자나무의 수관발달이 왕성하여 수관폭이 넓을 뿐만 아니 라 종실을 식용이나 약용으로 이용하기 위하여 꾸준히 집 약관리를 하였기 때문으로 사료된다.
비자나무림의 아교목층 중요치는 제주지역을 제외한 전 지역에서 비자나무의 중요치가 비교적 높게 나타나고 있는 데 그중 강진, 장흥에서는 우점하고 있으나 장성, 화순, 고 흥지역에서는 단풍나무나 동백나무 등과 경쟁으로 순화를 이루고 있었다. 제주지역은 비자나무가 전혀 출현하지 않 고 있으며 천선과나무, 단풍나무 등의 낙엽활엽수와 생달 나무, 후박나무 등의 상록활엽수가 높은 값을 나타내었다.
비자나무림은 관목층에서 해당 조사지역 모두 출현하 였으며, 중요치는 강진지역이 가장 높은 값을 나타내었고 제주지역이 가장 낮은 값을 보이고 있으며 제주지역의 아 교목층에서는 출현하지 않는 것으로 보아 교목층 울폐도
가 높아 수광량 부족 등의 원인으로 관목층에 발생된 치 수가 더 이상 자라지 못하고 고사하는 것으로 보인다. 이 러한 현상은 제주지역 외 지역에서도 대다수의 치수가 고 사되고 극히 일부의 개체만이 성장하는 것으로 추정된다.
출현종은 모두 82종 이였는데 장흥지역이 20종으로 종 구성이 가장 단순하였고 화순지역이 43종으로 비교적 다 양한 것으로 나타났다.
쥐똥나무, 찔레꽃, 송악, 마삭줄 등 4종은 전 지역에서 출현하는 것으로 보아 비자나무와 생육 지위를 같이하는 종으로 보이며, 장성이나 하순지역은 내장단풍, 단풍나무 의 중요치가 아교목층 및 교목층에 이어 높은 값을 나타 내어 비자나무와 경합을 보이거나 우점종으로 발달될 수 있을 것으로 예측된다. 강진지역은 사스레피나무나 동백 나무와 같은 상록활엽수가 비교적 높은 값을 보이고 있으 나 비자나무와는 차이를 보이고 있다. 송악, 마삭줄 등이 높은 값을 나타내고 있으나 이러한 종은 지면을 피복하고 있는 정도였다. 고흥지역은 국수나무가 높은 값을 보이고 있으나 이러한 종들은 관목성이므로 관목층을 벗어나지 못할 것이며 서어나무나 비목나무 등이 비자나무와 수관 경쟁을 벌일 것으로 예측된다. 장흥지역은 사찰림으로 비 자나무 수하에 차밭을 조성하여 집중관리를 하고있을 뿐 만 아니라 조릿대 등이 높은 값을 나타내어 다른 종의 입 지를 약화시키는 것으로 나타났다. 제주지역은 상산, 남오 미자, 생달나무, 송악 등이 높은 값을 보이고 있으나 이러 한 종들은 관목류나 만경류로 이들이 제거되어질 경우 생 달나무의 입지가 점차 확대될 것으로 추정된다.
Table 1. Tree growth measurements of Torreya nucifera in tree layer and subtree layer by each study region.
Region Tree layer
DBH(cm)
Tree height(m)
Species Min. Max. Mean Min. Max. Mean
Jangsung Torreya nucifera 12 92 38.8 11 14 12.3
Hwasun Torreya nucifera 16 48 30.1 7 11 9.1
Kangin Torreya nucifera 6 50 28.1 10 14 12.4
Gohung Torreya nucifera 14 38 24.2 10 13 11.5
Janghung Torreya nucifera 8 48 25.7 11 15 13.1
Jeju Torreya nucifera 14 86 44.8 8 12 9.7
Mean 11.7 60.3 32.0 9.5 13.2 11.4
Region Subtree layer
DBH(cm)
Tree height(m)
Species Min. Max. Mean Min. Max. Mean
Jangsung Torreya nucifera 6 8 7.0 5 6 5.5
Hwasun Torreya nucifera 2 4 2.8 3 5 4.0
Kangjin Torreya nucifera 1 16 6.1 5 6 5.9
Gohung Torreya nucifera 4 6 5.5 5 6 5.8
Janghung Torreya nucifera 4 18 7.2 4 6 5.1
Jeju Torreya nucifera 2 14 8.1 4 6 5.3
Mean 3.2 11.0 6.1 4.3 5.8 5.3
한 반 도 비 자 나 무 림 의 식 생 구 조 3 1 5
Ta ble 2. Importance val ues of each species occurr ed at the tr ee lay er, subt ree lay er, and shrub lay er in
Torreya nuciferafor est s. Sp ec ies (S cine tific nam e) Jangsung Hw asu n Ka ngjin Gohung
Janghung Jeju To tal Tr ee l. Subtr ee l. Sh rub l. Tr ee l. Su btr ee l. Shr ub l. Tr ee l. Subt ree l. Shr ub l. Tr ee l Subtr ee l. Shr ub l. Tr ee l. Subtr ee l. Shr ub l. Tr ee l. Subt ree l. Sh rub l. Tr ee l. Subtr ee l. Sh rub l. To rre ya nuci fer a 51. 5 21. 6 7.6
53. 9 9.0 4.0
48.0 32.0 10.5
65. 3 21. 9 4.2
82. 4 48. 0 7.7
63. 5
3.1
60. 8 22. 1 6.1 Ac er pal matum var . nakai i 18. 1 35. 3 12. 5
3.0 5.9 2.1 Ac er pal matum
4.1 13. 6 5.3
6.1 3.0
4.9
3.3 1.9
1.7 13.9 2.5
1.0 7.0 2.1 Qu erc us va ria bil is
7.7 4.9 14.4
3.7 0.8 Que rcus se rrat a
1.3 3.2 2.3
11 .1 3.4
11 .3 13. 8
3.9 3.4 0.4 Carpinus lax iflora 3.6 9.1 2.7
1.4
9.1 7.2
12. 2 10 .8 5.7
4.1 4.5 1.6 Cel tis si nensi s 8.2 11 .9
6.6
9.3
11 .5
4.4 3.5 Mall otus japonic us
1.6 2.0
9.8 0.0 5.2
1.6 0.3 1.2 Pi nus d ensi flora
7.6
1.3 Linde ra ery thr oc arpa 1.5
0.8
6.4 6.7 4.6
6.6 4.9 5.8
1.9
1.8
2.4 2.2 2.2 Pl aty cary a s trobi lace a
2.9
3.3
3.6
1.6 Ac er pal matum var . pilos um
1.6
3.8
0.3 0.6 Pr un us sar genti i
3.3
1.7
0.8 Cornus cont ro ve rsa 2.9 2.6 2.3
8.3
3.9 4.9
1.1 2.6 0.4 Castanopsi s cuspi data var . siebol dii
Ze lkov a serr ata 1.4 3.4 3.5
4.0 7.4 4.3
8.3
5.3 8.8 3.8
3.3
1.8 5.2 1.9 Eus caphis j aponi ca
3.1 10.5
0.5 1.8 Sty rax japon ica 2.6 5.8 0.0
1.3 12. 3
5.7 11 .2
4.9
3.5
1.6 6.3 Zanthox ylum ail anthoide s
12.7
25 .6
1.3 4.9
0.2 7.2 Cinnam omum japonic um
4.9 15.2
0.8 2.5 Gl edit sia japoni ca var . ste nocar pa
2.8
0.5 Al bizzi a j uli brissi n
1.4
5.6
0.0 7.3
1.3
0.6
1.4 1.2 0.1 Qu erc us s ali cin a
0.0
0.0 0.0
1.7
0.6
0.3 0.1 Que rcus ac utis sim a
3.0 6.6
0.5 1.1 Carpinus ts chonosk ii
2.7
0.5 Que rcus al iena 6.2
1.0 Mel iosm a my riantha 2.5 5.2
1.6
4.3 2.5
0.4 1.9 0.4 Di ospyr os ka ki 1.5
0.3 0.0 0.0 Machi lus thunbe rg ii
1.3 10.5 1.9
0.2 1.8 0.3 Fi cus e rec ta
21.9
3.6 0.0 Cornus kousa
5.2 2.3
1.5
4.3 1.7
1.8
1.9 0.9 Cornus walt eri
4.9 0.0
0.8 0.0 Coryl us het erophy lla va r. thunb ergi i
4.9 1.4
3.4
2.5
1.4 0.6 Staphy lea bum alda
1.6 3.0
1.3
0.3 0.7 Pr un us pe rsic a
2.6
0.4 Maack ia amur ensi s
8.7
1.5 Rhus t ric hocarpa
2.6
8.1
3.2
9.6
1.4
4.1 Eur ya japonic a
8.1 6.7
4.2
1.4 1.8 Viburnum awabuk i
3.6
0.6 Carpinus c or eana
2.1
0.4 Call icar pa j apo nica
4.0
4.9
2.2 6.6
0.4 2.6
3 1 6 韓 國 林 學 會 誌 제 9 9 권 제 3 호 ( 2 0 1 0 )
Table 2. Continued.
Spe cie s (S cinet ific na me ) Jangsung Hw asun Ka ngj in Gohung
Janghung Jej u To tal Tr ee l. Subtr ee l. Shr ub l. Tr ee l. Subt ree l. Sh rub l. Tr ee l. Subtr ee l.S hrub l. Tr ee l Subtr ee l. Shr ub l. Tr ee l. Subt ree l. Sh rub l. Tr ee l .S ubtr ee l. Shr ub l. Tr ee l. Subtr ee l. Shr ub l. Cor nus m acr oph yll a
1.8
0.3 Cal lic arpa mol lis
2.3
3.0
2.9
1.4 6.4
0.2 2.4 Vibur num er os um
2.2
0.4 Sy mpl ocos panic ulata
2.5
0.4 Qu erc us dent ata
1.8
0.3 M aa cki a f aurie i
2.8
0.5 Vibur num er os um
3.7
4.0
4.5
1.7
1.9
2.6 Se curi nega suff ruti cosa
0.7
0.1 Ste phanandra inc isa
1.0
3.9
3.7
9.9
3.1 Cudr an ia t ric uspidat a
0.7
0.1 Kads ura j aponica
0.0
7.0
1.2 Ac tini dia ar guta
2.7
1.3
0.7 Al angi um plat an ifol ium
1.9
0.3 Pa rthe noci ssus tri cus pidata
2.0
4.6
1.1 Cam ell ia j apon ica
5.5
4.1
1.8
1.9 Ar ali a e lat a
0.7
0.1 Wi stari a f lori bun da
1.7
0.3 Sty rax japonic a
3.3
1.7
3.8
1.5 Tr ac hel osper mum asi ati cum va r. inte rm ed ium
7.3
3.3
10. 2
5.0
10. 3
5.6
6.9 Stauntoni a he xaphy lla
2.5
0.4 Fi cu s nipponic a
1.3
0.2 Li nder a gl auca
0.7
0.1 He de ra r hom bea
9.3
4.3
8.0
3.4
8.8
6.7
6.7 Ar disia japonic a
4.3
3.3
1.3 Zanthoxy lum pl anispi num
1.5
4.5
1.0 El ae agnus mac rophyl la
3.0
3.1
1.0 Euonym us glabra
1.5
0.3 Cl em ati s api ifo lia
0.8
2.1
0.5 Zanthoxy lum sc hini fol ium
4.7
1.4
5.3
3.4
1.9
2.8 Or ixa j aponic a
4.0
0.7
3.4
10.5
3.1 Li nder a obt usiloba
5.0
2.7
6.1
5.0
4.7
3.9 Ci nnamom um japoni cum
6.4
1.1 Pourthi aea vi llosa
1.6
2.5
2.5
1.1 Ak eb ia quinat a
3.0
2.0
1.6
1.9
1.4 Kal opanax pic tus
0.7
0.1 Sasa bor ea lis
7.1
6.4
15. 3
4.8 Li gust rum obtusi foli um
5.7
4.0
4.6
1.7
3.8
1.3
3.5 Rhododendr on m ucr onul atum
1.4
0.2 Ro sa mul tiflora
0.8
2.0
3.1
1.7
2.9
1.9
2.0 Th ea sine nsis
1.5
21. 5
3.8 Fi cu s e rec ta
3.1
0.5 Sm ilax china
3.3
1.5
1.7
1.3
1.3
한 반 도 비 자 나 무 림 의 식 생 구 조 3 1 7
Table 2. Continued.
Sp ec ies (S cine tif ic nam e) Jangsung Hw asun Ka ngji n Gohung
Janghung Jeju To tal Tr ee l. Subt ree l. Shr ub l. Tr ee l. Subtr ee l. Shr ub l. Tr ee l. Subt ree l. Shr ub l. Tr ee l Subt ree l. Sh rub l. Tr ee l. Subtr ee l. Shr ub l. Tr ee l. Subt ree l. Sh rub l. Tr ee l. Subtr ee l. Sh rub l. Pu erari a t hunber giana
1.0
0.2 Le mm aphyl lum m icr ophyl lum
6.4
1.1 Ce ltis s ine ns is
2.3
2.7
1.9
1.1 Ri bes fa sci culat um var . ch ine nse
3.7
1.0
0.8 Hy drangea pet iol aris
1.3
0.2 Eu on ym us fo rtu ne i var . radic ans
1.5
2.9
1.3
0.9 Eu on ym us ox yphy llus
0.6
0.1 Juni perus ri gida
0.7
1.5
0.4 Le s pedez a c yrt obotry a
1.5
0.3 Po nc irus t rif oliat a
0.8
0.1 Eu on ym us al atus fo r. cil iato- de ntatus
0.8
1.5
5.0
1.9
1.5 Viburnum di latat um
0.7
0.1 Ac er m ono
1.4
1.7
0.5 Br oussone tia kaz inoki
1.4
0.2 Ru bu s c or eanus
0.7
0.1 Rh us ch inen sis
0.7
0.1 Ru bu s c ratae gif oli us
1.4
1.7
0.5 Le s pedez a m axi mowi czi i
0.7
0.1 Sapium japon icum
2.5
1.9
0.7 Rh odode ndr on schl ippe n bac hii
1.7
0.3 We igel a subs essi lis
1.7
0.3 Fr axi nus s iebol dina
0.9
0.1 Eu on ym us al atus
2.5
0.4 Ot hers
2.1
0.4 To tal 100 100 100
100 10 0 100
100 100 100
100 100 100
10 0 100 100
10 0 100 100
100 100 100
318 韓國林學會誌 제 99 권 제 3 호 (2010)
Table 3 에서 , 각 지역별 비자나무림의 중요치는 교목층
전 지역에서 우점하고 있으며 특히 장흥 보림사 지역은
80 을 상회하는 가장 높은 값을 보였다 . 관찰된 교목층 출
현 종 28 종 중 4 종만이 출현하고 있었는데 , 이러한 사실
은 이 지역이 사찰림으로 비자나무 위주로 집중 관리되어 온 결과 그 출현종이 단순하였기 때문으로 사료된다 . 강 진 칠량 지역은 비자나무가 산재되어 있는데 48 로 가장 낮은 값을 보이고 있지만 아교목층 및 관목층에서 상대적 으로 높은 값을 나타내어 조사지 중 비자나무림으로의 천 이가 활발하게 이루어지고 있는 것으로 보인다 . 제주 지 역은 아교목층이 형성되지 못하고 있는데 교목층 울폐도 가 높아 관목층에서 발아된 비자나무 실생묘가 수광량 부 족으로 더 이상 자라지 못하는 것으로 추정된다 . 아교목 층은 각 지역 간의 차이는 있으나 비교적 높은 값을 보이 고 있지만 교목층 수관의 발달로 관목층에서 발달된 치수 가 교목층 가지부분과 交互되어 頂芽부분이 절단되거나 빈 공간으로 편기되어 자라며 , 벌채에 의한 맹아지로부터 발달한 개체가 상당수 관찰되었다 . 건전한 임분구성을 위 하여 상당량의 간벌이 실시되어야 할 것으로 사료된다 .
출현종수는 전지역 모두 다른 수종에 비하여 전반적으 로 떨어지는 것으로 나타났는데 비자나무의 수관발달이 왕성하여 수관폭이 매우 넓게 차지할 뿐만 아니라 자연낙 지도 잘 이루어지지 않는 것으로 보이며 , 종실을 이용하 기 위하여 꾸준히 집약관리를 하였기 때문으로 사료된다 .
2. 비자나무림의 종다양성, 우점도 및 분포특성 지역별 출현종수 (S), 개체수 (N), 종다양도 (H'), 최대종다
양도 (H' max), 균재도 (J'), 우점도 (1-J') 에 의한 비자나무림 의 식생 특성을 분석한 결과는 Table 4 과 같다 .
화순 개천사 지역의 종다양도 , 최대종다양도 , 균재도 등 이 가장 높게 나타나 가장 건전한 임분을 형성하고 있는 데 어떤 특정한 종이 우점하지 않고 출현 종간에 고른 분 포를 보여 군락이 가장 안정된 것으로 사료된다 . 반면 장 흥 보림사 지역이 가장 낮은 값을 보이고 있는데 비자나 무 임분의 수하에 차나무 재배로 다른 종의 이입을 억제 하기 때문에 균재도 등이 낮은 것으로 보이며 전반적으로 종다양도 등이 다른 수종에 비하여 상대적으로 낮은 값을 보였다 .
Table 5 에서 , 각 지역별 , 상·아교목층에 대한 산포 상 태를 알아보기 위하여 밀도 , 비자나무 한 그루가 차지하 는 면적 및 개체간의 거리등을 나타내었는데 교목층에서 는 강진 칠량지역의 밀도가 가장 높게 나타나 본당 평균 면적은 10.1 m
2, 평균거리 3.2 m 로 나타났으며 장흥 보림
사지역의 밀도가 가장 낮아 본당 평균면적 15.4 m2, 평균
거리 3.9 m 로 나타났다 .
아교목층은 교목층에 비하여 밀도가 더욱 낮게 나타났 으며 , 그 중에서 장성 백양사지역이 가장 낮아 본당 평균
면적은 31.3 m2, 평균거리 5.6 m 이었으며 강진 칠량지역 이 11 m
2, 평균거리 3.3 m 로 가장 높게 나타났다 . 따라서 이러한 임분특성을 고려해 볼 때 , 생태적으로 균형이 잡 힌 비자나무림의 식생구조를 갖추기 위해서는 아교목층 에 있어서의 비자나무 발달을 유도하는 것이 우선 요구된 다 . 적절한 시업적 접근 방법으로는 교목층의 미래목을 선 정한후 나머지 형질 불량목 등에 대해서 점차적 간벌시업
Table 3. Summary of the importance value of Torreya nucifera species in each study region.
Study region I.V.* of Torreya nucifera Number of other species
Tree l.** Subtree l. Shrub l. Tree l. Subtree l. Shrub l.
Jangsung 51.5 21.6 7.6 11 10 28
Hwasun 53.9 9.0 4.0 12 16 43
Kangjin 48.0 32.0 10.5 8 12 26
Gohung 65.3 21.9 4.2 8 12 30
Janghung 82.4 48.0 7.7 4 9 20
Jeju 63.5 - 3.1 10 14 32
*I.V.; Importance value, **Tree l. means tree layer (T1) in tree height <8 m, Subtree .l. (T2) is subtree layer from 2 m to 8 m in tree height, and Shrub l. (S) is from 0.4 m to 2 m.
Table 4. Species diversity indices of Torreya nucifera forests in each study region.
Region No. of species
(S) No. of individuals
(N) Species diversity
(H') Maximum H'
(H' max) Evenness
(J') Dominance
(1-J')
Jangsung 13 108 0.739 1.113 0.664 0.336
Hwasun 20 130 1.092 1.301 0.839 0.161
Kangjin 16 190 0.946 1.204 0.786 0.214
Gohung 15 116 0.712 1.176 0.605 0.395
Janghung 11 118 0.519 1.041 0.499 0.501
Jeju 20 148 0.992 1.301 0.762 0.238
한반도 비자나무림의 식생구조 319
이 필요하다. 특히 일시적 교목층임분의 간벌은 임분내 환 경을 급변시켜 오히려 임분의 생태적 안정화에 악영향을 미칠 수 있으므로 점진적 교목층 불량목의 제거가 바람직 할 것으로 사료되는 바이다.
Table 6에서, 지역별, 층위별 비자나무림의 전 기저면적 을 산출하고 비자나무가 차지하는 비율을 분석한 결과이 다. 6개 비자나무 조사지역 중 기저면적이 가장 높게 나타 난 곳은 제주 지역 비자나무 임분이었으며, 이 임분의 직 경급 또한 대경목으로 구성되어 있어 비자나무 순림이 생 태적으로 비교적 안정된 상태를 유지하고 있는 것으로 나 타났다. 교목층 임분 조사에서 제주지역 다음으로 장흥지 역 역시 비자나무 기저면적 비율이 90%이상으로 높게 나 타나 비자나무 순림에 도달하고 있음을 알 수 있었다. 비 자나무 임분 조사지역 중에서 강진지역은 68%로 참나무 류와 장성지역은 72%로 단풍나무류와 혼효 상태로 있는 것으로 보인다.
아교목층에 분포하고 있는 비자나무의 기저면적은 교 목층 비자나무의 기저면적에 비하여 현저히 감소하고 있 는데 제주지역은 비자나무가 전혀 출현하지 않고 있을 뿐 만 아니라 화순지역도 9%로 매우 낮은 값을 나타났는데 이러한 지역은 교목층 울폐도가 너무 높아 아교목층이 상 대적으로 빈약하게 발달하고 있는 것으로 나타났다. 따라 서 건전한 임분형성을 조성하기 위하여 교목층 임분에 대 한 간벌 특히, 참나무류, 단풍나무류 등의 여타 침입수종 의 간벌시업을 적당량 실시하여야 할 것으로 사료된다.
Lee et al.(2009)는 특정 수종으로 구성된 임분을 조성하고 자 할 때 이와 경쟁관계에 있는 종을 제어함으로써 목적 임분으로의 유도가 가능하다고하여 본 연구의 간벌시업 제안과도 유사한 경향을 보이고 있다.
3. 수종간 상관 및 지역별 유사도지수 1) 비자나무림 내 출현 수종 간 상관관계
비자나무림에 출현하는 수종들 간의 상관관계 특성을 파악하기 위하여 상·아교목층의 목본식물 중 중요치가 높은 16종을 선정하였다. 이들의 출현과 비 출현을 바탕 으로 Agenew(1961)의 방법에 따라 2×2 분할표에 의한 Chi-square(x
2) 값으로 각 수종간의 종 상관관계 분석을 실 시한 결과는 Figure 1과 같다.
Figure 1에서, +는 양의 상관관계를 나타내는 것으로 생 육공간을 공동으로 소유하는 수종 군을 의미하며, −는 음 의 상관으로 서로 다른 서식처를 가진 수종 군을 의미한 다. 비자나무림에 나타난 수종은 25쌍 간에 상관관계가 유 의수준 0.05에서 인정되었다. 굴참나무를 중심으로 느티 나무, 동백나무, 때죽나무, 산벚나무 등과 양의 상관을 보 이고 있으며, 서어나무를 중심으로 층층나무, 느티나무, 비목나무, 동백나무 등과 양의 상관을, 그리고 자귀나무는 개옻나무 및 졸참나무와 양의 상관을 보여 이러한 종들과 서로 생육공간을 공유하고 있는 것으로 판단된다. Lee et al.(2009)는 특정 수종으로 구성된 임분을 조성하고자 할 때 이와 상호 경쟁관계에 있는 종을 제어함으로써 목적임
Table 5. Density, mean acreage, mean distance among individuals of Torreya nucifera species in each investigated area.
Region Tree layer Subtree layer
Density
(trees/m
2) Mean acreage
(m
2/trees) Mean distance
among individuals (m) Density
(trees/m
2) Mean acreage
(m
2/trees) Mean distance among individuals (m)
Jangsung 0.076 13.158 3.627 0.032 31.250 5.590
Hwasun 0.075 13.333 3.651 0.055 18.182 4.264
Kangjin 0.099 10.101 3.178 0.091 10.989 3.315
Gohung 0.079 12.658 3.558 0.037 27.027 5.199
Janghung 0.065 15.385 3.922 0.053 18.868 4.340
Jeju 0.076 15.158 3.627 0.072 13.889 3.727
Mean 0.088 12.939 3.594 0.057 21.264 4.540
Table 6. Percentage of basal areas of the T. nucifera for total basal area.
Region Tree layer Subtree layer
Total
(cm
2) T. nucifera
(cm
2) Percentage
(%) Total
(cm
2) T. nucifera
(cm
2) Percentage
Jangsung 91,285 65,773 72 428 156 (%) 36
Hwasun 34,541 28,450 82 393 35 9
Kangjin 47,678 32,367 68 1,811 943 52
Gohung 33,018 28,712 87 1,250 194 16
Janghung 40,035 37,053 92 4,769 1,144 24
Jeju 156,351 149,310 95 835 - -
Mean 402,908 341,665 85 0 9,486 2,472 26
320 韓國林學會誌 제99권 제3호 (2010)
분으로의 유도가 가능하다고 하여 본 연구에서도 마찬가 지로 비자나무림에 나타난 수종간의 경쟁관계 즉, 양 혹 은 음의 상관관계(Figure 1)를 적절히 이용하거나 조정함 으로써 생태적으로 안정된 비자나무림으로의 유도가 가 능할 것으로 판단된다.
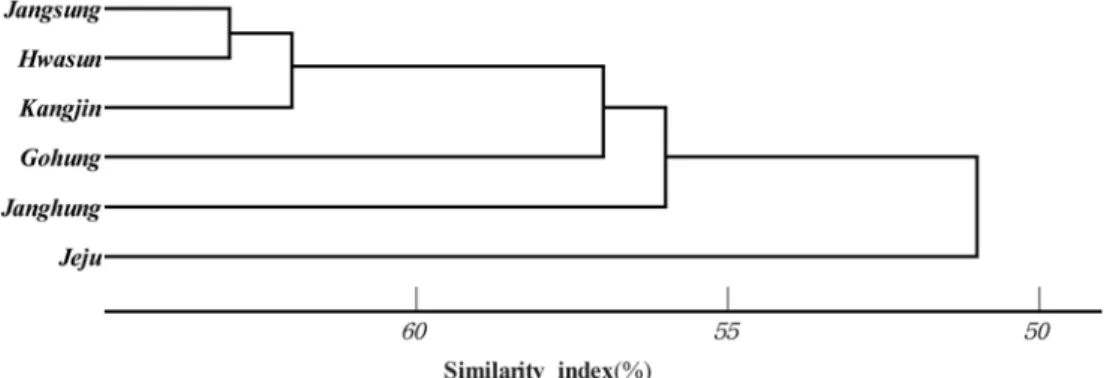
2) 비자나무림의 지역간 유사도 지수
각 조사 지역에 구성종의 유사한 정도를 알아보기 위하 여 Whittaker의 유사도지수식을 적용하여 비자나무림의 지역별 유집(Cluster)분석을 실시한 결과는 Figure 2와 같 다. 유사도 60에서 강진, 고흥, 장흥 지역을 1개 류집으로, 장성, 화순 지역을 1개 유집으로 제주 지역을 1개 유집으 로 하여 모두 3개 유집으로 구분되었다. Shin et al.(1998) 의 우리나무 주목림에 있어서 지역 간 구성종이 구분 되 는 것은 서로 인접한 지역의 온도, 습도, 강수량, 지형, 모
암 등의 입지 및 환경요인의 차이에 기인된 것이라 보고 한 바 있다. 즉, 특정 지역의 서식환경특성에 더욱 적합한 생육특성을 가진 식생이 타 식생에 비하여 그 임분을 구 성하는데 상대적으로 유리한 경쟁력을 가지고 있기 때문 에 결과적으로 지역 간 구성종의 차이가 발생하는 것을 알 수 있다. 따라서 본 연구에서 우리나라 비자나무림의 구성종에 의하여 지역별 비자나무림 특성을 구분한 결과 3개의 유집으로 나타난 결과는 Shin 등(1998)이 우리나라 주목림의 구성종이 지역에 따라 다르게 나타난 것이 입지 및 환경요인의 차이에 기인하였다는 결과와 유사한 경향 으로 해석할 수 있을 것이다.
4. TWINSPAN에 의한 군락분류
매목조사에서 얻은 정량적 측정치를 이용하여 비자나 무군락을 TWINSPAN으로 분석한 결과(Figure 3), 제 1축
Figure 1. Correlation among the importance value of the major woody plant species in Torreya nucifera stands.
Figure 2. Cluster dendrogram for the study area composed of Torreya nucifera stands forests.
한반도 비자나무림의 식생구조 321
에서 팽나무와 층층나무 그룹과 서어나무와 굴참나무 그 룹이 분리되고, 이들은 다시 제 2축에서 팽나무, 층층나무, 서어나무, 굴참나무로 구분되었다. 그리고 서어나무는 다 시 서어나무와 졸참나무로 구분되었다. 즉 비자나무-팽나 무군락, 비자나무-층층나무군락, 비자나무-서어나무군락, 비자나무-졸참나무군락, 비자나무-굴참나무군락으로 분류 되었다. Lee et al.(2009)는 서울시 소나무림 보전을 위한 관리대책으로 소나무림과 생태적 지위가 유사한 수종을 시업을 통하여 제거하고 관목층에서 이들 종의 보완이 필 요하다고 하였는데, 이는 지속적인 비자나무림 유지를 위 하여 각 계층에서 비자나무림과 경쟁단계에 있는 수종의 제거와 관목층 비자나무 치수의 보완이 필요하다라는 본 연구결과와 일치하는 보전관리 대책이다.
5. DCCA 방법에 의한 환경구배 분석
효과적인 ordination 분석을 위하여 1차 ordination을 실시 한 후, outliers을 제거한 55개 plot를 중심으로 ordination을 실시한 결과는 Figure 4와 같다. Figure 4은 TWINSPAN 에 의하여 분류된 5개 군락을 DCCA Ordination한 결과의 최초 1, 2 축에 의한 I/II 평면상에 나타낸 것이다.
비자나무 군락은 Figure 4에서 보는 바와 같이 13개의
환경요인에 따라 분포하고 있으며, 이들 환경요인들과 DCCA ordination 결과에 의한 제1, 제2축과 상관관계를 살펴보면 Table 7과 같다. 비자나무 군락 내 환경 요인들 이 군집의 분포와 깊은 상관관계가 발생하였으며, 제1축 에서는 해발고도와 사면향이 상관관계가 높으며, 제2축에 서는 해발고도와 경사가 높은 상관관계를 보여주고 있다.
제1축과 제2축에서 상관관계가 가장 높은 것을 기준으 로 볼 때, 군집의 분포에 영향을 미치는 중요한 환경요인 은 사면 향과 경사도로 나타났다. 사면 향과 경사도 요인 은 임분 내에서 고도와 수분에 영향을 미쳐 간접적으로 군집분포에 영향을 미치는 것으로 사료되며, 이러한 결과 는 Song et al.(1990)의 결과에서도 군집분포에 유사한 특 성을 보이는 환경요인인 것으로 드러나 본 연구결과와 유 사한 경향을 보였다.
또한 주요 군집들과 해발고도와 사면향 간의 관계 특성 을 보면, 해발고도가 높은 곳에서 낮은 곳으로 비자나무- 굴참나무군락, 비자나무-졸참나무군락, 비자나무-서어나 무군락, 비자나무-층층나무군락, 비자나무-팽나무군락 순 으로 분포하고 있다(Figure 4). 그리고 남사면에서 서巢욉 북사면으로 비자나무-굴참나무군락, 비자나무-졸참나무군 락, 비자나무-서어나무군락, 비자나무-층층나무군락, 비자 나무-팽나무군락 순으로 분포하고 있다.
또한 주요 군집들과 환경과의 상관관계로 보면, 비자나 무-굴참나무군락은 해발고도가 높은 남사면에 주로 분포 하고 있고, 비자나무-졸참나무군락은 해발고도는 높은 곳 에서 중간까지인 남사면에 주로 분포하고 있다(Figure 4).
Figure 3. The pathway of sub-division into groupings of Torreya nucifera community vegetation using TWINSPAN.
Figure 4. Torreya nucifera community vegetation data : DCCA (detrecded canonical correspondence analysis) ordination diagram with plots (
○,
●,
★,
△,
▲) and environmental variables (arrow).
Table 7. Torreya nucifera community vegetation data from Figure 2: canonical coefficients and the inter set correlation of environmental variables with the first two axes of detrended canonical correspondence analysis. For a description of variables, see Figure 2 legend.
Axis Canonical coefficients Correlation coefficients
Variables 1 2 1 2
Topography 0.073 0.036 -0.238 -0.230 Elevation -0.118 0.114 -0.444** -0.606**
Slope aspect 0.134 0.026 -0.460** -0.001 Slope degree 0.143 0.021 -0.059 -0.363**
pH 0.282 -0.016 -0.073 -0.123
Organic matter 0.042 0.054 -0.066 -0.224 Total nitrogen -0.044 -0.013 -0.062 -0.138
P
2O
5-0.190 0.017 -0.070 -0.145
C.E.C. -0.049 0.075 -0.070 -0.145
K -0.082 -0.014 -0.045 -0.164
Na 0.165 -0.095 -0.082 -0.117
Ca -0.036 0.016 -0.058 -0.087
Mg -0.122 0.003 -0.001 -0.107
Eigenvalue 0.219 0.060
**, *: significantly, correlation coefficient at a=0.01, 0.05, respec-
tively
322 韓國林學會誌 제 99 권 제 3 호 (2010)
비자나무-서어나무군락, 비자나무-층층나무군락, 비자나 무-팽나무군락은 고도가 낮은 북사면에 주로 분포하고 있 으나, 비자나무-서어나무군락이 상대적으로 급경사지에서, 비자나무-층층나무군락, 비자나무-팽나무군락은 비자나무 -서어나무군락보다 다소 완경사지에 분포하고 있는 것으 로 나타났다.
결 론
조사지역의 비자나무림 내 출현 종수는 교목층 28종, 아 교목층 38종, 관목층 82종으로 나타났으며, 전체 조사 지 역에서 공통으로 출현하고 있는 종은 쥐똥나무, 송악, 마 삭줄 등 5종이었다. 각 지역별 전 층에서는 비자나무가 우 점하고 있어 현재의 임상이 계속될 것으로 보이나 교목층 수관의 발달로 인하여 아교목층 및 관목층의 점유도가 떨 어지고 특히 아교목층 수관이 한곳으로 편기되어 수형이 불량하며 아교목층 및 교목층을 형성하고 있는 개체 중 상당량의 비자나무림은 맹아림으로 형성되어 있어 건전 한 임분을 유도하기 위하여 적정량의 도태간벌이 필요한 것으로 생각된다. 또한 비자나무림의 각 층위별 생물종다 양성을 보전함과 동시에 비자나무림의 안정적인 생태적 지위를 유지하기 위해서 각 층위별 비자나무림 조성이 필 요하다. 특히 비자나무림의 교목층과 아교목충은 천연발 아에 의해 조성된 임분이 대부분인데 이들의 임분밀도 조 절에 따른 임분내 광수분 조정을 통한 생태적 복원 및 이 에 따른 지속적 관리가 필요하다. 그리고 교목층의 비나 자무 임분의 밀도 조절에 따른 관목층에서 수광조건이 개 선된 곳에 비자나무의 천연갱신을 유도하여야 할 것이며, 이를 우점종으로 하는 안정된 생태기반 위의 식물군집구 조를 유도할 때 건전한 비자나무림의 조성이 가능할 것으 로 판단된다. 따라서 조사된 우리나라 비자나무림에서 선 택적 간벌시업을 통한 과도한 수관경쟁의 조정이 가능 할 것이며, 조사된 식물군집특성의 이해에 따른 임분유형 특 성별 현재 이러한 시업이 적절히 시행될 때 비자나무림의 안정된 생태계가 유지될 수 있을 것이다.
인용문헌
1. Agnew, A.D.Q. (1961) The ecology of Juncus effusus L.
in North Wales. Journal of Ecology 49(1): 83-102.
2. Hill, M.O. (1979) TWINSPAN-A FORTRAN program for arranging multivariate data in an order of two-way table by classification of the individuals and attributes. Ithaca, N. Y. Cornell University. Press, pp. 50.
3. Lee, C.H., Choi, Y.C., Kim, S.H. and Kwon, K.W. (2000)
Site characteristics, and vegetation structure, and dynam- ics of forest communities growing Kalopanax septemlo- bus (Thunb.ex Murray) Koidz. in Gangwon-do. Korean Journal of Plant Resources 13(3): 227-242.
4. Lee, S.D., Lee, K.J. and Choi, J.W. (2009) Management plan to consider ecological characteristic of Pinus densi- flora community in Seoul. Korean Journal of Environ- ment and Ecology 23(3): 258-271.
5. Park, M.S. (2007) Vegetation structure and growth envi- ronment of Torreya nucifera Forest in Mt. Gaecheon.
Korean Institute of Forest Recreation 11(1): 15-25.
6. Park, N.C., Jung, C.G., Choe, J.C., Shin, H.C., Kwon, Y.H., Cho. B.H., Kang, J.T., Chung, Y.G. and Kim, C.S.
(2001) Relationships on the growth pattern and site envi- ronmental factors in Castanopsis cuspidata var. sieboldii . KFRI Journal of Forest Science 64: 1-13.
7. Shin, H.C., Park, N.C., Song, H.K., Jung, C.G., Choe, J.C., Kwon, Y.H. and Jo, B.H. (2001) The analysis of vegeta- tion-environment relation of the Dendropanax morbifera forests by TWINSPAN and DCCA. KFRI Journal of For- est Science 64: 14-26.
8. Shin, H.C., Park, N.C., Song, H.K., Jeong, Y., Choi, J., Ykwon, Y., Lee, K.S. and Kim, Y. (2002) Analysis of vegeta- tion structure and vegetation-environment relationships in the Machilus thunbergii stands, Korea. Journal of Korean Forestry Society 91(6): 765-774.
9. Shin, H.C., Park, N.C. and Song, H.K. (1999) The vege- tation structure and community classification of Quercus acuta in warm-temperate region of Korean peninsula.
KFRI Journal of Forest Science 60: 11-25.
10. Shin, H.C., Chae, C.J., Jeong, Y., Park, N.C., Lee, S.T. and Song, H.K. (2004) Structure and correlation with environ- ment factors in Castanopsis cuspidata var. sieboldii For- ests. KFRI Journal of Forest Science 67: 8-19.
11. Shin, H.C., Lee, K.Y. and Song, H.K. (1998) The analy- sis of vegetation-environment relationship of the Taxus cuspidata forests by TWINSPAN and DCCA. Journal of Korean Forestry Society 87(4): 535-542.
12. Song, H.K. (1990) An Analysis of Vegetation - Environ- ment Relationships of Mt. Gyeryong and Mt. Deokyu by Detrended Canonical Correspondence Analysis. Journal of Korean Forestry Society 79(2): 216-221.
13. Ter Braak, C.J.F. 1987. CANOCO-a FORTRAN program for canonical community ordination by [partical] [dynni- cal] [cannical] correspondence analysis, principal compo- nents analysis and redundancy analysis(version2.1) TNO Institute of applied computer science, Statistics Depart- ment, Wageningen, The Netherlands.
(2010년 2월 23일 접수; 2010년 3월 22일 채택)