248 책임저자:최영현, 614-052, 부산시 진구 양정동 산 45번지
동의대학교 한의과대학 생화학교실 Tel: 051-850-7413, Fax: 051-853-4036 E-mail: [email protected]
접수일: 2010년 5월 26일, 1차수정일: 2010년 8월 31일, 2차수정일: 2010년 9월 3일, 게재승인일: 2010년 9월 7일
Correspondence to:Yung Hyun Choi
Department of Biochemistry, Dongeui University College of Oriental Medicine, San 45, Yangjung-dong, Busanjin-gu, Busan 614-052, Korea Tel: +82-51-850-7413, Fax: +82-51-853-4036
E-mail: [email protected]
U937 인체혈구암세포에서 발효 아가콩 에탄올 추출물에 의한 Apoptosis 유도
1동아대학교 생명공학과 및 대학원 의생명과학과, 동의대학교 2대학원
바이오물질제어학과 및 블루바이오 소재개발센터, 3한의과대학 생화학교실 및 한의학연구소
이혜현1ㆍ한민호2ㆍ김민정1ㆍ서민정1ㆍ정영기1ㆍ최영현2,3
Apoptosis Induction of U937 Human Leukemia Cells by the Ethanol Extracts of Fermented Agabeans
Hye Hyeon Lee1, Min Ho Han2, Min Jeong Kim1, Min Jeong Seo1, Yong Kee Jeong1 and Yung Hyun Choi2,3
1Department of Medical Bioscience, Graduate School and Department of Biotechnology, College of Natural Resources and Life Science, Dong-A University, Busan 604-714, 2Department of Biomaterial Control, Graduate School and Blue-Bio Industry Regional Innovation Center, 3Department of Biochemistry, College of Oriental Medicineand Research Institute of
Oriental Medicine, Dongeui University, Busan 614-714, Korea
We previously reported that the extracts of fermented beans exhibits strong anti-inflammatory properties by suppressing the transcription of pro-inflammatory cytokine genes through the NF-κB signaling pathway however, the mechanisms of their cytoxicities in human cancer cells are poorly understood. In this study, we investigated further possible mechanisms by which fermented beans including soybean (FS), black agabean (FBA) and yellow agabean (FYA) exert their anti-cancer actions in cultured U937 human leukemia cells. Treatment with FS, FBA and FYA in U937 cells resulted in inhibition of growth and induction of apoptosis in a concentration-dependent manner, as measured by MTT assay, fluorescence microscopy, agrose gel electrophoresis and flow cytometry analysis. The increase in apoptosis was associated with up-regulation of pro-apoptotic Bax and Bad expression, down-regulation of anti-apoptotic Bcl-2 and Bcl-xL expression, and activation of caspases (-3, -8 and -9) and degradation of poly (ADP-ribose) polymerase protein. Furthermore, exposure of FS, FBA and FYA induced the expression of Fas, Fas ligand (FasL), Fas-associated protein with death domain (FADD) and TNFRSF1A-associated via death domain (TRADD). Taken together, our data indicate that the extracts of fermented beans exhibit anti-cancer properties by induction of apoptosis through a signaling cascade of death receptor-mediated extrinsic and mitochondria-mediated intrinsic caspase pathways. (Cancer Prev Res 15, 248-256, 2010) Key Words: Fermented beans, Apoptosis, Bcl-2, Caspase
서 론
세포의 여러 가지 활동 중 세포의 죽음은 크게
apoptosis와 necrosis로 구분될 수 있으며, 이는 세포의 형 태적 및 생화학적 특성에 의하여 구분된다. 직접적으로 독성이 있거나 물리적 상해 등 갑작스런 외부의 환경에 의해 유발되는 수동적 죽음인 necrosis와는 달리 apoptosis
는 능동적인 세포사멸 과정으로 세포의 부피 감소, 세포 막의 파괴, 세포질 및 염색질 응축, 세포막 수포화 현상, DNA 단편화 등의 현상이 수반되는데 이러한 현상은 세 포 내부의 정교한 신호전달에 의하여 조절되며, 생체의 발생 및 항상성을 유지에 매우 중요하게 작용한다.
Apoptosis 유발은 death receptor 관련 인자들, Bcl-2 및 inhibitor of apoptosis (IAP) family 인자들, 그리고 caspase 등 의 유전자들이 관여하며, 이는 크게 intrinsic와 extrinsic 2 가지의 경로를 통하여 일어난다.1,2) Intrinsic pathway는 여 러 가지 외부 자극에 의하여 mitochondrial membrane permeability의 변화가 유발되어 mitochondria 내에 존재하 는 여러 종류의 apoptotic molecule들이 세포질로 방출됨 으로써 시작되며, 그와 달리 extrinsic pathway는 mito- chondria 비의존적인 apoptosis 과정으로 apoptotic ligand가 death receptor와 결합하여 death-inducing signaling complex (DISC)를 형성한 후 caspase-8과 caspase-3을 활성화 시킨 다.3,4) 이들은 최종적으로 effector moleculer로써 apoptosis 유발에 중요한 역할을 한다고 알려진 caspases를 활성화 시키는데,5,6) 이 중 caspase-3은 직접적인 효과를 가지는 caspase로서, caspase-3의 활성화는 직ㆍ간접적으로 poly (ADP-ribose) polymerase (PARP) 등과 같은 많은 표적 단백 질의 분해에 관여한다.7) 이러한 기작을 통해 이루어지는 apotposis는 암 치료에 효과적인 방법 중 하나로 알려져 있으며, 최근 apoptosis 유발에 의한 암치료법의 중요성이 강조되고 있다.
인류에게 식물성 단백질을 공급원뿐만 아니라 수많은 화학제품의 원료로도 사용되는 콩은 여러 가지 방법으 로 조리되어 섭취되어지고 있는데, 특히 미생물을 이용 하여 발효를 시키는 방법 중에는 대표적으로 청국장을 들 수가 있다. 고초균(Bacillus subtilis)에 의해 발효되는 청 국장은 다른 아시아 지역에서는 “Natto”, “Tempeh”,
“Douchi”이라고도 불리우며, 된장이나 간장과 같이 콩을 이용한 다른 발효식품과는 다르게 짧은 발효시간을 가 진다는 것과 발효기간 동안 소금이 필요 하지 않다는 것 이 큰 특징이다.8,9) 청국장은 항산화, 항균, 혈압조절 및 항당뇨 등과 같은 여러 가지 효과를 가진다고 보고되어 지고 있으며, 이러한 청국장의 구성물 중 가장 풍부하게 존재하는 phytochemcal에는 여성 호르몬인 estrogen과 비 슷한 heterocyclic diphenolic 구조를 가지는 isoflavone이 있 다. 이러한 isoflavone과 isoflavone이 풍부한 식품은 산화적 손상과 관련이 있는 유방암과 전립선암 등과 같은 다양 한 종류의 암과 동맥경화증 등과 같은 각종 성인병으로 부터 예방하는 효과를 가지는 것으로 알려져 있다.10∼12) 한편 경북대학교에서 품종 육성되어진 아가콩 3호(노란
아가콩) 및 4호(검정 아가콩)는 isoflavone의 함량이 다른 콩들에 비해 매우 높은 것으로 확인 되어졌으므로 다양 한 생리활성에서 아가콩의 효과가 다른 콩에 비해 더욱 높을 것이라 생각된다.13,14) 최근 본 연구팀에서는 대두, 아가콩 3호 및 4호의 발효산물에 의한 항염증효능을 조 사한 결과, phorbol 12-myristate 13-acetate 처리에 의해 유 도된 COX-2의 발현, prostaglandin E2의 생성 및 NF-κB의 활성 증가가 아가콩 추출물에 의하여 유의적으로 억제 됨을 보고한 바 있으나,15) 이들 추출물에 의한 항암 효과 는 보고된 바 없다. 따라서 본 연구에서는 대두, 아가콩 3호, 4호의 발효추출물의 항암활성 기전을 조사하기 위 하여 발효콩 추출물 처리에 의한 인체혈구암세포의 성 장억제 정도와 이로 인한 세포 및 핵의 형태적 변화를 조사하였고, apoptosis 유발의 가능성을 확인하였으며, apoptosis 조절에 중요한 몇 가지 유전자 산물의 발현 변 화를 조사함으로써 어떠한 경로를 통하여 apoptosis가 유 발되는지 조사하였다.
재료 및 방법 1. 세포 배양
실험에 사용한 인체 혈구암세포 U937은 한국생명공 학연구소(KRIBB, Taejeon, Korea)에서 분주 받아 사용하였 으며, 세포의 배양을 위해 10% fetal bovime serum (FBS, Gibco BRL, Grand Island, NY, USA) 및 1%의 penicillin 및 streptomycin (Gibco BRL)이 포함된 RPMI-1640 배지를 (Gibco BRL) 사용하여 37oC, 5% CO2 조건 하에서 배양하 였다.
2. 시료 준비 및 처리
본 실험에 사용된 시료는 시중에서 판매되는 대두와 경북대학교에서 품종 육성되어진 아가콩 3호(노란 아가 콩) 및 4호(검정 아가콩)를 제공받아 사용하였다. 시료 준비를 위해 콩들을 수세하고 24시간 침지시킨 뒤 가압 증자한 후 실험실에서 따로 분리되어진 Bacillus sp. sm26 을 50 g당 1 ml 접종하여 37oC, 72시간 배양하였고, 그 후 70% EtOH로 추출[extracts of fermented soybean (FS), black agabean (FBA) and yellow agabean (FYA)]하여 필터로 불순물을 제거한 후 농축시켜 사용하였다.15)
3. MTT assay를 이용한 세포 생존율 조사 및 세포 형 태변화 관찰
FS, FBA 및 FYA의 처리에 따른 생존율 비교를 위하여 6 well plate에 U937 세포를 1×105 개/ml로 분주하고 FS,
FBA 및 FYA를 처리한 후 48시간 동안 배양하였다. 시간 경과 후 배지에 tetrazolium bromide salt (MTT, Amersco, Solon, Ohio, USA) 시약을 0.5 mg/ml 농도로 각 well에 분주 하고 2시간 동안 배양하였다. 배양이 끝난 후 MTT 시약 을 깨끗하게 제거하고 dimethylsulfoxide (DMSO, Amersco) 를 1 ml씩 분주하여 well에 생성된 formazan을 모두 녹인 후 96 well plate에 200μl씩 옮겨서 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)로 540 nm에서 흡광도를 측 정하였다. 또한 FS, FBA 및 FYA를 농도별로 48시간 처리 하여 배양하였을 때 세포의 전체적인 변화를 관찰하기 위하여 도립현미경(inverted microscope, Carl Zeiss, Germany)을 이용하여 200배의 배율로 각 농도에 따른 세 포의 형태 변화를 관찰하였다.
4. DAPI 염색에 의한 세포핵의 형태 관찰
Apoptosis 유발 시 특이적으로 나타나는 핵의 형태변화 인 염색질 응축 현상의 유무를 관찰하기 위하여 FS, FBA 및 FYA를 처리한 세포를 모은 다음 37% formaldehyde 용 액과 phosphate buffered saline (PBS)를 1:9의 비율로 섞은 fixing solution으로 세포를 상온에서 10분 동안 고정하고 4,6-diamidino-2-phenylindole (DAPI, Sigma) 용액을 이용하 여 상온에서 암조건으로 15분간 염색시켰다. 염색된 세 포를 PBS 및 증류수로 세척한 다음 absolute alcohol로 탈 수과정을 거친 후 형광 현미경(Carl Zeiss)을 이용하여 400 배의 배율로 핵의 형태변화를 관찰하였다.
5. DNA flow cytometer에 의한 세포 주기의 분석
FS, FBA 및 FYA를 농도별로 처리한 배지에서 배양된 U937 세포를 모아서 PBS로 충분히 수세 후 CycleTEST Plus DNA Reagent Kit (Becton Dickinson, San Jose, CA, USA) 를 사용하여 고정 및 propidium iodide (PI) 염색을 하여 4oC, 암조건에서 10분 이상 반응을 시켰다. 염색된 세포 를 35-mm mesh를 이용하여 단일세포로 분리한 후 FACSCalibur (Becton Dickinson)를 이용하여 형광반응에 따 른 Cellular DNA content 및 histogram을 CellQuest software 및 ModiFit LT (Becton Dickinson) 프로그램을 이용하여 분 석하였다.
6. DNA fragmentation 분석
특정 세포에 apoptosis가 유발되었을 때 나타나는 DNA 단편화의 관찰을 위하여 동일한 조건으로 배양된 세포 를 모아 lysis buffer [5 mM Tris-HCl (pH 7.5), 5 mM EDTA, 0.5% Triton X-100]를 4oC에서 30분간 처리한 다음, 그 상 층액에 proteinase K (Sigma)를 0.5 mg/ml의 농도로 처리한
다음 50oC에서 3시간 동안 반응시켰다. 그 후 phenol:
chloroform:isoamyl alcohol 혼합용액 (25:24:1, Sigma)을 첨가 하여 30분간 inverting 한 뒤 원심분리하여 따로 분리한 상층액에 적정량의 isopropanal (Sigma)과 5 M NaCl를 첨 가한 다음 4oC에서 overnight시켰다. 이후 원심분리하여 분리된 DNA pellet을 70% ethanol로 충분히 세척하고 RNase A가 적당량 들어있는 DEPC water에 녹여 gel loading dye (Bioneer, Daejeon, Korea)를 혼합한 후 1.5%
agarose gel에 1시간 정도 50 V로 전기영동하였다. 이후 ethidium bromide (EtBr, Sigma)로 염색하여 ultra violet (UV) 하에서 관찰하였다.
7. Reverse transcription-polymerase chain re- action (RT-PCR) 분석
동일한 조건에서 배양한 U937 세포를 PBS로 세척하 고, TRIzol reagent (Invitrogen Co., Carlsbad, CA, USA)를 4oC 에서 1시간 동안 처리하여 total RNA를 분리하였다. 분리 된 RNA를 정량한 후, 각각의 primer, DEPC water 그리고 ONE-STEP RT-PCR PreMix Kit (Intron, Korea)를 넣고 Mastercycler gradient (Eppendorf, Hamburg, Germany)를 이 용하여 증폭하였다. 각 PCR 산물들의 양적 차이를 확인 하기 위하여 1 X TAE buffer로 1% agarose gel을 만들고 well 당 각각의 primer에 해당하는 PCR 산물에 DNA gel loading solution을 섞어서 loading 한 후 50 V에서 전기영동을 행하 였다. 전기영동으로 DNA 분리가 끝난 gel을 EtBr을 이용 하여 염색한 후 UV하에서 확인하고 Picture works' photo enhancer를 이용하여 사진 촬영을 하였다. 이 때 housekeeping 유전자인 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) 유전자를 internal control로 사용하였다.
8. 단백질의 분리, 전기영동 및 Western blot analysis
FS, FBA 및 FYA를 처리한 배지에서 자란 세포들을 모 아 PBS로 씻어내고, 적당량의 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenylmethylsulfonyfluoride (PMSF), 5 mM dithiothreitol (DTT)]를 첨가하여 4oC에서 1 시간 반응시킨 후, 14,000 rpm으로 30분간 원심분리하여 그 상층액을 취하였다. 상 층액의 단백질 농도를 Bio-Rad 단백질 정량시약(Bio-Rad, Hercules, CA, USA)을 사용하여 정량한 다음 동량의 Laemmli sample buffer (Bio-Rad)를 섞어서 sample을 만들었 다. 이렇게 만든 동량의 단백질을 sodium dodecyl sulphate (SDS)-polyacrylamide gel을 이용하여 전기영동으로 분리하 였다. 분리된 단백질을 함유한 acrylamide gel을 nitrocellul- ose membrane (Schleicher and Schuell, Keene, NH, USA)으로
Fig. 1. Effects of ethanol extracts of fermented beans on the cell viability in U937 human leukemia cells. The cells were incubated with indicated concentrations of fermented soybean (FS), black agabean (FBA) and yellow agabean (FYA) ethanol extracts for 48 h. The cell viabilities were measured by MTT assay. The data shown are means±SD of three independent experiments.
전이시킨 후, 5% skim milk를 함유한 PBS-T (0.1% Tween 20 in PBS)를 처리하여 비특이적인 단백질에 대한 block- ing을 실시하였다. 준비된 membrane에 1차 항체를 처리 하여 4oC에서 overnight 시킨 후 PBS-T로 세척하고 처리 된 1차 항체에 맞는 2차 항체를 사용하여 상온에서 1시 간 정도 반응 시켰다. 다시 PBS-T로 세척하고 enhanced cheiluminoscence (ECL) 용액(Amersham Life Science Corp., Arlignton Heights, IL, USA)을 적용시킨 다음 암실에서 X-ray film에 감광시켜 특정단백질의 양을 분석하였다.
본 실험에 사용된 항체들은 Santa Cruz Biotechnology Inc.
에서 구입하였으며, immunoblotting을 위해 2차 항체로 사용된 peroxidase-labeled donkey anti-rabbit immunoglobulin 및 peroxidase-labeled sheep anti-mouse immunoglobulin은 Amersham Life Science Corp.에서 구입하였다.
9. In vitro caspase activity 측정
Caspase의 활성 정도가 FS, FBA 및 FYA 처리에 의하여 어떠한 변화를 유발하는지 알아보기 위하여 선행조건에 의해 처리된 배지에서 배양된 세포를 모은 뒤 상기와 동 일한 조건으로 단백질을 추출하고 정량하였다. 이렇게 준비된 단백질 150μg에 fluorogenic peptide 기질 100μM 이 함유된 extraction buffer [40 mM HEPES (pH 7.4), 20%
glycerol (v/v), 1 mM EDTA, 0.2% NP-40 and 10 mM DL-DTT] 50μl을 혼합하였으며, microtiter plate에 다시 extraction buffer에 희석하여 각 sample 당 총 volume이 100 μl가 되게 하였다. 실험에 사용된 기질은 caspase-3의 경 우에는 Asp-Glu-Val-Asp (DEVD)-p-nitroaniline (pNA)이었 고, caspase-8의 경우에는 Ile-Glu-Thr-Asp (IETD)-pNA이었 으며, caspase-9은 Leu-Glu-His-Asp (LEHD)-pNA였다. 준비 된 plate를 37oC에서 빛을 차단한 다음 3시간 동안 incubation 시킨 후 ELISA reader를 이용하여 405 nm의 흡 광도를 이용하여 반응의 정도를 측정하였다. Caspases 활 성 측정을 위한 colorimetric assay kits는 R&D Systems (Minneapolis, MN, USA)에서 구입하였다.
결 과
1. FS, FBA 및 FYA 처리에 의한 U937 세포의 증식 억제 및 형태 변형
FS, FBA 및 FYA가 인체 혈구암세포인 U937의 증식에 미치는 영향을 알아보기 위하여 FS, FBA 및 FYA를 적정 농도별로 처리하여 48 시간 배양한 다음 MTT assay를 실 시한 결과는 Fig. 1에서 나타낸 바와 같다. 결과에서 알 수 있듯이 FS, FBA 및 FYA를 48시간 처리한 결과 농도
의존적으로 인체 혈구암 세포 증식의 억제 효과를 가지 는 것을 확인할 수 있었다. 특히 이러한 효과는 FS보다 FBA와 FYA에서 뛰어났는데, FBA, FYA를 3 mg/ml로 처 리하였을 때 각각 약 70%, 60%의 억제능을 보였다. FS, FBA 및 FYA 처리에 의한 U937 세포의 증식억제 현상과 연관된 세포의 형태에는 어떠한 영향을 미치는지를 알 아보기 위해 동일조건에서 배양된 세포를 도립현미경을 통하여 관찰하였다. Fig. 2A에서 알 수 있듯이 FS, FBA 및 FYA를 처리하지 않은 세포와 비교하였을 때 FS, FBA 및 FYA를 처리한 세포의 밀도는 농도 의존적으로 감소 하였고, 세포의 모양 또한 불규칙적으로 변화된 것을 확 인할 수 있었다.
2. FS, FBA 및 FYA 처리에 의한 U937 세포의 apop- tosis 유발
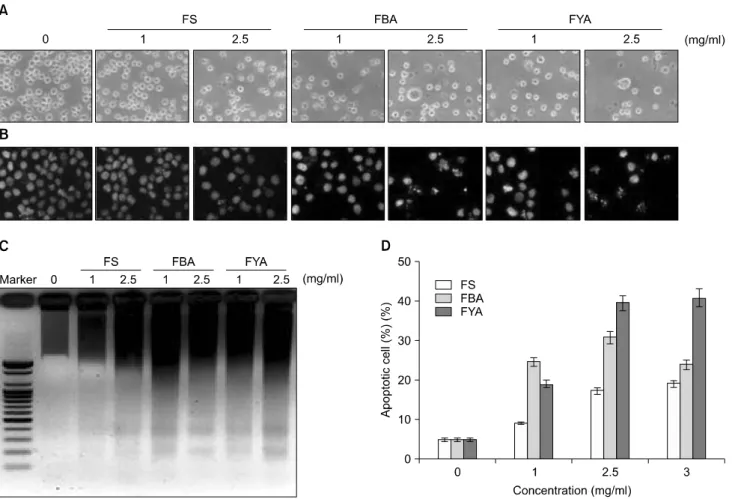
FS, FBA 및 FYA 처리에 의한 U937 세포의 증식억제가 apoptosis의 유발과 관계가 있을 것으로 예상 되어 핵산에 특이적으로 결합하는 DAPI 염색을 실시하고 형광현미 경을 통하여 핵의 형태 변화를 관찰하였다. 그 결과 Fig.
2B에서와 같이 FS, FBA 및 FYA를 처리하지 않은 배지에 서 배양된 세포의 경우에는 핵의 전체가 고루 염색되는 양상을 보였으나 FS, FBA 및 FYA를 처리하였을 처리 농 도가 높아질수록 전형적으로 apoptosis가 일어난 세포에 서 관찰되는 염색질의 응축에 의한 apoptotic body의 형성 이 증가한 것을 확인하였다. 특히 이러한 현상들은 FS에 비하여 FBA와 FYA의 고농도에서 더욱 확연히 나타났다.
Fig. 2. Apoptosis induction of U937 cells by ethanol extracts of fermented beans treatment. (A) U937 cells were seeded at 1×105/ml in a 6-well plate and incubated for 24 h. The cells were treated with variable concentrations of FS, FBA and FYA for 48 h. Cell morphology was visualized by an inverted microscopy (Original magnification, ×200). (B) Cells were treated with FS, FBA and FYA for 48 h and then stained with DAPI solution. After 15 min incubation at room temperature, the cells were washed with PBS and nuclear morphology was photographed with a fluorescent microscope using blue filter (Original magnification, ×400). (C) Cells grown under the same condition as A were collected and the genomic DNA was extracted. For the analysis of DNA fragmentation, the genomic DNA was analyzed on a 1.5% agarose gel containing EtBr. (D) To quantify the degree of apoptosis induced by FS, FBA and FYA, the cells grown under the same conditions as (A) were collected and stained with PI for flow cytometry analysis. The percentages of cells with hypodiploid DNA (sub-G1 phase) contents represent the fractions undergoing apoptotic DNA degradation. Data are expressed as mean±SD of three independent experiments.
또한 염색질 응축과 더불어 apoptosis 유발의 직접적인 증 거에 해당하는 DNA의 단편화 여부를 확인한 결과, Fig.
2C에서 볼 수 있듯이 FS, FBA 및 FYA의 처리에 의해 DNA가 단편화되었음을 알 수 있었다. 이러한 apoptosis 유발 정도를 정량적으로 분석하기 위하여 DNA flow cytometry를 이용하여 세포주기 중 apoptosis가 일어난 세 포군에 해당하는 sub-G1기의 빈도를 조사한 결과, FS, FBA 및 FYA 처리 농도의 증가에 따라 sub-G1기에 해당 하는 세포의 빈도가 증가하는 것을 확인하였다. 이 결과 는 특히 FYA의 처리군에서의 증가율이 FS와 FBA보다 높 았으며, 최고 처리농도인 3 mg/ml에서는 약 40%로 나타 났다(Fig. 2D). 이상의 결과를 통하여 FS, FBA 및 FYA에
의해 일어나는 인체혈구암세포의 증식억제 및 형태적 변화는 apoptosis와 밀접한 연관성이 있음을 확인하였다.
3. Bcl-2 및 IAP family의 발현에 미치는 FS, FBA 및 FYA의 영향
다음으로 FS, FBA 및 FYA의 처리에 의해 유발되는 apoptosis에 어떠한 유전자가 관여하는지 조사하기 위하 여 먼저 Bcl-2 family에 속하는 인자들의 발현 변화를 RT-PCR과 Western blotting을 통하여 조사하였다. 그 결과 전사수준에서는 apoptosis 유발인자와 억제 인자 모두 큰 변화가 없었지만, 번역수준에서는 FS, FBA 및 FYA를 처 리하였을 때 apoptosis 억제 인자인 Bcl-2 및 Bcl-xL의 발현
Fig. 3. Effects of FS, FBA and FYA treatment on the Bcl-2 family members expression in U937 cells. (A) After 48 h incubation with FS, FBA and FYA, total RNAs were isolated and reverse-transcribed. The resulting cDNAs were subjected to PCR with the indicated primers, and the reaction products were subjected to electrophoresis in 1.0% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells grown under the same conditions as (A) were lysed and then cellular proteins were separated by 10∼
13% SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
Fig. 4. Effects of FS, FBA and FYA treatment on the IAP family members expression in U937 cells. (A) After 48 h incubation with FS, FBA and FYA, total RNAs were isolated and reverse-transcribed. The resulting cDNAs were subjected to PCR with the indicated primers, and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells grown under the same conditions as (A) were lysed and then cellular proteins were separated by 10%
SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
이 감소하였음을 확인하였고, 이러한 발현 감소는 FS 보 다 FBA 및 FYA 처리시, 특히 FYA를 처리하였을 때 현저 하게 감소하는 것으로 나타났다. 또한 apoptosis 유발 인 자인 Bax 및 Bad의 경우 FS, FBA 및 FYA의 처리 농도가 높아질수록 발현 양이 훨씬 증가하는 것으로 나타났고, 이 결과 역시 고농도의 FYA 처리군에서 더욱더 증가된 발현을 관찰할 수 있었다(Fig. 3). 한편 IAP family에 속하 는 인자들은 caspase와의 직ㆍ간접적인 결합을 통하여 그 들의 활성을 억제하여 apoptosis 유발을 억제하는 것으로 알려져 있는데, 본 실험의 조건에서 몇 가지 이들 family 의 발현에는 큰 변화가 관찰되지 않았다(Fig. 4).
4. Death receptor family의 발현에 미치는 FS, FBA 및 FYA의 영향
다음은 apoptosis의 extrinsic pathway에 관여하는 유전자 들의 발현에 미치는 FS, FBA 및 FYA의 효과를 RT-PCR 및 Western blotting으로 확인하였다. Fig. 5A에 나타낸 바 와 같이 FS, FBA 및 FYA 처리군에서 Fas, Fas ligand (FasL) 의 mRNA 발현이 대조군에 비하여 증가되었으며, 단백 질 수준에서 Fas, FasL, Fas-associated protein with death domain (FADD) 및 TNFRSF1A-associated via death domain (TRADD)의 발현이 증가되었음을 확인하였는데(Fig. 5B), 이 결과에서도 FS 처리군보다 FBA와 FYA 처리군에서 발
Fig. 6. Effects of FS, FBA and FYA treatment on the express- ion and activity of caspases in U937 cells. (A) After 48 h incubation with FS, FBA and FYA, the cells were lysed, and cellular proteins were separated by 8∼13% SDS-polyacryla- mide gel and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies.
Proteins were visualized using an ECL detection system. Actin was used as an internal control. (B) The cells grown under same condition as (A) were collected and then lysed. Aliquots (100μg proteins) were assayed for in vitro caspase-3, caspase-8 and caspase-9 activity using DEVD-pNA, IETD- pNA, and LEHD-pNA, respectively, as substrates. The released fluorescent products were measured. The data represents the average±SD of three independent experiments.
Fig. 5. Effects of FS, FBA and FYA treatment on the levels of death-receptor related gene products in U937 cells. (A) After 48 h incubation with FS, FBA and FYA, total RNAs were isolated and reverse-transcribed. The resulting cDNAs were subjected to PCR with the indicated primers, and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells grown under the same conditions as (A) were lysed and then cellular proteins were separated by 10∼13% SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
현 변화의 정도가 높게 나타났다. 이상의 결과에서 FS, FBA 및 FYA 처리에 의한 apoptosis 유발은 death receptor pathway에 관여하는 death receptor 관련 유전자의 발현 증 가와 mitochondrial pathway에 관여하는 Bcl-2 family의 pro-apoptosis 인자의 발현 증가 및 anti-apoptosis 인자의 발
현 억제 등의 복합적인 경로를 통하여 이루어지는 것을 확인하였다.
5. Caspases의 발현 및 활성에 미치는 FS, FBA 및 FYA의 영향
다음은 apoptosis 유발에서 가장 핵심적인 역할을 하는 caspases의 활성에 미치는 FS, FBA 및 FYA의 영향을 조사 하였다. 먼저 Western blotting을 통하여 caspases 단백질 수준에서의 발현 변화를 관찰한 결과, Fig. 6A에서 볼 수 있듯이 FS, FBA 및 FYA를 처리하였을 때 조사된 3가지 caspases (-3, -8 및 -9) 모두 비활성형의 발현이 감소하였 으며, caspase-8 및 -9의 경우 활성형의 증가도 확인할 수 있었다. 또한 apoptosis가 유발되었음을 보여주는 지표 단 백질로서 활성화된 caspase-3의 표적 단백질인 PARP 또 한 단편화가 유도되었음을 알 수 있었다.7) 또한 FS, FBA 및 FYA에 의한 caspases의 활성 여부의 정량적 확인을 위 하여 in vitro 분석을 실시한 결과, Fig. 6B에 나타낸 것처 럼 FS, FBA 및 FYA 처리군에서 모두 처리 농도 의존적으 로 활성이 증가되었음을 알 수 있었다.
고 찰
본 연구에서는 인체 혈구암유래 U937 세포의 성장에 미치는 발효콩 추출물의 영향과 발효콩 추출물 처리에 의한 암세포 성장 억제와 연관된 apoptosis 유발여부 및 관련 유전자들의 발현변화를 조사하였다. 먼저 FS, FBA 및 FYA가 U937 세포의 증식에 미치는 영향을 조사해본 결과 FS, FBA 및 FYA를 처리하였을 경우 처리 농도 의존 적으로 세포의 증식을 억제되었음을 확인할 수 있었으 며, 이때 시료의 농도가 증가함에 따라 세포의 밀도도 감소되며 다양한 세포의 형태적 변화가 동반되었음을 알 수 있었다(Fig. 1, 2A). 이러한 FS, FBA 및 FYA의 처리 에 의한 U937 세포의 증식억제가 apoptosis 유발과 연관 되어 있는지를 확인하기 위해 DAPI staining을 실시하였 으며, FS, FBA 및 FYA를 처리하였을 때 처리 농도 의존 적으로 전형적인 apoptosis가 일어난 세포에서 관찰되는 염색질 응축에 의한 apoptotic body의 형성이 증가되었고, 세포의 밀도 또한 감소하는 것을 확인하였다(Fig. 2B). 또 한 염색질 응축과 더불어 apoptosis 유발의 또 다른 직접 적인 증거에 해당하는 DNA의 단편화 여부를 조사한 결 과 FS, FBA 및 FYA의 고농도 처리군에서 모두 DNA의 단편화가 일어났음이 관찰되었으며(Fig. 2C), 이러한 apoptosis 유발 정도를 정량적으로 알아보기 위해 DNA flow cytometry를 이용하여 세포 주기 중 apoptosis가 일어 난 세포군에 해당하는 sub-G1기의 빈도를 조사한 결과, FS, FBA 및 FYA 처리 농도의 증가에 따라 sub-G1기에 해
당하는 세포의 빈도가 증가하는 것을 확인하였다(Fig.
2D).
Apoptosis의 유발 경로는 크게 2가지로 구분할 수 있는 데, 이 중 intrinsic (mitochondrial-mediated) pathway의 경우 에는 mitochondria 외막에 존재하는 Bcl-2 family가 중요한 역할을 하게 된다. Bcl-2 family에 속하는 단백질은 apoptosis를 유발하는 인자인 Bax, Bak, Bad 및 Bid 등이 있으며 apoptosis를 억제하는 인자인 Bcl-2, Bcl-xL 등이 존 재하며 이들은 dimer의 형태로 결합하여 균형을 이루고 있는 것으로 알려져 있다. 하지만 세포적 스트레스나 세 포 독성에 의해 mitochondria의 막 전위가 변화되면, 이 균형은 깨어지고, mitochondria에서 cytochrome c와 같은 apoptosis 유발 인자들이 세포질로 방출된다. 방출되어진 cytochrome c는 apoptotic protease activating factor (Apaf)-1과 caspase-9에 결합함으로써 apoptosome을 형성하여 caspase-9 를 활성화시키며, 결과적으로 caspase-3을 활성화시킴으 로써 apoptosis를 유발한다.1,2) Apoptosis의 또 다른 경로로 알려진 extrinsic (death receptor-mediated) pathway의 경우에 는 TNF family의 death receptor들의 활성에 의해 야기된 다. Apo-2 ligand로 알려진 TRAIL은 두 가지 death domain- containing agonistic receptor인 death receptor (DR) 4 또는 DR5와 결합함으로써 여러 종류의 암세포에서 apoptosis 를 유발하는 것으로 알려져 있다. 한편 tumor necrosis factor (TNF) receptor superfamily에 속하는 Fas에 FasL가 결 합하거나 TNFR-1에 TNF-α가 결합하면 FADD 또는 TRADD 및 procaspase-8이 DISC를 형성하게 되어 caspase-8 의 활성화에 의한 caspases cascade에 의하여 apoaptosis를 유발하게 된다.3,4) 본 연구에서 나타난 FS, FBA 및 FYA 처리에 따른 apoptosis가 어떠한 경로를 통하여 일어나는 지 확인하기 위해 먼저 Bcl-2 및 IAP family 인자들의 발 현에 FS, FBA 및 FYA의 영향을 알아본 결과 전사수준에 서는 apoptosis 유발인자와 억제 인자 모두 큰 변화가 없 었지만, 번역수준에서는 apoptosis 억제 인자인 Bcl-2 및 Bcl-xL의 발현 감소와 apoptosis 유발 인자인 Bax 및 Bad의 발현 증가를 관찰할 수 있었다(Fig. 3). 하지만 IAP family 는 FS, FBA 및 FYA의 처리에도 큰 변화가 관찰되지 않았 다(Fig. 4). 다음으로 FS, FBA 및 FYA의 처리에 의한 death receptor에 속하는 유전자들의 변화를 RT-PCR 및 Western blotting으로 확인한 결과, 먼저 전사 수준에서 FS, FBA 및 FYA의 처리에 따라 Fas 및 FasL의 발현이 대조군에 비하여 증가된 것을 확인하였고 Fas, FasL, FADD, TRADD의 발현양도 증가하는 것을 관찰하였다(Fig. 5).
한편 cysteine에 의존적으로 aspartate에 직접적인 단백질 분해효소 그룹인 caspases는 apoptosis에 중요한 역할을 하
는 것으로 알려져 있다. 평상시 caspases는 비활성 전구체 로 합성되어져 있지만, 여러 활성화된 신호에 의해서 apoptosis가 시작되면 활성화되기 시작한다. 활성이 시작 될 때 caspase는 많은 표적단백질의 분해를 유발하여 세 포의 분해를 주재함으로써 apoptosis 과정에서 큰 역할을 한다. 이러한 caspase에는 initiator caspases (caspase-8, -9, -10 및 -12)와 effector caspases (caspase-3, -6 및 -7)로 나누어 지는데 외부의 다양한 신호전달에 의하여 initiator caspase 가 활성화가 되어 effector caspase를 활성화 시켜 apoptosis 를 유발하므로, 많은 선행연구에서 caspases의 활성화가 apoptosis의 유발에 대한 증거가 될 수 있다고 보고되어지 고 있다.5,6) 따라서 FS, FBA 및 FYA가 caspasee의 발현에 어떠한 영향을 미치는지 알아본 결과 FS, FBA 및 FYA를 처리에 의하여 caspase-3, -8 및 -9의 활성이 농도 의존적 으로 증가하는 것을 Western blot analysis와 in vitro caspase activity assay를 통하여 확인할 수 있었으며, PARP 또한 단편화가 유발되었음을 확인하였다. 이러한 결과들은 FS, FBA 및 FYA의 처리에 의한 apoptosis 유발에 두 apoptosis 유발 경로가 모두 부분적으로 관여하고 있음을 보여주는 것이다.
결 론
본 연구에서는 isoflavone이 다량으로 함유되었을 것으 로 추정되는 콩의 발효 추출물에 의한 U937 인체 혈구암 세포의 증식억제가 apoptosis 유발과 밀접한 관련이 있음 을 확인하였다. 본 연구의 결과에 의하면 FS, FBA 및 FYA의 처리에 따라 U937 세포의 증식이 처리 농도의존 적으로 억제되었으며, 이러한 세포 증식억제가 apoptosis 유발에 의한 것임을 다양한 방법으로 제시하였다. 또한 RT-PCR과 Western blot analysis를 통해 FS, FBA 및 FYA에 의한 apoptosis는 세포 표면에 존재하는 Fas에 FasL가 결합 하여 일어나는 death receptor pathway 또는 pro-apoptosis 인 자인 Bax와 Bad의 증가와 anti-apoptosis 인자인 Bcl-2와 Bcl-xL의 감소에 의해 일어나는 mitochondrial pathway를 통하여 유발될 것이라고 생각되어진다. 이러한 콩 발효 산물의 항암 효과는 대두 추출물(FS)에서 보다 아가콩 추 출물(FBA 및 FYA)에서 더욱 효과가 높게 나타났으며, 이 는 향후 아가콩 추출물의 암예방 및 항암식품으로의 개 발 가능성이 매우 높음을 제시하여 주는 결과이다.
감사의 글
본 연구는 농림수산식품부 농림기술개발사업의 지원
(Medi-Farm 산업화 연구사업단, 610003-03-1-SU000)에 의 해 이루어진 것임.
참 고 문 헌
1) Khan N, Adhami VM, Mukhtar H. Apoptosis by dietary agents for prevention and treatment of cancer. Biochem Pharmacol 76, 1333-1339, 2008.
2) Burz C, Berindan-Neagoe I, Balacescu O, Irimie A. Apoptosis in cancer: key molecular signaling pathways and therapy targets. Acta Oncol 48, 811-821, 2009.
3) Brenner D, Mak TW. Mitochondrial cell death effectors. Curr Opin Cell Biol 21, 871-877, 2009.
4) Jeong SY, Seol AW. The role of mitochondria in apoptosis.
BMB Rep 41, 11-22, 2008.
5) Earnshaw WC, Martins LM, Kaufmann SH. Mammalian caspases: structure, activation, substrates, and functions during apoptosis. Annu Rev Biochem 6, 383-424, 1996.
6) Stennicke HR, Salvesen GS. Properties of the caspases. Biochim Biophys Acta 1387, 17-31, 1998.
7) Kaufmann SH, Desnoyers S, Ottaviano Y, Davidson NE, Poirier GG. Specific proteolytic cleavage of poly(ADP-ribose polymerase: an early marker of chemotherapy-induced apop- tosis. Cancer Res 53, 3976-3985, 1993.
8) Inomata N, Osuna H, Kawano K, Yamaguchi J, Yanagimachi M, Matsukura S, Ikezawa Z. Late-onset anaphylaxis after ingestion of Bacillus subtilis-fermented soybeans (Natto):
clinical review of 7 patients. Allergol Int 56, 257-261, 2007.
9) O'Toole DK. Characteristics and use of okara, the soybean residue from soy milk production-a review. J Agric Food Chem 47, 363-371, 1999.
10) Barnes S. The biochemistry, chemistry and physiology of the isoflavones in soybeans and their food products. Lymphat Res Biol 8, 89-98, 2010.
11) Pasqualini JR. Estrogen sulfotransferases in breast and endometrial cancers. Ann N Y Acad Sci 1155, 88-98, 2009.
12) Perabo FG, Von Löw EC, Ellinger J, von Rücker A, Müller SC, Bastian PJ. Soy isoflavone genistein in prevention and treatment of prostate cancer. Prostate Cancer Prostatic Dis 11, 6-12, 2008.
13) Choi UK, Kim MH, Lee NH, Jeong YS, Kwon OJ, Kim YC, Hwang YH. The characteristics of cheonggukjang, a ferment- ed soybean product, by the degree of germination of raw soybeans. Food Sci Biotechnol 16, 734-739, 2007.
14) Phommalth S, Jeong YS, Kim YH, Dhakal KH, Hwang YH.
Effects of light treatment on isoflavone content of germinated soybean seeds. J Agric Food Chem 21, 10123-10128, 2008.
15) Lee HH, Park C, Kim MJ, Seo MJ, Choi SH, Jeong YK, Choi YH. Inhibition of cyclooxygenase-2 activity and prostaglandin E2 production through down-regulation of NF-κB activity by the extracts of fermented beans. J Life Sci 20, 388-395, 2010.