IntroductIon
Oryzias, a ricefish genus which lives in ponds, stagnant streams and irrigated fields with no fast flowing, has cur- rently 33 species in the world(Parenti, 2008; Mokodon- gan et al., 2014). Among them, it is certain that the hab- itat of O. sinensis could be on chemically and physically adverse conditions caused by water pollution and a dras- tic change such as water levels and currents in the rice fields where used to get dry periodically. Recently, it has been revealed that the micro-structure of its egg envelope has relevance to environmental changes in its habitat(Kim and Park, 2014). Likewise, even the olfactory organ of te- leost can be a good indicator to acknowledge its environ- mental adaptation. It allows fish to perform the olfaction as a crucial sense for survival, such as a feeding, repro- duction, migration and predator avoidance(Hara, 1986).
Therefore, the olfactory organ in teleost fishes show any
difference based on the following characters: 1) the gross structure, 2) the arrangement and number of lamellae, 3) the distribution of the olfactory epithelium, 4) cell types forming the epithelium(Zeiske et al., 1992; Kasumyan, 2004). Even if in a sort of genus Oryzias prominent and large eyes play major roles in the over habits, the olfacto- ry organ is not only presumed to be important throughout the lifecycle but also may be affected by environmental factors unknown. For this assumption in teleost fishes, previous anatomical and histochemical studies have been only conducted through visual observation as well as histochemical studies under diverse microscopes(Ghosh and Chakrabarti, 2009, 2011; Sarkar and De, 2011; Kim et al., 2014, 2016). However, some of the olfactory or- gan in O. latipes has only been reported by Yamamoto (1982), and information on the anatomy and histochem- istry of genus Oryzias is required further for any relation between the environment and adaptation. Thus, the aim of this study is to describe the anatomy and histology of the olfactory organ in O. sinensis and draw comparison analysis with relationship to its habitat.
—223 — http://www.fishkorea.or.kr
* Corresponding author: Jong Young Park Tel: 82-63-270-3344, Fax: 82-63-270-3362, E-mail: [email protected]
ISSn: 1225-8598(Print), 2288-3371(online)
accepted: December 23, 2016
An Anatomical and Histochemical Study of the Olfactory Organ in Rice-fish Oryzias sinensis (Pisces: Adrianichthyidae) in South Korea
By Hyun Tae Kim, Yong Joo Lee
1and Jong Young Park*
Department of Biological Science and Institute for Biodiversity Research, College of Natural Sciences, Chonbuk National University, Jeonju 54896, Republic of Korea
1Jeonju National University of Education, Jeonju 55101, Republic of Korea
ABSTRACT the anatomy and histology of the olfactory organ in Oryzias sinensis was researched using a stereo microscope and light microscope. In the gross structure, the paired olfactory organs on the dorsal part of the head consist of two nostrils(a circular anterior nostril and a slit posterior nostril in a distance), a single olfactory chamber and a single accessory nasal sac. In the histological study, the epithelium of the olfactory chamber is classified into both sensory and non-sensory regions. the sensory epithelium consists of olfactory receptor neurons, supporting cells, basal cells and vesicles, and is islet in distributional pattern. the non-sensory epithelium is composed of stratified epithelial cells and two types of mucous cells(acidic and neutral cells). the epithelium of the accessory nasal sac has swollen stratified epithelial cells, mucous cells with a rich glycoprotein. Such an olfactory anatomy and histology of O. sinensis may reflect its habitat surrounding stagnant and polluted water.
Key words: Two nostrils, single olfactory chamber, single accessory nasal sac, mucous cell
MaterIalS and MethodS
O. sinensis ranging from 21.0 to 27.9mm in standard length(Fig. 1B) were collected between August and Octo- ber 2016 in farm waterway of Songcheon2-dong, Deok- jin-gu, Jeonju-si, Jeollanbuk-do, South Korea, 35°53′
28″N, 127°06′ 56″E(Fig. 1A). Twenty specimens caught by a kick net(2×2mm) were fixed in 10% neutral buffered formalin solution. Ten were used for anatomic observation under stereo microscope(SM; Stemi DV4, Carl Zeiss, Germany) and photographed with a digital camera(TG- 3, Olympus, Japan). The remainder(5 each for sex) was used to identify histological and cytological structures under light microscope(LM; LE REL 4.4, Carl Zeiss, Germany). For a LM observation, fish’s head with the olfactory organ was resected from specimens. It was de- hydrated properly through an ascending series of ethanol, was cleaned with xylene, and then was embedded in par- affin wax(Paraplast, Oxford) for 24 hours. Sections with the olfactory tissue were cut at five micrometer, deparaf- finized, tested such as Harris’s hematoxylin-eosin(H-E) and Masson’s trichrome(Gurr, 1956), and Alcian blue-
Periodic acid Schiff(AB-PAS, pH 2.5) stainings(Mowry, 1956).
reSultS
Gross structure
Externally, the paired olfactory organs are situated dor- sally on the each side of the snout in the head. They have each two nostrils for the ventilation of water inlet and outlet, being formed a circular anterior nostril and a slit posterior nostril in a distance(Fig. 2A). An internal struc- ture has a single olfactory chamber without a folded lam- ellae and an accessory nasal sac to be expanded ventrally (Fig. 2B, C). The olfactory chamber opens externally by two nostrils(Fig. 2C).
histology
Within the olfactory chamber of triangle shape in a cross section, the epithelial region is mainly subdivided into both a sensory epithelium on ventrally and laterally
Fig. 1. The habitat(A) and photograph(B, male) of Oryzias sinensis. The bar indicates 1cm.
A B
Fig. 2. The gross olfactory structure of Oryzias sinensis. A, an external structure showing an anterior nostril(AN) and a posterior nostril(PN); B, an internal structure with an olfactory chamber(OC) and a single accessory nasal sac(NS); C, a diagram of the olfactory structure.
A B C
and a non-sensory epithelium on dorsally(Fig. 3A).
The sensory epithelium of a pseudo-stratified layer is made up of three types of cells: olfactory receptor neu- rons(ORN), supporting cells, basal cells(Fig. 3B), and is an islet in the distributional pattern, being separated by the non-sensory epithelium(Fig. 3A and D). As the ORNs are bipolar neurons extending from the bottom to the api- cal surface of the epithelium, their length by the measure- ment of the epithelial thickness is mean 126.7μm(103.3
~169.0). The ORNs have nuclei stained strongly with hematoxylin, and their axon and dendritic processes are stained faintly with eosin. The supporting cells are cylin- drical to have more faintly stained-nuclei with hematox-
ylin than the neurons. The position of its nuclei is also in the upper part than those of the neurons. The basal cells with elliptical nuclei are situated at the bottom of the ep- ithelium. A number of vesicles are observed in the sen- sory epithelium. They, with the range of 16.0~25.5μm in diameter, are circular and do not react to H-E staining (Fig. 3B).
The non-sensory epithelium of a stratified cuboidal lay- er is composed of three types of cells: stratified epithelial cells, mucous cells and unidentified epithelial cells(Fig.
3C-F). The stratified epithelial cells have nuclei stained strongly with hematoxylin, and are swollen in the cyto- plasm(Fig. 3C). Mucous cells, which are positive to AB-
Fig. 3. A histology and histochemistry of the olfactory organ of Oryzias sinensis with hematoxylin-eosin(A to C), an alcian blue-periodic acid Schiff(pH 2.5)(D, E) and Masson’s trichrome(F) stainings. A, the epithelium of the olfactory chamber is subdivided into a sensory(SE) and a non-sensory epithelium(NSE); B, the SE comprising olfactory receptor neurons(ORN), supporting cells(SC), basal cells(BC), vesicles(V) and capillaries(CA) just under the bottom of the epithelium; C, the NSE built of stratified epithelial cells(SEC) and mucous cells(MC) at the apical part; D, acidic mucous cells with a blue color and unidentified epithelial cells(UC) in the NSE; E, acidic mucous cells with a blue color and neu- tral mucous cells with a murex color on the NSE; F, mucous cells and unidentified cells on the NSE. Arrows indicate NSEs. Arrowheads indicate mucous cells. Bars indicate 200μm in A and 50μm in B to F, respectively.
A
B
C
D
E
F
PAS staining but negative to H-E and Masson’s trichrome staining, are classified into two types of cells(acidic and neutral cells). They also are holocrine gland that both cytoplasm and secretion are released(Fig. 3C-F). Acidic mucous cells of a blue color were alcian blue positive, whereas neutral mucous cells of a red-purple color were basic fuchsin positive(Fig. 3E). An unidentified cylindri- cal cells are discovered only in the non-sensory epitheli- um, and are marked with red color by basic fuchsin and biebrich scarlet stainings(Fig. 3D, F).
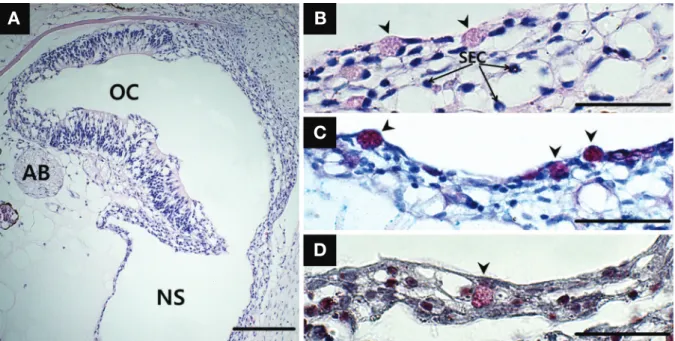
In the accessory nasal sac expanding ventrally from the olfactory chamber(Figs. 2B and 4A), the stratified cuboi- dal epithelium on its floor comprises two types of cells:
swollen stratified epithelial cells and mucous cells. It also is built of the 4~5 cells layer(Fig. 4B). The cytoplasm of stratified epithelial cells, one of major constituent units, is decreasing in size while getting closer to the surface (Fig. 4B). Mucous cells positive to H-E staining show a weak blush color and a strong red-purple with AB-PAS (pH 2.5) staining and a bright red color with Masson’s trichrome staining(Fig. 4B-D).
dIScuSSIon
In both stereo and light microscopic levels, the struc- tural and histological results on the olfactory organ of
Oryzias sinensis, are as follows: two nostrils in a dis- tance, a single olfactory chamber without lamellae, a single accessory nasal sac, an islet in the distributional pattern of the sensory epithelium and various types of mucous cells.
Compared with other teleost fishes, the gross olfactory structure of O. sinensis corresponds to the general pattern such as the dorsal position of two nostrils in the head, the formation of an olfactory chamber and an accessory nasal sac(Zeiske et al., 1992; Kasumyan, 2004). However, based on the ecological habits by given species, the detailed structure on the olfactory organ in bony fishes differs sig- nificantly across species. Despite of fishes living a stag- nant freshwater, an inactive swimmer O. sinensis exhibits small two nostrils separated by a nasal bridge, whereas an active swimmer Zebra danio has large two nostrils close to each other(Hansen and Zeiske, 1988). Meanwhile, each paired nostril of many fishes, which are active or live in flowing water, is situated close to each other extremely due to demarcation of a thin nasal flap projecting out from the skin(Ojha and Kapoor, 1973; Cox, 2008; Kumari, 2008; Timm-Davis and Fish, 2015). In contrast, O. sinen- sis without a nasal flap may be related to a physical force for the absorption of a fluid in the stagnant water. Thus, it is considered that such a small two nostrils and an acces- sory nasal sac of O. sinensis assist in a water ventilation by the pumping mechanism of the sac. With the above
Fig. 4. A cross section of the accessory nasal sac(NS) of Oryzias sinensis. A, the structure of the NS; B, the epithelium consisting of stratified epithelial cells(SEC) and mucous cells(arrowhead). Mucous cells are positive to hematoxylin and eosin(B), Alcian blue-Periodic acid Schiff(pH 2.5)(C) and Masson’s trichrome(D) stainings. AB, axon bundle. Bars indicate 200μm in A and 50μm in B to D, respectively.
A B
C
D
mechanical function, the gross olfactory structure of O.
sinensis also can be used for a taxonomic study(Saru- watari et al., 1997; Cao et al., 2012).
O. sinensis prefers flowing to stagnant water in farming canals and streams(Kim and Park, 2002). So, there is a high possibility that the habitat can be typically polluted by nutrients, nitrogen and phosphorus accumulated on.
In addition, the water condition is possible to affect seri- ously the olfactory cells by hazard chemical substances.
Therefore the olfactory structure is considered a suitable protection strategy to overcome any adverse environ- ments. The mosquitofish Gambusia affinis and Japanese rice fish O. latipes, though they have similar habitats to O.
sinensis, has been known to have an olfactory chamber lacking a lamellae and an islet distribution of a sensory epithelium, which is to minimize an interaction with out- side materials(Yamamoto, 1982).
In particular, the single accessory nasal sac and mucous cell are likely related to its environments. The pumping of a nasal sac has been generally known to assist fish’s water ventilation via the olfactory organ(Doving et al., 1977).
So, it might let the olfactory organ detect new odors from the outside and discharge remaining odors and residues in the olfactory chamber. In addition, the mucous film made by various mucous cells plays the following roles on: the reduction of a friction force for water ventilation, cell protection against bacterial and physical particles, osmoregulation and ion exchange(Shephard, 1994). In general, the mucous cell on the olfactory epithelium has been known not be tested positive to H-E staining(Ghosh and Chakrabarti, 2011, 2014). However, our result con- firmed that in the accessory nasal sac mucous cells are stained with a weak blush color to H-E staining and are also AB-PAS and Masson’s trichrome staining positive, which strongly suggest the mucous cell is rich in glyco- proteins. From this microscopic study of O. sinensis, therefore, it hints that the olfactory organ is suitable to a standing(stagnant or lentic) water system, in particular polluted water.
referenceS
Cao, L., R. Causee and E. Zhang. 2012. Revision of the loach spe- cies Barbatula nuda(Bleeker 1865)(Pisces: Balitoridae) from North China, with a description of a new species from Inner Mongolia. Zootaxa, 3586: 236-248.
Cox, J.P. 2008. Hydrodynamic aspects of fish olfaction. J. Roy. Soc.
Interface, 5: 575-593.
Døving, K.B., M. Dubois-Dauphin, A. Holley and F. Jourdan. 1977.
Functional anatomy of the olfactory organ of fish and the ciliary mechanism of water transport. Acta Zool., 58: 245- 255.
Ghosh, S.K. and P. Chakrabarti. 2009. Cellular architecture and functional aspects of the olfactory rosette of Wallago attu (Bleeker). Turkish J. Fish Aquat. Sci., 9: 187-190.
Ghosh, S.K. and P. Chakrabarti. 2011. Distribution and organiza- tion of different cells lining the olfactory epithelium of the Indian minor carp, Labeo bata(Hamilton 1822): a light and scanning electron microscopic analysis. Pakistan J. Biol.
Sci., 14: 736-741.
Ghosh, S.K. and P. Chakrabarti. 2014. Histophysiological studies on the olfactory epithelium of banded pearl spot Etroplus suratensis(Bloch, 1790). J. Entomol Zool. Stud., 2: 78-82.
Gurr, G.T. 1956. A practical manual of medical and biological stain- ing techniques. Interscience, New York, pp. 1-99.
Hansen, A. and E. Zeiske. 1998. The peripheral olfactory organ of the zebrafish, Danio rerio: An ultrastructural study. Chemi- cal Senses, 23: 39-48.
Hara, T.J. 1986. Role of olfaction in fish behaviour. In: Pitcher, T.J.
(ed.), The behaviour of teleost fishes. Springer US, pp. 152- 176.
Kasumyan, A.O. 2004. The olfactory system in fish: structure, func- tion, and role in behavior. J. Ichthyol., 44: 180-223.
Kim, H.T. and J.Y. Park. 2014. Structure of the egg envelope in rice-fish Oryzias sinensis Chen, Uwa & Chu, 1989, with comments on the relationship to Oryzias latipes. J. Appl.
Ichthyol., 30: 156-159.
Kim, H.T., H.S. Kim and J.Y. Park. 2016. The anatomy and histol- ogy of the olfactory organ in the Korean sand goby Favo- nigobius gymnauchen(Pisces, Gobiidae). Korean J. Ichthy- ol., 28: 29-35.
Kim, H.T., Y.J. Lee, J.S. Park and J.Y. Park. 2014. A study on the structure of peripheral olfactory organ in the Korean mud- skipper, Scartelaos gigas(Pisces, Gobiidae). Korean J. Ich- thyol., 26: 281-287.
Kim, I.S. and J.Y. Park. 2002. Freshwater fishes of Korea. Kyo-Hak Publishing Co. Ltd., Korea. pp. 388-389.(in Korean).
Kumari, K. 2008. Morphology and morphometry of the olfactory rosette of a teleostean fish: Catla catla(Ham.). Our Nature, 6: 30-37.
Mokodongan, D.F., R. Tanaka and K. Yamahira. 2014. A new rice- fish of the Genus Oryzias(Beloniformes, Adrianichthyidae) from Lake Tiu, Central Sulawesi, Indonesia. Copeia, pp.
561-567.
Mowry, R.W. 1956. Alcian blue technique for the hisochemical study of acidic carbohydrates. J. Histochem. Cytochem., 4:
407-408.
Ojha, P.P. and A.S. Kapoor. 1973. Structure and function of the ol- factory apparatus in the freshwater carp, Labeo rohita Ham.
Buch. J. Morphol., 140: 77-85.
Parenti, L.R. 2008. A phylogenetic analysis and taxonomic revision of ricefishes, Oryzias and relatives(Beloniformes, Adrian-
ichthyidae). Zool. J. Linn. Soc., 154: 494-610.
Sarkar, S.K. and S.K. De. 2011. Functional morphoanatomy of olfactory sensory epithelial cells of Pseudapocryptes lance- olatus(Bloch and Schneider). I.J.S.N., 2: 1-6.
Saruwatari, T., J.A. Lopez and T.W. Pietsch. 1997. A revision of the osmerid genus Hypomesus Gill(Teleostei: Salmoniformes), with the description of a new species from the southern Ku- ril Islands. Species Diversity, 2: 59-82.
Shephard, K.L. 1994. Functions for fish mucus. Rev. Fish. Biol.
Fisheries, 4: 401-429.
Timm-Davis, L.L. and F.E. Fish. 2015. Flow through the nasal cav- ity of the spiny dogfish, Squalus acanthias. Eur. Phys. J.
Special Topics, 224: 3407-3417.
Yamamoto, M. 1982. Comparative morphology of the peripheral olfactory organs in teleosts. In: Hara, T.J.(ed.), Chemore- ception in fishes; Elsevier, Amsterdam, pp. 39-59.
Zeiske, E., B. Theisen and H. Breucker. 1992. Structure, develop- ment, and evolutionary aspects of the peripheral olfactory system. In: Hara T.J.(ed.), Fish chemoreception; Springer Netherlands, pp. 13-39.
한국산 대륙송사리 Oryzias sinensis (Pisces: Adrianichthyidae) 후각기관의 해부 및 조직화학적 연구
김현태
·
이용주1·
박종영전북대학교 자연과학대학 생물학과, 1전주교육대학교 과학교육과
요 약 : 한국산 대륙송사리 Oryzias sinensis의 후각기관을 실체현미경과 광학현미경을 이용하여 해부 및 조직
화학적 특징들을 조사하였다. 전체적 구조에서 후각기관은 두부의 주둥이 위에 한 쌍으로 존재하였으며 다소 떨어
져있는 전비공과 후비공, 한 개의 비강, 한 개의 비낭으로 구성되었다. 비강내 상피는 감각상피와 비감각상피층으
로 구분되었다. 감각상피는 후감각세포, 지지세포, 기저세포 그리고 공포들로 구성되었다. 비감각상피는 층상상피
세포, 산성과 중성 점액세포들로 구성되었다. 비낭의 상피층은 층상상피세포와 배상세포들로 구성되었다. 결과적으
로 이러한 대륙송사리의 후각기관의 해부 및 조직화학적 특징들은 농경지에서의 정체되고 오염되어있는 수환경을
반영하는 것으로 사료된다.
찾아보기 낱말 : 비공, 비강, 비낭, 점액세포