독도 주변의 해수에서 분리한 세균의 다양성과 군집구조 분석
성혜리·김사열*
경북대학교 생명과학부, 경북대학교 울릉도·독도연구소
Bacterial Diversity and Distribution of Cultivable Bacteria Isolated from Dokdo Island. Sung, Hye Ri and Sa-Youl Ghim*. School of Life Sciences and Research Institute for Ulleungdo & Dokdo Islands, Kyungpook National University, Daegu 702-701, Korea − One hundred sixty three strains showing different colony morphological characteristics on different concentration of marine agar (MA) plates were isolated from ambient seawater near Dokdo island. Bacterial diversity and distributions were studied by phylogenetic analysis of the partial 16S rRNA gene sequences. One hundred sixty three strains were partially sequenced and analyzed phylogenetically. They were composed of 5 phyla, of which gamma-proteobacteria (58%), alpha-proteobacteria (20%), bacteriodetes (16%) were predominant. They were affiliated with 90 species.
The 16S rRNA sequence similarity of the isolates was in 93.3 to 100 % range to reported sequence data.
Thirty six isolates of among them were assumed to be novel species candidates based on similarity analysis of the 16S rRNA gene sequences. Overall, Proteobacteria and Bacteriodetes of the Dokdo coastal sea water showed a high diversity.
Key words: Dokdo, sea water, bacterial diversity, proteobacteria, bacteriodetes
서 론
독도는 북서태평양 연해인 동해의 중남부에 위치하고 있 는 화산섬으로 수리적 위치는 동도, 서도 및 그 주변에 흩어 져 있는 89개의 바위섬으로 이루어진 화산섬이다. 동도는 동 경 131도 52분 10.4초, 북위 37도 14분 26.8초에, 서도는 동경 131도 51분 54.6초, 북위 37도 14분 30.6초에 위치하 는 아한대남단 해역에 속해 있지만, 대만난류와 북한한류가 교차하는 해역으로 이 난류와 한류의 세력에 따라 다양한 시 공간적이 변화를 가져온다[16]. 이곳은 플랑크톤이 풍부하여 회유성 어족이 풍부하고, 독도 주변의 파식대지와 암초에는 미역, 다시마 등의 해조류와 전복, 소라 등 패류가 많이 서 식하고 있다. 독도의 표면 수온은 바다 표층수온이 9.0~26.0oC의 범위 내에서 변하는데, 3~4월에 10oC 전후로 가장 낮고 8월에 25oC 전후로 가장 높은 것으로 알려져 있 다. 표면수 염분 농도는 33~34%, 표층 산소량은 6.0 ml/L, 투명도는 17~20 m로 비교적 높은 염분 농도와 맑은 수역을 형성하고 있다.
해양생태계는 식물, 동물플랑크톤 및 미생물 등 구성원들 간 서로 밀접한 관계를 가지고 상호작용 한다. 그러므로 어 느 한 구성원이 변화를 보이면 다른 구성원의 변화를 초래 하게 된다[26]. 특히 해양미생물군집은 유기물 분해와 영양
물질 순환 등으로 인하여 해양 생태계에서 생산자이면서 분 해자로 생태계의 안정한 동적 평형을 유지시키는 중요한 생 물군집이다. 그래서 해양생태계의 가장 기본적인 단위인 미 생물에 의한 역할과 그 중요성이 점차 커지고 있다[6, 9, 22].
해양생태계는 매우 다양한 생물들에 의해 복잡, 다양하게 구 성되어있기 때문에, 해양세균의 분포 종류와 군집은 물리 화 학적 환경에 의해 지배된다[25, 37].
1990년대 후반 독도 연구를 시작한 이후 독도의 식물군집 과 동물군집에 대해서는 여러 연구팀에 의해 꾸준히 보고 되 어왔고, 미생물에 대해서는 2004년부터 미생물 탐사가 시작 된 이래로 2005년 윤 등[41]에 의해 독도 해수에서 Virgi- bacillus dokdonensis를 분리하여 보고하였고, 이어서 2009 년까지 새로운 속 4종을 포함한 40여 개의 새로운 종의 미 생물을 분리하여 보고하였다. 그러나 독도에 존재하는 미생 물군집에 관한 연구는 이루어지지 않았고, 2009년 전 등[20]
에 의해 독도에 서식하는 식물근권에서 분리한 세균군집의 분포에 대해 처음 보고 되었다. 독도는 육지와 격리된 지리 적 위치로 인하여 내륙지역과는 다른 독특한 기후대를 형성 하고, 동해와는 다른 환경적 특징을 보이기 때문에 독도에 는 다른 곳엔 없는 희귀생물종이 많다. 그래서 학계에 보고 되지 않은 생물신종들이 발견돼 ‘독도’라는 지명이 담긴 국 제학명이 붙는 일도 잦은 편이다.
우리나라 해양세균의 종 다양성 및 분포에 대해서는 여러 학자들이 남해와 서해의 갯벌과 해저퇴적물에서 연구한 바 있지만, 생물지화학순환과 생물환경 정화에서 결정적인 역 할을 하는 만이나 해변 지역에서 미생물군집의 조성에 대해
*Corresponding author
Tel: 82-53-950-5374, Fax: 82-53-955-5522 E-mail: [email protected]
서는 거의 알려지지 않았다[15]. 특히, 독도 연안의 해양세 균의 분포에 대한 연구는 거의 없다. 그러므로, 본 연구에서 는 독도의 동도 연안과 선착장 주변, 서도 어민 숙소 근처에 서 2007년과 2008년에 3차례에 걸쳐 해수와 모래 등을 채 집하여 해양 세균을 분리한 후, 그 염색체 DNA를 추출하여 16S rRNA의 염기서열을 분석하여 부분동정을 통하여 독도 에 존재하는 해양미생물의 종류를 밝혔다. 동시에 독도에 서 식하는 배양가능한 세균의 우점속을 파악하며, 새로운 종 또 는 새로운 속으로의 가능성을 제시하고, 나아가 보다 유용 한 세균 자원 확보를 위한 기초 데이터를 제공하였다.
재료 및 방법 시료 채취 및 배양가능한 세균의 분리
독도를 구성하고 있는 2개의 큰 섬인 동도와 서도 연안에 서, 먼저 접근하기 쉬운 동도 계단 주위 연안을 중심으로 바 닷물을 채취하고, 선착장 주변에서 고여 있거나 흐르는 바 닷물과, 서도에서는 어민 숙소 앞과 물골 주위의 바닷물을 각각 채취하였다. 2007년 9월, 2008년 4월과 6월 등 3회에 걸쳐 채취하였다. 채취한 시료는 멸균된 cornical tube에 담 았고, 실험실에 옮겨서는 4oC에 보관하였다.
채취한 시료는 단계별로 멸균된 artificial sea water[4]로 희석하여 standard dilution plating technique을 이용하여 Marine agar 2216(MA; Difco, USA) 배지에 도말하였다 [41]. 1/10 MA, 1/2 MA, MA 배지 등에 희석한 시료를 각 각 100 µL 도말한 후, 25oC에서 5일간 배양하였다. 세균의 순수 분리를 위하여, 배지 상에 나타난 군락을 형태적으로 분류, 선별하여 각각의 한천 배지에 획선 도말하였다.
16S rRNA 유전자의 염기배열 결정
육안으로 관찰했을 때 콜로니의 모양, 크기, 색깔 등에 따 라 세균을 순수 분리하였다. 분리한 세균을 Marine broth (MB; Difco, USA)에 배양한 후 원심분리하여 세균을 모았 다(10,000×g, 5 min). QIAamp DNA mini kit(QIAGEN;
USA)을 사용하여 total genomic DNA를 추출하였다. 분리 한 total DNA에서 16S rRNA를 증폭시키기 위하여 5'- GAGTTTGATCATGGCTCAG-3'(Escherichia coli nucleo- tides 9에서 27)를 forward primer로 5-ACGGTTACCTTG- TTACGACTT-3'(E. coli nucleotides 1492에서 1512)를 reverse primer로 사용하여 PCR을 수행하였다. PCR 조건은 denaturation(94oC, 5분) 시킨 후, denaturation(94oC, 30초), annealing(60oC, 30초), elongation(72oC, 45초) 단계를 35회 반복하고, 마지막으로 elongation(72oC, 10분)하는 조건으로 반응시켰다[39]. PCR 산물을 DNA prep mateTM(Bioneer, Korea)를 사용하여 회수하고, 염기서열을 결정하였다.
16S rRNA 유전자의 염기서열에 대한 계통분석
분리된 세균을 동정하기 위해서 16S rRNA 유전자의 염 기서열 간 유사도를 확인하기 위하여 BLAST(Basic Local Alignment Search Tool) search를 이용하여 NCBI(National Center for Biotechnology Information)의 GenBank와 Ez- Taxon server(http://www.eztaxon.org)의 데이터베이스에서 유사한 염기서열을 비교하였다[27]. 분리균 DNA로부터 결 정된 염기서열과 데이터베이스에서 얻어진 염기서열은 PHYDIT program(http://plaza.snu.ac.kr/~jchun/phydit)을 이 용하여 Clustal W multiple alignment로 정렬하여 계통수 제 작에 이용하였다. 염기서열 간의 유전적 거리는 Jukes &
Cantor distance model을 사용하여 추정하였다[21].
Neighbour-joining data의 Bootstrap 분석 시 1,000회의 resampling을 적용하여 tree topology의 평가 기준으로 사용 하였고[12, 33], 이를 토대로 하여 phylogenetic tree를 결정 하였다.
결과 및 고찰
독도 연안의 해수로부터 배양 가능한 세균의 분리 독도 연안의 해수로부터 전체 163개의 분리균을 얻었다.
3~4차례에 걸친 순수 분리과정을 통해 육안으로 관찰하였을 때, 군락의 크기, 형태, 색깔 등의 차이를 보이는 개체를 선 별 분리하였다.
채취한 장소에 따라 세균 종류와 수적 차이가 있었는데, 흐르는 해수를 채취 했을 때 보다 고여 있는 해수 시료에서 더 다양한 종류의 세균이 분리되었다. MA 원래 조성을 사 용한 고체 배지에서보다 1/2 MA, 1/10 MA로 희석한 고체 배지에서 적은 숫자의 세균이 분리되었지만, 해당 배지에서 만 특징적으로 분리되는 세균도 있었다.
시료 채취의 시기에 따라 분리되는 세균의 수적 차이는 크지 않았다. 2007년 9월과 2008년 4월에 채취한 시료에서 는 모두 61개의 분리균을 얻었고, 2008년 6월에 채취한 시 료에서는 76개의 분리균을 획득하였다. 그러나 2007년 9월 시료에서와는 달리, 2008년 4월과 6월 시료에서는 한천 성 분 분해균이 발견된 것처럼 분리균의 종류에 있어서는 차이 는 있었다. 전체적으로 163종을 순수 분리하였지만, 미생물 은 환경적인 요소와 상호작용하면서 서식하므로 실험실에서 배양하여 순수 분리할 수 있는 미생물은 1% 미만이고, 더욱 이 MA 배지 한 가지만을 사용하여 분리하였기 때문에 실 제로 독도 연안에 존재하는 미생물의 종류나 숫자는 훨씬 많 을 것으로 추측할 수 있다[2, 30].
분리된 해양미생물의 부분동정과 채취시기 별, 채취시점 별 종 조성 분석
163개의 분리균에서 16S rRNA 유전자의 염기서열을 분 석할 수 있었는데, BLAST 로써 분석한 결과 56속 94종으
로 부분동정 되었다. 전체적으로 보면 94개의 분리균이 gamma-proteobacteria(57.7%), 32개의 분리균이 alpha-pro- teobacteria(19.6%), 27개의 분리균이 bacteriodetes(16.6%) 에 속하고 있었고, 그 외에 6개의 분리균이 low G+C Gram positive bacteria(3.7%) 4개의 분리균이 epsilon-proteobac- teria(2.5%)에 각각 속했다. Proteobacteria 중 강이나 호수 에서 주로 분리되는 beta-proteobacteria는 분리되지 않았고 [8, 13, 27, 29], 대부분의 해양 침적지에서 우점으로 분포되 는 delta-proteobacteria도 역시 분리되지 않았다. 강화 갯벌 의 세균군집 조사에서는 황 환원세균과 유연관계가 있는 delta-proteobacteria가 많이 분포되고 있음이 보고되었다[23].
독도 연안 해양세균의 분포를 살펴보면 proteobacteria와 bacteriodetes가 대다수 해양생태계에서 우점으로 존재한다 는 결과와 일치하였다[1, 10]. 특히 alpha-, gamma-proteo- bacteria가 일반적인 해양생태계에서 주로 우점으로 존재한 다는 결과와도 일치하였다[7, 14].
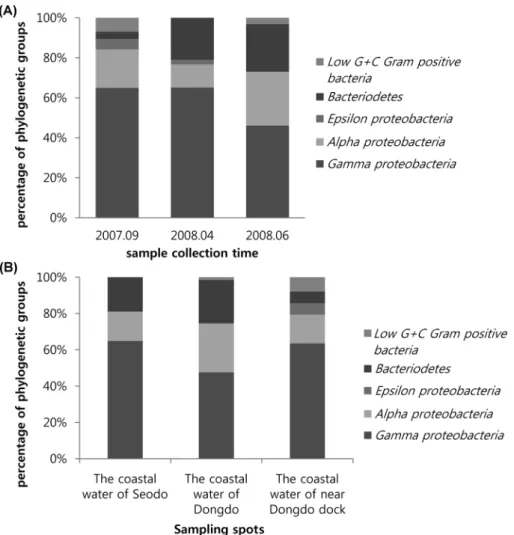
시료 채취 시기에 따른 세균 분포는 proteobacteria 중에 서도 gamma-proteobacteria가 모든 시기에서 가장 많이 분
포하고 있었지만, 2008년 6월 시료에서는 비교적 gamma- proteobacteria의 분포가 낮았던 대신에 alpha-proteobaceria 와 bacteriodetes의 분포가 높았다(Fig. 1A). 시료 채취 시점 에 따른 세균 분포는 gamma-proteobacteria가 모든 시점에 서 가장 많이 분포하고 있었지만, 동도 앞바다에서 채취한 시료에서는 다른 시점보다 alpha-proteobaceria의 분포 비율 이 높았고, Epsilon-proteobacteria는 동도 선착장 주변에서 채취한 시료에서만 분리되었으며, 동도 선착장 주변에서 채 취한 시료에서 가장 다양한 그룹에 속하는 세균이 분리되었 다(Fig. 1B).
Gamma-proteobacteria는 19속에 속하는 51종 94개의 분 리균이 부분 동정되었다. Vibrio 속이 33개로 동정되어 가장 우점속으로 나타났고(Fig. 2A), 세 번의 시료 채취시기 중 해수 온도가 가장 높았던 2007년 9월 시료에서는 사람과 어 류에 병을 유발한다고 보고되고 있는 V. harveyii, V.
alginolyticus와 상동성이 높은 udc311, udc324가 분리되기 도 하였다[11]. 두 번째로는 Shewanella 속이 15개로 동정되 었다. 2007년 9월 시료에서는 gamma-proteobacteria 중
Fig. 1. Comparison of the cultivable bacterial community groups based on 16S rRNA sequence analysis; (A) bacterial community groups as sample collection time (B) bacterial community groups as sampling spots.
Shewanella 속이 차지하는 비율이 약 22%로 Vibrio속보다 더 우점을 차지하고 있었다. 분리균 udc398은 S. japonica와 99.8%의 상동성을 가지고 있는데, S. japonica는 protease, lipase, amylase, agarase와 alginase 등과 같은 다양한 가수 분해효소와 eicosapentaenoic acid를 생산한다고 알려졌다[18, 19]. 분리균 udc398도 α-agarase 활성을 확인하였다.
Agarivorans 속은 2008년 4월 시료에서 6개의 분리균이 Agarivorans albus와 99.1~99.5%의 염기서열 상동성을 보였 다. 이들 분리균은 한천 성분을 분해하여 액체상태로 만드 는 β-agarase 활성이 있었다. Agarase는 한천 성분을 분해하 여 agar oligosaccharides를 생성하는데, agar oligosacchari- des는 항산화, 항세균 효과 등을 가지고 있어서 식품, 화장 품, 의료분야 등에서 다양하게 응용되고 있다[36].
2007년 9월 시료에서는 2008년 4월과 6월 시료보다는 더 다양한 gamma-proteobacteria에 속하는 균들 분리 동정되었 고, 4월과 6월 시료에서는 대장균군이 검출되지 않았지만, 9 월 시료에서는 동도 선착장 주변의 해수 시료에서 대장균군 에 속하는 세균인 Enterobacter aerogens가 검출되었는데, 이는 그 시기에 동도 선착장 인근 해역이 분변성 오염이 있 을 수 있음을 암시하고 있다.
EzTaxon server의 데이터베이스에서 16S rRNA 염기서열 상동성을 비교했을 때 상동성이 98.5% 이상인 분리균은 이 전에 분리 발표된 세균과 동일한 종으로 추정되었지만, 98.5% 미만인 분리균은 새로운 종일 가능성이 있다고 추정 되었다[38]. Gamma-proteobacteria로 분류된 94개의 분리균 중 16S rRNA 염기서열 상동성이 98.5% 미만인 분리균은 14개로 약 15%의 분리균이 새로운 종으로 분류될 가능성을 가진 균주로 추정되었다(Table 1). 2007년 9월 시료에서는 다른 시료보다 gamma-proteobacteria의 비율도 높았고, 다 양한 종류의 gamma-proteobacteria가 분리되었고, 새로운 종 으로 분류될 가능성을 가진 균주의 비율도 약 24%(9/37)로 2008년 4월 시료 약 14%(4/18), 6월 시료 약 3%(1/29)보다 높았다.
Alpha-proteobacteria는 17속에 속하는 22종 33개의 분리 균이 부분 동정되었다. Alpha-proteobacteria는 31개의 분 리균이 Rhodobacteriaceae과에 속했고, 2개의 분리균이 Sphingomonadaceae과에 속했다. 이 결과는 바닷물로부터 분 리한 alpha-proteobacter는 Rhodobacteriaceae과와 Sphin- gomonadaceae과가 우점을 차지하고 있다는 다른 지역의 분 석 결과와 일치하는 분포를 보이고 있다[33]. Rhodo- bacteriaceae과와 Sphingomonadaceae과 모두 해양 Roseo- bacter 계통(alpha-3 subclass)에 속하는데, Roseobacter 계통 은 특히 해양의 해안가와 극지방의 해양에서 배양할 수 미 생물 중 약 25%를 차지한다고 알려져 있다[35]. 독도의 해 안가에서 배양되는 미생물의 군집에서 분리된 alpha- proteobacteria는 모두 Roseobacter 계통으로 전체 염기서열 분석된 분리균의 약 20%(33/163)를 차지하고 있었다.
Roseobacter 계통은 호기성 비산소성 광합성을 담당하고, 온 실 가스인 일산화탄소의 산화를 할 수 있어서 지구의 탄소 와 황 순환에 중요한 역할을 하고 있다고 한다[35]. Alpha- proteobacteria는 10종이 분리된 Phaeobacter 속이 우점속을 차지하고 있었고 나머지 분리균들은 특별히 우점속을 차지 하지 않았고 다양한 alpha-proteobacteria에 속하는 분리균 들이 동정되었다(Fig. 2B). 분리균 2007년 9월 시료에서 분 리된 udc336과 udc337은 Sulfitobacter pontiacus로 동정되 었는데, Sulfitobacter pontiacus는 아황산염과 일산화탄소를 산화 할 수 있고, 방향족 화합물을 분해 할 수 있는 roseo- bacter 계통에 속하는 균주로 알려졌다[3]. 2008년 4월 시료 에서는 아황산염의 산화능력이 있고, 바다 양식에서 독성을 가지는 세균에 항균활성이 있다고 보고된 Phaeobacter 속이 Fig. 2 Composition of cultivable bacterial flora; (A) gamma-
proteobacteria, (B) alpha-proteobacteria, (C) bacteriodetes.
[32] 우점속을 차지하고 있었다. 2008년 6월 시료에서는 가 장 많은 17개의 alpha-proteobacteria가 분리되었다(Fig.
1A).
Alpha-proteobacteria로 분류된 33개의 분리균 중 16S rRNA 염기서열 상동성이 98.5% 미만인 분리균은 14개로 약 42%의 분리균이 새로운 종이 될 가능성을 가진 균주로 추정되었다(Table 1). 이 결과는 gamma-proteobacteria로
분류된 분리균보다 훨씬 새로운 종으로 동정될 가능성이 높 았다.
Epsilon-proteobacteria는 주로 동물의 소화관에서 분리 동 정되고 있고 환경에서 분리되는 epsilon-proteobacteria는 대 부분 심해저 화산 열수구나 냉용수 같은 극한 환경에서 다 수 분포하고 있음이 알려져 있다[5]. 우리나라 제주 해안 바 닷물에서 미생물군집 분포의 다양성을 조사했을 때도 epsilon- Table 1. Isolates exhibited below 98.5% sequence similarity of the coastal water bacteria to the nearest neighbor.
Site Isolated month
Isolate No.
Accession
No. Phylogenetic group Nearest neighbor based on 16S rRNA sequence
Similarity (%)
The coastal water of Dong do
2007.9 udc302 GQ245891 alpha proteobacteria Thalassococcus halodurans 95.67%
2007.9 udc303 GQ245892 alpha proteobacteria Ponticoccus litoralis 95.53%
2007.9 udc305 GQ245894 gamma proteobacteria Thalassomonas haliotis 96.45%
2007.9 udc306 GQ245895 gamma proteobacteria Neptuniibacter caesariensis 97.89%
2007.9 udc315 GQ245904 alpha proteobacteria Celeribacter neptunius 96.04%
2007.9 udc316 GQ245905 epsilon proteobacteria Arcobacter nitrofigilis 95.51%
2007.9 udc317 GQ245906 gamma proteobacteria Marinobacterium sediminicola 98.16%
2007.9 udc319 GQ245908 alpha proteobacteria Celeribacter neptunius 96.75%
2007.9 udc329 GQ245918 gamma proteobacteria Shewanella fodinae 97.89%
2008,4 udc402 HM031991 gamma proteobacteria Vibrio tapetis 94.87%
2008.4 udc406 HM031993 gamma proteobacteria Vibrio natriegens 94.73%
2008.4 udc409 HM031995 alpha proteobacteria Phaeobacter daeponensis 98.23%
2008.6 udc452 HM032005 alpha proteobacteria Phaeobacter inhibens 97.31%
2008.6 udc455 HM032006 alpha proteobacteria Thalassobius aestuarii 96.98%
2008.6 udc459 HM032007 alpha proteobacteria Octadecabacter antarcticus 97.51%
2008.6 udc470 HM032009 bacteriodetes Arenibacter echinorum 98.14%
2008.6 udc475 HM032010 alpha proteobacteria Tateyamaria omphalii 97.70%
2008.6 udc479 HM032011 alpha proteobacteria Nereida ignava 97.19%
2008.6 udc481 HM032012 alpha proteobacteria Maribius pelagius 94.60%
2008.6 udc482 HM032013 alpha proteobacteria Jannaschia donghaensis 95.70%
2008.6 udc483 HM032014 alpha proteobacteria Octadecabacter antarcticus 97.42%
The coastal water of
near Dong do
dock
2007.9 udc309 GQ245898 gamma proteobacteria Photobacterium leiognathi 97.56%
2007.9 udc310 GQ245899 gamma proteobacteria Photobacterium aquimaris 97.77%
2007.9 udc312 GQ245901 epsilon proteobacteria Arcobacter nitrofigilis 95.67%
2007.9 udc313 GQ245902 epsilon proteobacteria Arcobacter nitrofigilis 94.93%
2007.9 udc339 GQ256628 alpha proteobacteria Seohaeicola saemankumensis 97.75%
2007.9 udc342 GQ246631 gamma proteobacteria Halomonas meridiana 98.35%
2007.9 udc343 GQ246632 gamma proteobacteria Pseudomonas argentinensis 98.10%
2007.9 udc347 GQ246635 alpha proteobacteria Catellibacterium aquatile 97.84%
2007.9 udc353 GQ246641 bacteriodetes Salinimicrobium xinjiangense 98.35%
2007.9 udc354 GQ246642 gamma proteobacteria Bowmanella pacifica 94.64%
2008.4 udc415 HM031998 epsilon proteobacteria Arcobacter nitrofigilis 95.66%
2008.4 udc418 HM031999 alpha proteobacteria Jannaschia donghaensis 96.41%
2008.4 udc421 HM032000 bacteriodetes Polaribacter dokdonensis 96.26%
2008.6 udc499 HM032018 gamma proteobacteria Vibrio tapetis 94.41%
The coastal water of
Seo do
2007.9 udc363 GQ246649 alpha proteobacteria Phaeobacter gallaeciensis 97.50%
2008.4 udc369 HM031970 bacteriodetes Cytophaga fermentans 93.30%
2008.4 udc377 HM031972 bacteriodetes Lutibacter litoralis 94.84%
2008.4 udc385 HM031979 bacteriodetes Cytophaga fermentans 93.04%
2008.4 udc392 HM031984 gamma proteobacteria Vibrio sinaloensis 95.01%
2008.4 udc395 HM031985 gamma proteobacteria Shewanella gaetbuli 98.45%
proteobacteria는 분리되지 않았다[26]. 2007년 9월 시료와 4 월 시료에서 Campylobacteraceae과에 속하는 미산소성이고 질소고정능력을 가지는 세균인 Arcobacter nitrofigilis와 16S rRNA 염기서열 상동성이 94.6~95.7%인 4개가 분리되었지 만 분리종간에 상동성은 94~98%로 각각이 새로운 종이 될 가능성이 있다고 추정되었다[38]. Arcobacter nitrofigilis는 Vandamme 등[28, 36]에 의해 형태적 생화학적 특성에 따라 Campylobacter nitrofigilis에서 학명이 바뀐 세균으로 식물 뿌리에 분리된 세균이다. 이전까지 독도에서 분리되어서 새 로운 종으로 등록된 미생물 중에 epsilon-proteobacteria에
속하는 종은 없었기 때문에 다양한 생리화학적 조사를 통하 여 잠재적인 새로운 종으로 분리할 수 있었다.
Bacteriodetes에 속하는 세균은 26개의 분리균이 14속 14 종으로 부분동정 되었다. Bacteriodetes에 속하는 균은 대부 분 호기성이며 그람 음성균으로 complex macromolecule을 분해할 수 있는 능력을 가지는 균이 다수 알려져 있다[17].
7개의 분리균이 Cellulophaga 속으로 동정되어 가장 우점속 을 차지하고 있었고, 그 다음 Maribacter 속이 5개가 분리 되었다(Fig. 2C).
2008년 4월 시료에서 분리된 udc421은 2006년 윤 등[42]
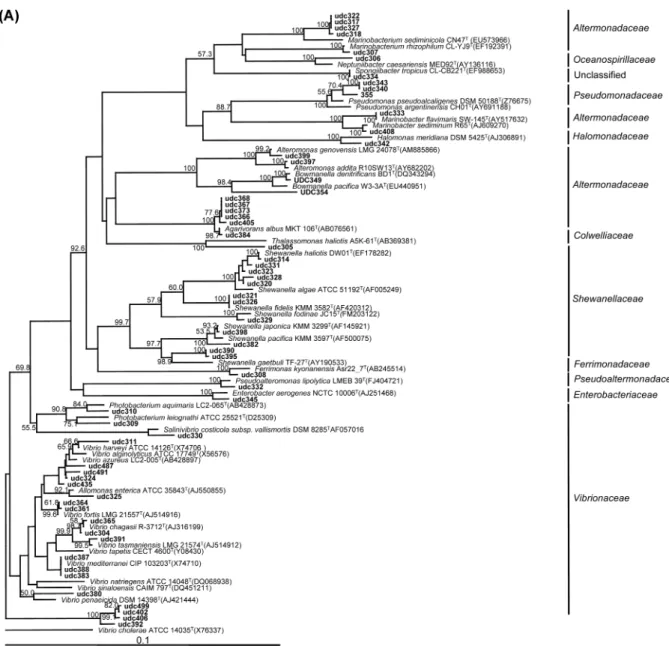
Fig. 3. Phylogenetic tree based on comparison of the 16S rDNA gene sequences of cultivable bacteria isolated from Dokdo coastal sea water and some of their closest phylogenetic relatives. The phylogenetic tree was generated using the neighbor-joining method.
Bootstrap values higher than 50 % are given at branching points. Bold type indicates bacterial isolates in this study. The bar represents 0.1 estimated sequence divergence. (A) Phylogenetic relationships of bacterial 16S rRNA sequences within gamma-proteobacteria. Vibrio chol- erae (X76337) served as the out group. (B) Phylogenetic relationships of the bacterial 16S rRNA sequences within alpha-, epsilon-proteo- bacteria. Hydrogenimonas thermophilus (AB105049) served as the out group. (C) Phylogenetic relationships of the bacterial 16S rRNA sequences within bacteriodetes. Flexibacter flexilis (M62794) served as the out group.
에 의해 독도 바닷물에서 분리한 Flavobacteriaceae과의 Polaribacter dokdonensis와 96.3%의 상동성을 가지고 있었 는데, 독도에서 분리한 Polaribacter속의 또 다른 종으로 동 정될 수 있다고 추정되었다. 2008년 6월 동도 연안 바닷물 에서 분리한 다른 5개의 분리균은 2005년 윤 등[40]에 의해 분리된 독도의 바닷물에서 분리된 Maribacter dokdonensis 와 각각 98.7~99.7%의 비교적 높은 상동성을 보이고 있었다.
Bacteridetes로 분류된 26개의 분리균 중 16S rRNA 염기 서열 상동성이 98.5% 미만인 분리균은 8개로 약 31%의 분 리균이 새로운 종이 될 가능성을 가진 균주로 추정되었다 (Table 1). 2008년 4월 시료에서는 9개의 Bacteriodetes에 속 하는 종 중에 4종(약 44%)이 신종이나 신속으로 동정될 수 있다고 추정되기 때문에, 2008년 6월 시료보다 다양성도 높 았고, 신종이나 신속의 가능성을 가진 균주의 비율도 높았 다(Table 1).
2007년 9월 시료와 2008년 6월 시료에서는 low G+C Gram positive bacteria도 몇 종류가 분리되었다(Fig 1A). 균 의 다양성을 보면 육안으로 볼 때 다르다고 분리했지만, 실
제로 중복으로 분리되는 경우도 있었다. 하지만 종 분포의 다양성은 매우 높았다. 분리균의 종 분포를 보면 2007년 9 월에 분리한 균들이 2008년 4월이나 6월에 분리한 균과 차 이가 많았다. 이는 해수온도의 차이 때문에 세균 분포가 다 를 수 있다고 추정할 수 있었다[31]. 그리고 분리균의 종을 살펴보면 지리적으로 가까이 위치한 한국의 다른 해안가나 독도에서 분리된 균들과 염기서열 상동성이 높은 균들이 다 른 지역에서 분리한 균보다는 많은 비율을 차지하고 있었다.
전체 염기서열이 분석된 분리균 163개 중 16S rRNA 염 기서열이 기존 세균의 염기서열과 상동성을 비교했을 때, 98.5% 이하인 균주는 gamma-proteobacteria 14개, alpha- proteobacteria 14개, Bacteriodetes 8개로 전체 36개(약 22%)의 분리균이 새로운 종이 될 가능성을 가진 균주로 추 정되었고, alpha-proteobacteria로 분류된 분리균이 새로운 종으로 분류될 가능성이 가장 높았다. 이 결과는 전 등[20]
에 의해 연구된 독도 식물근권에서 분리한 포자형성세균과 질소고정세균의 신종 가능성을 가진 균주의 비율인 20%보 다 약간 높았다.
Fig. 3. Continued.
독도 해양세균의 계통분석학적 다양성
염기서열이 분석된 분리균 163종 중 16S rRNA 유전자 염기서열의 거의 전체가 분석된 107종을 대상으로 각각의 분리균을 크게 gamma-proteobacteria와 alpha-, epsilon- prteobacteria와 Bacteriodetes 그룹으로 분류한 뒤 계통수를 작성하였다. Fig. 3A는 가장 많이 분리된 gamma-proteo- bacteria의 phylogenetic tree를 보여준다. Vibrionaceae과, Shewanellaceae과와 Altermonadaceae과가 다수를 차지하고 있었고, 그 외에 10과와 19속에 속하는 다양한 세균이 분리 되었다. Fig. 3B는 alpha-proteobacteria와 epsilon-proteo- bacteria의 phylogenetic tree를 보여준다. Alpha-proteobac- teria의 대부분이 Rhodobacteriaceae과에 속했고, 한 종의 Sphingomonadaceae과에 속하는 세균이 분리되었다. Epsilon- proteobacteria는 한 종이 분리되었지만, 각각의 상동성이 낮 아서 다른 종으로 분리될 수 있다. Fig. 3C는 Bacteriodetes 의 phylogenetic tree를 보여주고 있다. 대부분 Flavobac-
teriaceae과에 속했고 그 외에 Flammeovirgaceae과, Cyclo- bacteriaceae과와 Flexibacteriaceae과에 속하는 다양한 세균 이 분리되었다. 독도의 해수에서 분리한 균은 gamma-, alpha-, epsilon-proteobacteria, Bacteriodetes, low G+C Gram positive bacteria 등 5개의 계통으로 나누어졌다. 그 중에 gamma-proteobacteria에 속하는 균이 약 58%를 차지 하고 있었고, high G+C Gram positive bacteria는 분리되지 않았다. 이전에 한국의 해수환경인 제주 문섬의 세균군집 조 사에서는 high G+C Gram positive bacteria가 약 59%가 분리되었고, 그 다음으로는 alpha-proteobacteria가 분리되었 다는 결과와는 큰 차이를 보이고 있었다[26]. 독도 미생물의 군집분포는 한국의 다른 해수환경과는 상당히 다른 양상의 군집분포를 보이고 있었고[22, 23, 24, 25], 매우 다양한 미 생물이 분포하고 있으면서, 신종후보로 동정된 균주의 분포 가 높았다. 이는 독도의 해양이 비교적 다른 환경조건을 가 지고 있으며, 동시에 인간에 의한 오염이 비교적 적도록 잘 Fig. 3. Continued.
보존된 환경이기 때문이라고 추정할 수 있다.
요 약
독도 연안에 존재하는 배양 가능한 미생물의 다양성을 16S rRNA 분석으로 조사하였다. 동도 선착장 주변과 서도 숙소 부근을 중심으로 채취한 시료에서 163개의 해양 미생물을 분리하였다. 분리한 미생물 163종을 16S rRNA 염기서열 의 분석을 이용하여 부분동정 할 수 있었다. 부분동정된 미 생물은 gamma-proteobacteria(58%), alpha-proteobacteria (20%), bacteriodetes(16%) 계통이 대부분을 차지하고 있었 고, 그 외에도 low G+C Gram positive bacteria와 epsilon- proteobacteria가 소수 동정 되었다. 염기서열이 분석된 미생 물들은 이전에 보고된 미생물들의 16S rRNA 유전자와 93.3%에서 100%의 유사도를 보이며 56속 94종으로 부분 동 정되었다. 163종의 부분 동정된 미생물 중 36개의 분리 미 생물이 새로운 종으로 분류될 후보군으로 추정되었다. 본 연 구의 결과 독도연안 바닷물에는 proteobacteria와 bacterio- detes의 비율이 높게 나타났고, 미생물 다양성을 높게 유지 하고 있었다. 이 다양한 미생물로부터 다양한 유용미생물 자 원을 확보할 수 있고, 새로운 종으로 분류될 후보군 들은 추 후 여러 생리생화학적 실험을 수행하여 새로운 종 또는 새 로운 속으로 발표할 수 있을 것으로 판단된다.
REFERENCES
1. Alfreider, A., J. Pernthhaler, R. Amann, B. Sattler, F. O.
GlÖckner, A. Wille, and R. Psenner. 1996. Community analysis of the bacterial assemblages in the winter cover and pelagic layers of high mountain lake by in situ hybridization.
Appl. Environ. Microbiol. 62: 2138-2144.
2. Amann, R. I., W. Ludwig, and K. H. Schleifer. 1995.
Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 59: 143- 169.
3. Buchan, A., J. M. González, and M. A. Moran. 2005.
Overview of the marine Roseobacter lineage. Appl. Environ.
Microbiol. 71: 5665-5677.
4. Burns, A., M. Roche, and L. Berthe-Corti. 2001. Muricauda ruestringensis gen. nov., sp. nov., a facultatively anaerobic, appendaged bacterium from German North sea intertidal sediment. Int. J. Syst. Evol. Microbiol. 51: 1997-2006.
5. Campbell, B. J., A. S. Engel, M. L. Porter, and K. Takai.
2006. The versatile ε -proteobacteria: key players in sulphidic habitats. Nat. Rev. Microbiol. 4: 458-468.
6. Cho B. C. and F. Azan. 1988. Major role of bacteria in biogeochemical fluxes in the ocean's interior. Nature 332:
441-443.
7. Cottrell, M. T. and D. L. Kirchman. 2000. Community composition of marine bacterioplankton determined by 16S
rRNA gene clone libraries and fluorescence in situ hybridization. Appl. Environ, Microbiol. 66: 5116-5122.
8. Cottrell, M. T. and D. L. Kirchman. 2000. Natural assem- blages of marine proteobacteria and members Cytophaga- Flavobacter cluster consuming low- and high-molecular- weight dissolved organic matter. Appl. Environ. Microbiol.
66: 1692-1697.
9. Ducklow H. W., A. P. Duncan, P. J. L. Williams, and M. D.
John. 1986. Bacterioplankton: a sink for carbon in a coastal marine plankton community, Science 232: 865-867.
10. Eilers, H., J. Pernthaler, F. O. GlÖckner, and R. Aman. 2000.
Culturability and in situ abundance of pelagic bacteria from the North Sea. Appl. Environ, Microbiol. 66: 3044-3051.
11. Fabiano, L. Thompson, I. Tetsuya, and S. Jean. 2004. Bio- diversity of Vibrios. Microbiol. Mol. Biol. Rev. 68: 403-431.
12. Felsenstein, J. 1985. Confidence limits on phylogenesis: an approach using the bootstrap. Evolution 39: 783-791.
13. GlÖckner, F. O., B. M. Fuchs, and R. Aman. 1999. Bacterio- plankton compositions of lakesand oceans: a first compar- ison based on fluorescence- in situ-hybridization. Appl.
Environ. Microbiol. 65: 3721-3726.
14. Gonzalez, J. M., and M. A. Moran. 1997. Numerical domin- ance of a group of marine bacteria in the α-subclass of the class Proteobacteria in coastal seawater. Appl. Environ.
Microbiol. 63: 4237-4242.
15. Gonzalez, J. M., W. B. Whitman, R. E. Hodson, and M. A.
Moran. 1996. Identifying numerically abundant culturable bacteria from complex communities: an example from a lignin enrichment culture. Apple. Environ. Microbiol. 62:
4433-4440.
16. Gong, Y. and S. J. Son. 1982. A study of oceanic thermal fronts in the Southern Japan Sea. Bull. Fish. Res. Dev.
Agency 28: 25-54.
17. Holmes, B. 1991. The genera Flavobacterium, Sphingobac- terium, and Weeksella, In A. Balows, H. G. Trüper, M.
Dworkin, W. Harder, and K.-H. Schleifer (eds.). The pro- karyotes. Springer-Verlag, Berlin, Germany. pp. 3620-3630.
18. Ivanova, E. P., T. Sawabe, N. M. Gorshkova, V. I. Svetashev, V. V. Mikhailov, D. V. Nicolau, and R. Christen. 2001.
Shewanella japonica sp. nov. Int. J. Syst. Evol. Microbiol.
51: 1027-1033.
19. Ivanova, E. P., T. Sawabe, N. V. Zhukova, N. M. Gorshkova, O. I. Nedashkovskaya, K. Hayashi, G. M. Frolova, A. F.
Sergeev, K. G. Pavel, V. V. Mikhailov, and D. V. Nicolau.
2003. Occurrence and diversity of mesophilic Shewanella strains isolated from the North-West Pacific Ocean. Syst.
Appl. Microbiol. 26: 293-301.
20. Jeon, S. A., H. R. Sung, Y. M. Park, J. H. Park, and S.- Y.
Ghim. 2009. Analysis of endospore-forming bacteria or nitrogen-fixing bacteria community isolated from plants rhizosphere in Dokdo island. Kor. J. Microbiol. Biotechnol.
37: 189-196.
21. Jukes, T. H. and C. R. Cantor. 1969. Evolution of protein molecules. In Mammalian protein metabolism, (Munro, H.
N. eds.) pp. 21-132. New York, Academic Press, USA.
22. Kang H. and D. S. Kang. 2002. Contribution of marine microbes to particulate organic matter in the Korea Strait. J.
Kor. Soc. Oceanogr. 1: 35-44.
23. Kim, B. S., H. M. Oh, H. J. Kang, S. S. Park, and J. S.
Chun. 2004. Remarkable bacterial diversity in the tidal flat sediment as revealed by 16S rDNA analysis. J. Microbiol.
Biotechnol. 14: 205-211.
24. Lee, D. S., Y. S. Kim, S. Y. Jeong, C. K. Kang, and W. J.
Lee. 2008. Environmental characteristics and distributions of marine bacteria in the surface sediments of Kamak Bay in winter and summer. J. Environ. Sciences 17: 755-765.
25. Lee, J. B., M. S. Han and H. S. Yang. 1998. The ecosystem of the southern coastal waters of the East Sea, Korea 1.
Phytoplankton community structure and primary produc- tivity in September, 1994. J. Korean Fish . Soc. 31: 45-55.
26. Lee, J. H., H. H. Shin, D. S. Lee, K. K. Kwon, S. J. Kim, and H. K. Lim. 1999. Bacterial diversity of cultivable isolates from sea water and a marine coral, Plexauridae sp.
near Mun-Sum, Cheju-Island. J. Microbiol. 37: 193-199.
27. Maidak, B. L., J. R. Cole, T. G. Parker, CT Jr., P. R. Saxman, J. M. Stredwick, G. M. Garrity, B. Li, G. H. Olsen, S.
Paranik, T. M. Schmidt, and J. M. Tiedje. 2000. The RDP (Ribosomal Database project) continues. Nucleic acids Res.
28: 173-174.
28. McClung, C. R., D. G. Patriquin, and R. E. Davis. 1983.
Campylobacter nitrofigilis sp. nov., a nitrogen-fixing bac- terium associated with roots of Spartina alternijlora Loisel.
Int. J. Syst. Bacteriol. 33: 605-612.
29. MEthé, B. A., W. D. Hiorns, and J. P. Zehr. 1998. Contrasts between marine and freshwater bacterial community com- position: analyses of communities in Lake George and six other Adirondack lakes. Limnol. Oceanogr. 43: 368-374.
30. Olsen, G. J., D. J. Lane, S. J. Giovannoni, and N. R. Pace.
1986. Microbial ecology and evolution: a ribosomal RNA approach. Ann. Rev. Microbiol. 40: 337-365.
31. Pernthaler, J., F. O. GlÖckner, S. Unterholzner, A. Alfreider, R. Psenner, and R. Amann. 1998. Seasonal community and population dynamics of pelagic bacteria and archaea in a high mountain lake. Appl. Environ, Microbiol. 64: 4299- 4306.
32. Prado, S., J. Montes, J. L. Romalde, and J. L. Barja. 2009.
Inhibitory activity of Phaeobacter strains against aquacul-
ture pathogenic bacteria. Int. Microbiol. 12: 107-114.
33. Saitou, N. and M. Nei. 1987. The neighbor-joining method:
a new method for reconstructing phylogenetic trees. Mol.
Biol. Evol. 4: 406-425.
34. Vandamme, P., E . Falsen, R. Rossau, B. Hoste, P. Segers, R.
Tytgat, J. De Ley. 1991. Revision of Campylobacter, Helico- bacter, and Wolinella taxonomy: emendation of generic descriptions and proposal of Arcobacter gen. nov. Int. J.
Syst. Evol. Microbiol. 41: 88-103.
35. Wagner-Döbler, I. and H. Biebl. 2006. Environmental biology of the marine Roseobacter lineage. Annu. Rev.
Microbiol. 60: 255-280.
36. Wang J, X. Jiang, X. Mou, and H. Guan, 2004. Anti- oxidation of agar oligosaccharides produced by agarase from a marine bacterium. J. Appl. Phycol. 16: 333-340.
37. Wimpenny, J. W., W. L. T. Robert, and C. Philip. 1983.
Laboratory model systems for the investigation of spatially and temporally organized microbial ecosystems. In microbes in their natural environments. Cambridge Univ. Press. pp.
67-117.
38. Woo., P. C. Y., S. K. P. Lau, J. L. L . Teng, H. Tse, and K. Y.
Yuen. 2008 Then and now : use of 16S rDNA gene se- quencing for bacterial identification and discovery of novel bacteria in clinical microbiology laboratories. C.M.I. 14:
908-934.
39. Yoon, J. H., S. T. Lee, and Y. H. Park. 1998. Inter- and intraspecific phylogenetic analysis of the genus Norcar- dioides and related taxa based on 16s rRNA gene sequences.
Int. J. Syst. Bacteriol. 48: 187-194.
40. Yoon, J. H., K. H. Kang, S. Y. Lee, C. H. Lee and T. K. Oh.
2005. Maribacter dokdonensis sp. nov., isolated from sea water off a Korean island, Dokdo. Int. J. Syst. Evol.
Microbiol. 54: 487-491.
41. Yoon, J. H., S. J. Kang, S. Y. Lee, M. H. Lee, and T. K. Oh.
2005. Virgibacillus dokdonensis sp. nov., isolated from a Korean island, Dokdo, located at the edge of the East Sea in Korea. Int. J. Syst. Evol. Microbiol. 51: 1079-1086.
42. Yoon, J. H., S. J. Kang, and T. K. Oh. 2006. Polaribacter dokdonensis sp. nov., isolated from seawater. Int. J. Syst.
Evol. Microbiol. 56: 1251-1255.
(Received June 9, 2010/Accepted August 24, 2010)