남극 로스해에서 분리한 Croceibacter atlanticus 균 유래 리파아제의 생산, 고정화, 효소특성 연구
박채경, 김형권*
가톨릭대학교생명공학과
Received: April 12, 2018 / Revised: June 23, 2018 / Accepted: June 9, 2018
서 론
남극해는태평양, 대서양, 인도양의최남부를차지하며표 층의수온이빙점에가까운남극대륙을둘러싸고있는해 역이다. 지구상에서가장추운미지의장소이며그만큼새로 개발할수있는생물자원이풍부한곳이다. 극지의생물자원 은극지의환경에서생존할수있는생물학적특성을가지 고있기때문에산업적활용가능성이매우높을것으로추 정한다.
리파아제효소(EC 3.1.1.3)는지방가수분해효소이며트
리글리세리드의에스테르결합을가수분해하여글리세롤과 지방산을생성하는효소이다. 반응조건에따라서에스테르 합성반응, 트랜스에스테르화반응도촉매하는산업용효소 이다. 한편, 극지미생물이생산하는리파아제효소들은다 양한온도및 pH 특성, 기질특이성, 입체특이성을갖고있 는것으로밝혀져서식품, 제지, 바이오디젤, 세제, 화장품산 업의효소촉매로연구되고있다[1].
지구상특수환경에서발굴한리파아제를산업용촉매로활 용하기위해서는유전공학기법으로재조합효소를생산해야 한다. 발굴한리파아제유전자 ORF를특수프로모터를지 닌발현벡터에넣어미생물을형질전환하고, 배지조성, 배양 온도, 배양시간, 유도물질농도, 유도배양시간등을조절하 여활성형태로대량생산하면적은비용으로재조합리파아 Production, Immobilization, and Characterization of Croceibacter atlanticus Lipase Isolated from the Antarctic Ross Sea Chae Gyeong Park and Hyung Kwoun Kim*
Division of Biotechnology, The Catholic University of Korea, Bucheon 14662, Republic of Korea
The Antarctic Ocean contains numerous microorganisms that produce novel biocatalysts that can have applications in various industries. We screened various psychrophilic bacterial strains isolated from the Ross Sea and found that a Croceibacter atlanticus strain (Stock No. 40-F12) showed high lipolytic activity on a tributyrin plate. We isolated the corresponding lipase gene (lipCA) by shotgun cloning and expressed the LipCA enzyme in Escherichia coli cells. Homology modeling of LipCA was carried out using the Spain Arreo lake metagenome alpha/beta hydrolase as a template. According to the model, LipCA has an α/β hydrolase fold, Gly-X-Ser-X-Glymotif, and lid sequence, indicating that LipCA is a typical lipase enzyme.
Active LipCA enzyme was purified fromthe cell-free extract by ammonium sulfate precipitation and gel fil- tration chromatography. We determined its enzymatic properties including optimum temperature and pH, stability, substrate specificity, and organic solvent stability. LipCA was immobilized by the cross-linked enzyme aggregate (CLEA) method and its enzymatic properties were compared to those of free LipCA.
After cross-linking, temperature, pH, and organic solvent stability increased considerably, whereas sub- strate specificities did not changed. The LipCA CLEA was recovered by centrifugation and showed approx- imately 40% activity after 4th recovery. This is the first report of the expression, characterization, and immobilization of a C. atlanticus lipase, and this lipase could have potential industrial application.
Keywords: Lipase, Croceibacter atlanticus, the Antarctic Ross Sea
*Corresponding author
Tel: +82-2-2164-4890, Fax: +82-2-2164-4865 E-mail: [email protected]
© 2018, The Korean Society for Microbiology and Biotechnology
제효소를만들수있다. 또한, 안정성을향상시키고반복사 용할수있도록효소를고정화해야한다. 효소의고정화방 법으로 흡착(adsorption), 포괄법(Entrapment), 가교결합 (Cross-linking) 방법이일반적으로사용되고있다[2]. Cross- linked enzyme aggregate (CLEA) 방법은가교결합법의일
종으로가교제를사용하여효소내의 Lysine 잔기를공유결
합시킴으로써안정성을증대시키는방법이다. 또한고가의
비드(bead)를사용하지않고쉽고저렴하게고정화하는방
법이기때문에많이연구되고있다[3].
현재까지많은미생물리파아제효소에대한 X선결정구 조가밝혀져서보고되었다. 대부분의경우, 효소구조의중심 에 parallel β-sheet를지닌α/β hydrolase fold를갖고있다 [4]. 활성부위에 Ser-His-Asp의 catalytic triad를가지며, 이 중에서 Ser은 Gly-X-Ser-X-Gly 보존서열속에들어있다[5].
또한, 활성부위포켓이 amphipathic α-helix로구성된 lid로
덮여 있는데 이 구조가 리파아제 특이적인 ‘Interfacial
activation’ 기작과밀접하게관련되어있다. 에스터라제효 소의경우, 리파아제와대부분유사한구조를갖고있지만, 활성부위에 lid 구조를갖고있지않다.
본 실험에서는 남극 로스해(Ross sea)에서 분리한 Croceibacter atlanticus Stock No. 40-F12 균으로부터 shotgun cloning 방법을통해리파아제유전자(lipCA)를찾 고 Escherichia coli에서 대량 생산하였다. 현재까지 C.
atlanticus 균으로부터리파아제효소의분리및특성규명
에대한보고가전혀없기때문에 LipCA의특성을규명하
고고정화함으로써산업적으로이용할수있는가능성을알 아보았다.
재료 및 방법
실험재료
p-Nitrophenyl acetate (C2), p-nitrophenyl butyrate (C4), p-nitrophenyl caprylate (C8), p-nitrophenyl caprate (C10), p-nitrophenyl laurate (C12), tributyrin (TBN), tricaprylin (TCN), 올리브유 (olive oil), glutaraldehyde (25% H2O), amipicillin, isopropanol, 1,2-dimethoxyethane, acetonitrile, 1-butanol, 2-butanone, propionitrile은 Sigma Aldrich Co. (USA)에서구입하였으며, p-nitrophenyl caproate (C6)는 Tokyo Chemical Industry Co. (Japan)에서 구입하 였다. Dimethylsulfoxide (DMSO), acetone, pyridine, ethyl acetate는 Junsei Co. (Japan)에서구입하였으며, methanol, ethanol은 Merck Chemical Co. (Germany)에서구입하였다. β- D-thiogalactopyranoside (IPTG)는 Duchefa Biochemie B.V. Co. (Netherlands)에서구입하였으며, Marine 배지는 Becton, Dickinson and Co. (USA)에서구입하였다.
지방분해 효소 생성 균주 분리
남극로스해(Ross sea)에서분리한균중에서지방분해활
성을가지고있는균을선별하기위해 TBN/Marine agar 배 지를이용한지방분해균주스크리닝을진행하였다. TBN/
Marine agar 배지의제조법은 다음과 같다. TBN과 gum arabic 용액(200 mM NaCl, 10 mM CaCl2, 10% gum arabic) 을 1:9 부피비율로섞어멸균한다. 이것을 Waring blender 에넣고고속으로 2분간유화시켜 10% TBN emulsion을만 든후, 이것과 Marine agar 배지를다시 1:9 부피비율로혼 합해서 TBN/Marine agar 배지를만든다. 분리균을 15℃에 서 96시간동안배양한후, 가장뚜렷한투명환을생성하는 균주를선발하였다.
지방분해 효소 유전자 클로닝과 서열분석
지방분해활성이가장큰 Croceibacter atlanticus 균으로 부터 리파아제 유전자를 분리하기 위해서 다음과 같이 shotgun cloning을 수행하였다. C. atlanticus 균으로부터 genomic DNA을 추출한 후, HindIII를 처리하여 DNA library를만들었다. DNA library를 pUC19 vector에연결시 켜 E. coli XL1-Blue에 형질전환 시킨 후, ampicillin (100μg/ml)을포함한 TBN/Luria Bertani (LB) agar 배지 에도말하고 37℃에서배양하였다. 투명환을생성하는 colony 를 선별하여 plasmid (pUC19-lip1)를 추출한 후, sub cloning을 진행하였다. pUC19-lip1 plasmid를 HindIII와 EcoRI으로처리하여 1.3 kb 크기의 DNA 조각을얻었다. 이 것을 pUC19 vector와연결하여 E. coli XL1-Blue에형질전 환함으로써재조합 plasmid (pUC19-lip2)를제조하고삽입 된 DNA의염기서열을분석하였다. 리파아제유전자(lipCA) 의염기및아미노산서열분석과데이터베이스검색은각 각 DNAStar 프로그램과 National Center for Biotechnology Information (NCBI)의 BLAST 프로그램을이용하였다.
16s rRNA 동정
C. atlanticus (Stock No. 40-F12)의 genomic DNA을 514F, 1542R primer를이용하여 16s rRNA 분석을진행하 였다. 514F; 5'-CGT GCC AGC AGC CGC GGT-3', 1542R; 5'-AGA AAG GAG GTG ATC CAC CC-3'. 16s rRNA의 부분 염기 서열 분석과 데이터베이스 검색은 DNAStar 프로그램과 National Center for Biotechnology Information (NCBI)의 BLAST 프로그램을이용하였다.
상동성 모델링
LipCA의 아미노산 서열을 SWISS-MODEL (https://
swissmodel.expasy.org)에입력하여상동성모델링을진행하 였다. LipCA의상동성모델은 Spain Arreo lake metagenome
에서유래한 alpha/beta hydrolase enzyme (PDB ID 5JD4) [6]을기준으로수행하였고 PyMOL 프로그램을통해단백질 3차구조를만들었다.
리파아제 유전자(lipCA) 발현
재조합 plasmid (pUC19-lip2)에삽입된 lipCA 유전자를 lipCA-F/lipCA-R 프라이머를사용한 PCR반응을통해증폭 하였다. lipCA-F; 5'-GAT CAT ATG AAT CCT GCT GTT TTT-3', lipCA-R; 5'-GAT AAG CTT TTA CAA CCG CTG AGC-3'. 증폭한 DNA를 pGEM-T vector에연결시켜재조합 플라스미드(pGEM-T-lipCA)로만들고 E. coli XL1-Blue에 형질전환시켰다. E. coli에서분리한재조합 plasmid를 NdeI 과 HindIII로처리하여 lipCA 유전자를얻었다. 이것을 pET- 22 vector에연결하여재조합 plasmid (pET-22-lipCA)를만 들고 E. coli BL21 (DE3)에형질전환시켰다. 형질전환된 E.
coli 균을 ampicillin (100 μg/ml)이포함된 LB에접종시켜 37℃에서흡광도(600 nm)가 0.5가될때까지배양시켰다. IPTG (0.1 mM)을첨가하여추가로 20℃에서 20시간배양 시켰다. 배양후에원심분리(3,500 ×g, 10 min)로균을모으 고증류수로세척시킨후에다시증류수로재부유시켰다. 초 음파 분쇄법으로 균을 파쇄시킨 후 원심분리(14,000 ×g, 10 min)하여세포추출물(cell-free extract)를얻고, 리파아
제정제와 CLEA 제조를수행하였다.
LipCA의 정제
세포추출액에 ammonium sulfate를 30% 포화되도록넣 고 1시간동안 단백질을침전시켰다. 원심분리(14,000 ×g,
4 min)하여상층액을버리고침전된단백질에완충용액 PBS
(Phosphate Buffer Solution, NaCl 137 mM, KCl 2.7 mM, Na2HPO4 10 mM, KH2PO4 1.8 mM)를넣고재부유시켰다. SuperoseTM 12 10/300 GL gel filtration column (GE healthcare, Buckinghamshire, UK)을 장착한 FPLC에서 겔여과크로마토그래피를통해단백질의탈염과정제를진 행하였으며이때용출용액으로 20 mM Tris-HCl (pH 8.5)를 사용하였다.
LipCACLEA의 제조
세포추출액에 ammonium sulfate를 30% 포화되도록넣 고 1시간동안단백질을침전시켰다. 침전시킨후에가교제 인 glutaraldehyde (25% 수용액)을각각 25, 50, 75, 100 mM 농도로첨가하고 10℃에서 24시간동안교반시켰다[7, 8]. 원 심분리(14,000 ×g, 10 min)를 통해 제조된 LipCACLEA를 50 mM potassium phosphate (pH 8.0)로 2회 세척하고 500μl의동일한완충용액으로현탁하여 10℃에서보관하 였다.
리파아제 활성 측정
리파아제활성은다음과같은기준조건에서측정하였다. 0.1 mM p-nitrophenyl caprylate (pNPC)를기질로사용하 였으며 50 mM Tris-HCl (pH 8.5) 완충용액에기질과효소 액을넣고 37℃에서 3분동안반응시켜흡광도(405 nm)를 측정하였다. 효소활성 1 unit은 1분 동안 1 μmol의 p- nitrophenol을생성하는효소의양으로정의하였다[9, 10].
온도, pH 및 NaCl 농도에 대한 특성
LipCA와 LipCACLEA의최적온도를확인하기위해 10℃에 서 80℃사이의온도에서효소반응을진행하였다. 효소활성
측정은 pNPC assay로 진행하였으며, 완충용액으로는
50 mM potassium phosphate (pH 8.0)을사용하였다. 최적 pH도 pNPC assay를통해진행하였으며반응온도는 37℃에서진행하였다. pH 7.0에서 pH 10.0까지측정하였으 며 사용된 완충용액은 다음과 같다. 50 mM potassium phosphate (pH 7.0−8.0), 50 mM Tris-HCl (pH 8.5−9.0), 50 mM sodium bicarbonate (pH 9.5−10.0).
기질에 대한 특이성
기질특이성을 조사하기위하여 pNP ester assay와 pH
stat assay로다양한기질에대한분해능력을측정하였다.
pNP ester assay는기준조건에서탄소수가다른다음과같
은 기질을 사용하였다. p-nitrophenyl acetate, butyrate, caproate, caprylate, caprate, laurate.
pH stat assay는 triglycerides에대한기질특이성을확인 하기위한방법으로사용되었으며기질로 1%의 TBN, TCN, 그리고올리브유를사용하였다. pH stat assay의반응조건 은 37℃에서 3분이며, 효소반응이진행되기전 1%의기질과 10% gum arabic이유화된 emulsion에 10 mM NaOH 용액 을첨가하여 pH를 8.0으로맞춘다. 효소반응이진행되면서 triglycerides는 fatty acid와 glycerol로분해되어 NaOH 용
액이첨가된다. 첨가된 NaOH 양으로효소활성을측정하며
1 unit은 1분동안 1 μmol의 fatty acid를생성하는효소의 양으로 정의하였다. pH stat assay는 718 Titrino pH titrator (Metrohm, Switzerland)로측정하였다[11].
온도, pH 및 유기용매에 대한 안정성 실험
LipCA와 LipCACLEA의온도에대한안정성을확인하기위 해 10℃에서 80℃까지 10℃간격으로 1시간동안방치한후 pNPC assay를 수행하였다. LipCA와 LipCACLEA 모두 50 mM potassium phosphate (pH 8.0)을완충용액으로사 용하였다. 효소의잔존활성은기준조건의 pNPC assay를 통하여측정하였으며, 기준조건은 LipCA와 LipCACLEA 를각 각 50 mM Tris-HCl (pH 8.5, pH 9.0)의완충용액에서효소
와 0.1 mM pNPC을섞고 37℃에서 3분동안반응시킨후, 생성되는 p-nitrophenol의양을흡광도(405 nm)로측정하는 것이다.
리파아제의 pH 안정성을확인하기위해효소를 pH 1.5− 12.0 조건에서 1시간동안방치한후, 잔존활성을측정하였 다. 사용된 pH에따른완충용액은다음과같다. 50 mM KCl- HCl (pH 1.0−2.0), 50 mM glycine-HCl (pH 3.0), 50 mM acetate-sodium acetate (pH 4.0−5.0), 50 mM potassium phosphate (pH 6.0−8.0), 50 mM Tris-HCl (pH 9.0), 50 mM sodium-bicarbonate (pH 10.0), 50 mM NaHCO3- NaOH (pH 11.0), 50 mM KCl-NaOH (pH 12.0). 효소의잔 존활성을측정하기위해 pNPC assay를사용하였으며 37℃ 에서 3분동안 LipCA는 50 mM Tris-HCl (pH 8.5)의완충 용액, LipCACLEA는 50 mM Tris-HCl (pH 9.0)의완충용액 에서반응시킨후잔존활성을측정하였다.
유기용매에대한안정성을실험하기위해유기용매에 1시 간동안효소를방치한후, 기준조건하의 pNPC assay를통 해잔존활성을측정하였다. 사용된유기용매는다음과같 다. DMSO, methanol, acetonitrile, ethanol, acetone, 1,2- dimethoxyethane, isopropanol, propionitrile, 2-butanone, pyridine, ethyl acetate, 1-butanol.
LipCACLEA의 회수
LipCACLEA의회수율을측정하기위한회수방법으로원
심분리(19,000 ×g, 10 min)를이용하였다. 총다섯번의원 심분리를 진행하였으며 원심분리 후의 상층액과 50 mM potassium phosphate (pH 8.0)로재부유한하층액의잔존활 성을기준조건에서 pNPC assay를통해측정하였다.
결과 및 고찰
지방분해 효소 생성 균주 분리와 유전자 클로닝, 서열분석 과 발현
남극로스해(Ross sea)로부터추출한 42개의분리균 중 24개의균이 TBN/Marine agar 배지에서투명환을형성했 으며, 이중에서가장큰활성을보이는분리균인 Croceibacter atlanticus (Stock No. 40-F12, 75°44'S, 176°57'E, depth
320 m)에서리파아제효소유전자를찾는실험을진행하였
다(Supplementary Fig. S1). HindIII 제한효소를사용하여 shotgun cloning을 진행한 결과, 약 10 kb 크기의 insert DNA를가지고있는재조합플라스미드(pUC19-lip1)를얻 었다. 이후, EcoRI으로 10 kb DNA를자른결과약 1.3 kb와
9 kb 크기의유전자조각을얻었고이것들을각각 pUC19에
연결하여 두 개의 재조합 plasmid를 만들었다. 재조합
plasmid를 E. coli XL1-Blue에 형질전환 시켜 TBN/LB agar배지에서 screening한결과 1.3 kb 크기의 DNA를가지 고있는 E. coli에서투명환이형성되었다. Sub cloning으로
찾아낸리파아제유전자는 pUC19 vector에연결되어있으
Fig. 1. Nucleotide and amino acid sequence of Croceibacter atlanticus LipCA. Catalytic residues are underlined.
므로 M13-20F/M13-47R primer를사용하여서열분석을수 행하였다. DNAStar 프로그램을통해 1.3 kb 전체염기서열 중에서하나의 ORF를찾아내었는데, 이것은 1,029 bp 크기 의염기서열로구성되어있고 342개의아미노산서열을코 딩하였다. 아미노산서열로부터계산된단백질의분자량은 37,099 Da이며, 리파아제/에스터라제의 signature sequence 인 Gly-Asp-Ser-Ala-Gly motif를가지고있는것으로밝혀졌 다(Fig. 1).
C. atlanticus 균에서분리한리파아제유전자를 lipCA라 고명명하였다. lipCA를 NCBI의 BLAST 프로그램으로검 색한결과, Alcanivorax sp. DG881의 putative alpha/beta hydrolase (Sequence ID, WP_007151635)와 100% 상동성 을보였다(Fig. 2). Alcanivorax sp. DG881균은남극해에서 분리한균인데 C. atlanticus 균과는전혀다른 phylum에속 하는균이다[12, 13]. 분리균주(Stock No. 40-F12)에대해서 16s rRNA 분석을진행한 결과, C. atlanticus HTCC2559 와 99.9%의상동성을보였으며, Alcanivorax sp. DG881와 는 79.2%의낮은상동성을보였다(Supplementary Fig. S2).
그럼에도불구하고 C. atlanticus 균과 Alcanivorax 균이동 일한리파아제효소유전자를가지고있다는사실은매우흥
미로운일이며남극의동일환경에서사는균들간에 gene
transfer가진행됐을가능성을보여준다. 아직까지 C. atlanticus 균으로부터 리파아제가 보고되지 않았고, Alcanivorax
putative hydrolase에대한연구가전혀진행되지않았기때
문에본연구팀은 LipCA 효소를대량생산하고특성규명연
구를진행하였다.
LipCA 상동성 모델링
TBN agar 배지에서 screening을 진행하였기 때문에
lipCA가 lipase 유전자인지 esterase 유전자인지확인할필 요가있다. Lipase와 esterase의구조적차이점은 amphipathic α-helix lid 유무이다. LipCA의 3차원구조를알아보기위해 SWISS-MODEL 프로그램의 protein structure homology- modelling server를이용해서상동성모델링을진행하였다. LipCA와 38.24% 동일성을 갖고 있는 Spain Arreo lake hydrolase를주형으로모델링을수행한결과, LipCA는중심 에 8개의 strand로 구성된 parallel β-sheet를 지닌 α/β hydrolase 구조를갖는것으로밝혀졌다. 활성부위에 Ser191- His315-Asp285의 catalytic triad를가지고있으며, 활성부위 포켓이 amphipathic α-helix lid (Leu242-Phe252)에덮여있는
전형적인 리파아제 효소임을 알 수 있다(Supplementary
Fig. S3)[5, 7].
LipCA 효소의 분리
lipCA 유전자를 pET-22에 클로닝하여 E. coli BL21 (DE3)에형질전환하였다. E. coli 배양액에 IPTG를첨가하 Fig. 2. Neighbor-joining tree showing the phylogenetic positions of Croceibacter atlanticus LipCA and some other related pro- teins. For making phylogenetic tree, DNAStar megalign program was used.
여 20℃에서 20시간배양한결과, LipCA가대량으로생산 되었으며, SDS-PAGE 분석을통해서 LipCA가수용성단백
질형태로세포추출액의상층액에존재함을알게되었다(Fig.
3). 세포추출액에 ammonium sulfate를 30% 포화되도록첨 가하여 salting-out을수행한결과, 대부분의 LipCA 단백질 이침전되었다. 소량의용액으로단백질을현탁한후, 겔여 과크로마토그래피를통해탈염과효소분리를동시에진행
하였다[14]. 일반적으로 대부분의 단백질이 ammonium
sulfate 30−70% 포화용액에서침전되는데 반해서[15, 16], LipCA가 ammonium sulfate 0−30% 포화용액에서침전된
것은매우특이한결과인데이것은 LipCA 효소표면에소
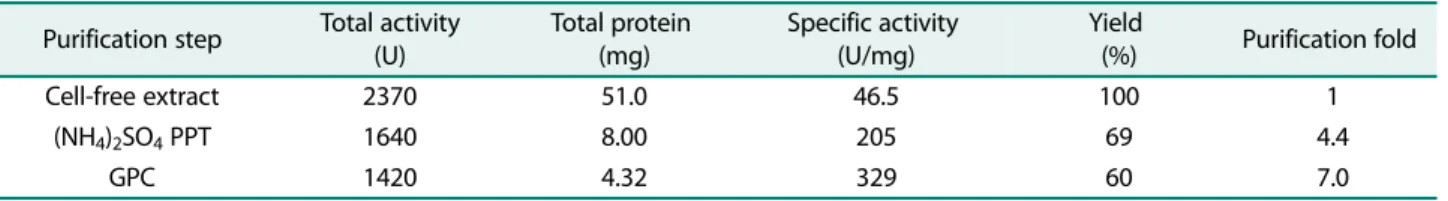
수성아미노산잔기가많이분포하기때문이라고추정한다. 분리된 LipCA는 SDS-PAGE 상에서약 37 kDa 크기를나타 냈으며(Fig. 3), 329 U/mg의활성을가지고있다(Table 1).
SDS-PAGE 결과를통해 LipCA가분리되었음을확인하였
으며분리된 LipCA를이용해서효소특성연구를수행하였다.
LipCACLEA의 제조
세포추출액에 ammonium sulfate를 30% 포화되도록넣 어 LipCA를선택적으로침전시킨후, 4가지농도(25, 50, 75,
100 mM)의 glutaraldehyde를첨가하여 LipCACLEA를제조 하였다. 생성한 4종류의 LipCACLEA의 activity retention은 세포 추출액의효소활성을 100%로기준 잡았을때, 각각 108%, 73%, 37%, 24%로측정되었다(Supplementary Table S1). Glutaraldehyde의농도가높아질수록활성이줄어드는
이유를다음과같이설명할수있다. LipCA 단백질은전체
아미노산조성의 10%를 lysine과 arginine으로구성하고있 다. Glutaraldehyde는가교제로서 lysine 및 arginine 잔기
들사이를 schiff base 형태의공유결합으로연결하기때문
에침전된 LipCA 단백질사이를결합시킬수있다[17, 18].
하지만, Glutaraldehyde의농도가높아질수록단백질분자 내부및분자간공유결합이점점많아지게되면서 LipCA 단 백질분자에서 lysine 및 arginine가원래형성하고있던이 온결합이깨지게된다. 즉, 효소의활성부위유연성이떨어 지는현상과함께구조변성이동시에진행되는것으로추
정된다[7, 8, 19]. 따라서 본연구에서는 활성이가장 높은
25 mM 농도의 glutaraldehyde로만든 LipCACLEA를사용하 여특성연구를수행하였다.
LipCA, LipCACLEA의 온도, pH에 대한 특성과 안정성 실험 LipCA와 LipCACLEA의온도와 pH에대한특성을 pNPC 기질에대한분해활성을측정함으로써확인하였다. LipCA 와 LipCACLEA의 최적 온도는 모두 40℃이며(Fig. 4A) LipCA의최적 pH는 pH 8.5, LipCACLEA의최적 pH는 pH 9.0인것으로밝혀졌다(Fig. 4B). 효소의고정화로인하여최 적 pH가증가한것으로미루어보아효소의활성부위가일 부변형되었음을알수있다.
LipCA와 LipCACLEA의온도, pH에대한안정성실험을수 행함으로써고정화로인한효소의안정성증가를확인할수 있었다. 먼저열안정성의경우, LipCACLEA가 LipCA보다 20℃이상높은온도에서더안정하였다(Fig. 4C). 또한 pH 안정성에대해서도 LipCACLEA가 LipCA보다 pH 안정성범 위가 넓어졌다(Fig. 4D). Glutaraldehyde에 의해 진행된 LipCA 단백질의 lysine 및 arginine 잔기간의공유결합을통 해효소가견고한구조를이루게되었음을알수있다[14, 15].
기질 특이성
기질에대한특이성을알아보기위하여 pNP-ester assay
Fig. 3. Expression and purification of LipCA. Lane 1, protein size marker; lane 2, cell-free extract of E. coli cells harboring empty vector; lane 3, insoluble fraction of E. coli cells expressing LipCA; lane 4, cell-free extract of E. coli cells expressing LipCA;
lane 5, purified LipCA by 30% ammonium sulfate precipitation;
lane 6, purified LipCA by Gel filtration chromatography after ammonium sulfate precipitation.
Table 1. Purification of LipCA.
Purification step Total activity (U)
Total protein (mg)
Specific activity (U/mg)
Yield
(%) Purification fold
Cell-free extract 2370 51.0 46.5 100 1
(NH4)2SO4 PPT 1640 8.00 205 69 4.4
GPC 1420 4.32 329 60 7.0
와 pH stat assay를수행하였다. 다양한탄소길이를지닌 pNP 기질을대상으로 pNP-ester assay로측정한결과, 탄 소수가 8개인 p-nitrophenyl caprylate를기질로사용하였을 때상대활성이가장높았다(Fig. 5A). pH stat assay의경우, 탄소수가 4개인 butyric acid로구성된 TBN을기질로사용 하였을때가장높은활성을보였다. 그러나탄소수가 18개
인 oleic acid를지닌올리브유를기질로사용하였을때에도
높은상대활성을보였다(Fig. 5B). 이것은 LipCA가짧은탄 소사슬뿐만아니라긴탄소사슬도자를수있는리파아제효 소임을의미하며, glutaraldehyde를이용한고정화과정이 효소의활성부위에영향을미치지않음을의미한다.
유기용매에 대한 안정성 실험
LipCA를대상으로 10% 유기용매에서의안정성실험을진
행하였다. 실험에서사용한 12종류의유기용매중, 친수성 유기용매에서높은안정성이드러났으며(Fig. 6A), 그결과
를바탕으로 30% 친수성유기용매에서안정성실험을진행
하였다. LipCACLEA는 30% 친수성유기용매에서만안정성 실험을 진행하였다. 그 결과 LipCA와 LipCACLEA 모두 DMSO와 1,2-dimethoxyethane에서상대적활성이높았다 (Fig. 6B) [20, 21]. 30% 1,2-dimethoxyethane에서 상대적 활성이높은이유는 1,2-dimethoxyethane이단백질침전제 로작용하였기때문이다[22]. 30% ammonium sulfate로침 전시켜 1차로분리한 LipCA의전체효소활성이겔여과크 로마토그래피를이용한 2차분리한 LipCA의전체효소활성
보다크다. 이는 LipCA는응집되어있을때가수용성상태
였을때보다활성이더높다는것을시사한다. 30% 친수성
유기용매에대해서 LipCACLEA도 LipCA와비슷한그래프양 상을보였지만상대적활성이 DMSO와 1,2-dimethoxyethane 에서더높다. 이는고정화로인하여유기용매에대한안정 성이증가하였다는것을의미한다(Fig. 6B).
LipCACLEA 회수
본연구에서는 LipCA 효소를 CLEA 방법으로고정화하 Fig. 4. Effect of temperature and pH on LipCA and LipCACLEA activity and stability. (A) Effect of temperature on LipCA and LipCACLEA activity, (B) Effect of pH on LipCA and LipCACLEA activity, (C) Effect of temperature on LipCA and LipCACLEA stability, (D) Effect of pH on LipCA and LipCACLEA stability. Open circle, LipCA; closed circle, LipCACLEA.
였다. 이방법의가장큰장점은저렴한제조비용과손쉬운 회수에있다. 즉, 원심분리를이용해서 CLEA 효소를빠르 고효과적으로회수할수있다. 본실험에사용된 LipCACLEA 는 glutaraldehyde를첨가하고 48시간이지난 LipCACLEA를 사용하였다. 총다섯번의원심분리를수행하였으며, 4번의 회수후에 40% 이상의활성을유지하였다(Fig. 7). LipCACLEA 를 여러 번 사용하면 활성이 점차적으로 감소하는 것은
CLEA로부터효소가떨어지면서생기는결과이고, LipCA 효소의 Lys 함량이적고 cross-linker로사용한 glutaraldehyde 의분자길이가짧기때문이라고추정된다. 기존문헌[23]에 따르면, BSA 첨가, pH 조절, dextran aldehyde 사용등을 통해서효소가떨어지는것을방지할수있다고밝혀졌기때
문에 LipCA의경우에도향후에고정화의최적화연구를통
해효소활성을안정화시킬수있을것으로기대한다. Fig. 5. Substrate specificity of LipCA and LipCACLEA. (A) Various p-NP esters were used as substrates. (B) Three triglycerides were used as substrates and pH stat assay was used. Open bar, LipCA; closed bar, LipCACLEA.
Fig. 6. Effect of organic solvents on the stability of LipCA and LipCACLEA. (A) Enzyme was pre-incubated with 10% organic solvent for 1 h at room temperature, and residual activity was measured, (B) Enzyme was pre-incubated with 30% organic solvents for 1 h at room temperature, and residual activity was measured. Log P values are as follows. DMSO (-1.3), Methanol (-0.76), Acetonitrile (-0.34), Ethanol (-0.24), Acetone (-0.23), 1,2-Dimethoxyethane (-0.2), Isopropanol (-0.05), Propionitrile (0.17), 2-Butanone (0.29), Pyridine (0.65), Ethyl Acetate (0.73), 1-Butanol (0.839). Open bar, LipCA; closed bar, LipCACLEA.