45 책임저자:김정상, 702-701, 대구시 북구 산격동 1370
경북대학교 응용생명과학부 Tel: 053-950-5752, Fax: 053-950-6750 E-mail: [email protected]
접수일:2010년 1월 8일, 1차수정일: 2010년 1월 11일, 2차수정일: 2010년 1월 13일, 게재승인일:2010년 1월 15일
Correspondence to:Jong-Sang Kim
Department of Life and Food Sciences, Kyungpook National University, 1370, Sangyeok-dong, Buk-gu, Daegu 702-701, Korea
Tel: +82-53-950-5752, Fax: +82-53-950-6750 E-mail: [email protected]
Effect of GRP78 Overexpression on Antiproliferative Activity of Anticancer Agents against Breast Cancer Cells
Young Ran Ha1, In-Ae Lee1, Hyo Jung Kim1, Hee Jung Kang2, Dae Hwan Nam1, Jia Park1 and Jong-Sang Kim1
1School of Applied Biosciences and Department of Animal Science and Biotechnology, Kyungpook National University, Daegu 702-701, 2Department of Human Ecology, Kyungsung University, Busan 608-736, Korea
Our previous study showed that genistein, a tyrosine kinase inhibitor and one of the major isoflavones in soybean, induced the expression of glucose regulated protein 78 (GRP78) which is one of the heat shock proteins in both MCF-7 and MDA-MB-231 cells. Pretreatment with genistein caused the increased resistance to doxorubicin in mammary tumor MCF-7 cells while it sensitized estrogen receptor (ER)-negative mammary tumor MDA-MB-231 cells to the drug. To address the discrepancy in response to the drug between two different cell lines pretreated with genistein, the transfectants expressing high level of GRP78 were generated by transfecting with pcDNA3.1 vector carrying GRP78 gene. Over- expression of GRP78 did not affect the cell morphology but modulated the expression of a set of genes involved in cell expressing different levels of GRP78 survival in cell type-specific manner. The transfectants were used to evaluate whether the level of GRP78 regulates the resistance or sensitivity against anti-cancer drugs including doxorubicin, tamoxifen, paclitaxel, mitomycin C. Overexpression of GRP78 made MCF-7 cells more resistant to doxorubicin and mitomycin C, and it slightly sensitized MDA-MB-231 cells to tamoxifen and paclitaxel. In conclusion, the cellular level of GRP78 or any exogenous factor affecting GRP78 expression may alter the efficacy of anticancer drugs on mammary tumor cells. (Cancer Prev Res 15, 45-51, 2010)
Key Words: Doxorubicin, Glucose-regulated protein78 (GRP78), Anticancer drugs, Mammary tumor, Drug resistance
INTRODUCTION
The glucose-regulated protein 78 (GRP78) is a molecular chaperone 78 kDa protein which is also known as BiP, the immunoglobulin heavy chain binding protein.1,2) GRP78 resides in the lumen of endoplasmic reticulum where it functions as a molecular chaperone in an ATP-dependent manner. In several culture systems, GRP78 expression is dramatically enhanced under a variety of stressful conditions including glucose deprivation, ER Ca2+ depletion, blockade of glycosylation, oxidative stress, and hypoxia.3,4) Overexpression of GRP78 is prominent in a wide variety of tumors and protects tumor cells
against ER stress as well as a range of anticancer agents.5~9) The potential mechanisms responsible for this protection include preventing protein misfolding, binding of ER Ca2+, and blocking activation of caspases and proapoptotic proteins associated with the ER.10~12) Downregulation of GRP78 has been shown to reverse these cytoprotective effects.11) For instance, knockdown of GRP78 sensitizes glioma cells to 5-fluorouracil and CPT-11.13)
Our previous study has shown that genistein, one of isoflavones present in soybean and tyrosine kinase inhibitor, induced GRP78 in mammary tumor MCF-7 and MDA-MB- 231 cell lines.14) Interestingly the compound increased sensiti- vity to anticancer drug doxorubicin in ER-negative mammary
tumor MDA-MB-231 cells while it conferred resistance to MCF-7 cells, estrogen-positive mammary tumor cells. In this study we attempted to determine whether overexpression of GRP78 itself affected sensitivity of mammary tumor cells to various anticancer drugs.
MATERIALS AND METHODS 1. Materials
All cell culture reagents and fetal bovine serum were obtained from Gibco BRL (Gaitherburg, MD, USA). Estrogen receptor-positive and negative mammary tumor cells (MCF-7, MDA-MB-231) were from Korean Cell Line Bank (Seoul, Korea). All other chemicals were of reagent grade.
2. Cell culture
MCF-7 (ER-positive) and MDA-MB-231 (ER-negative) hu- man mammary tumor cells were plated at a density of 3×105 cells per 100 mm plate (Nunc, Roskilde, Denmark) in 10 ml of Eagle's Minimal Essential Medium (EMEM, Gibco BRL) supplemented with 10% FBS (Hyclone, Logan, Utah, USA).
The plates were normally incubated for 3∼4 days in a humidified incubator in 5% CO2 at 37°C.
3. Cytotoxicity assay
MCF-7 or MDA-MB-231 cells (4×103 per well) and their transfectants overexpressing GRP78 were seeded into 96-well plates (Nunc), cultured in fresh EMEM containing 10% FBS for 24 h. After removing the media and washing the cells with PBS, cells were incubated in fresh medium containing various concentrations of doxorubicin (0, 5, 10, 25, 50, 100 nM), paclitaxel (0, 1, 5, 10, 20 μM), and tamoxifen (0, 2.5, 5, 10, 20 μM) for another 48 h, following determining cell survival by sulforhodamine B (SRB, Sigma) assay.15)
4. DNA cloning and overexpression
The full-length cDNA of GRP78 was cloned human GRP78 by RT-PCR using primers corresponding to the sequence NM_005347 (related sequences: BC 020235, AF216292, AJ2717292). Plasmids encoding hGRP78 was constructed by PCR-based amplification and purified PCR product was ligated into TOPO vector. The construct was then transformed into expression host E. coli DH5α. The insert fragment was relea- sed by restriction digestion and subcloned into expression
vector pcDNA3.1(−) (Invitrogen, Carlsbad, California, USA).
Forward and reverse primers with BamH1 and Xho1 restriction sites, corresponding to 5' and 3' ends of transcripts, were used to amplify the desired GRP78 gene from mRNA of MCF-7 and MDA-MB-231 tumor cells. The sequences of primers designed based on the sequence of the GRP78 gene with the restriction sites BamHI and XhoI were as follows:
GRP-F; 5'-CAA CTG GAT CCC AAG ATG AAG CTC TCC-3'
GRP-R; 5'-GCA CTC GAG GAT CAG TGT CTA CAA CTC ATC-3'
PCR was performed according to the following thermocycle conditions: initial denaturation at 95°C for 2 min; 35 cycles of denaturation at 95°C for 1 min; annealing at 64°C for 1 min; and extension at 72°C for 2 min. The amplified PCR product was eluted from the gel using Qiagen gel extraction kit (Qiagen-Korea, Seoul, Korea) to remove unused primers and dNTPs. The purified PCR product was ligated into a TOPO vector. The construct was then transformed into expres- sion host E. coli DH5α. The insert fragment was released by restriction digestion and subcloned into pcDNA3.1 (Invitrogen, Calsbad, CA). E. coli strain DH5α was used for cloning and maintaining the constructs. The recombinant expression plas- mid was subsequently verified by DNA sequencing done by SolGent Co. Ltd. (Seoul, Korea).
5. Transfection
MCF-7 and MDA-MB-231 cells were plated on 6-well cul- ture plates (at a density of 3×105 cells/well) in normal growth medium (EMEM, 10% FBS). After an overnight incubation, cells were washed with media, then serum free fresh EMEM
+plasmid DNA 0.5 μg/μl (GRP78)+2.8 μl polyMAC (Chemicell, Berlin, Germany) was added to each well contai- ning cells. And then, the cell culture plates were incubated with positioning on the Magneto-FACTOR plate (Chemicell, Berlin, Germany) for 15 min. After removing the Magneto- FACTOR plate, the cell culture plates were replaced with fresh medium. After incubation for 48 h, the cells were transferred to fresh medium containing the neomycin (G418) for selection.
6. Western blotting
Western blotting was performed on cytosolic fractions pre- pared from cultured cells to estimate the level of GRP78 accor- ding to a protocol described previously.16) Electrophoresis was
Fig. 1. Morphology and GRP78 levels of mammary tumor cells transfected with GRP78 gene. MCF-7 (A) and MDA-MB-231 (B) cells were transfected with pcDNA3.1 vector carrying GRP78 gene construct. The levels of GRP 78 in transfectants were compared from each other by Western blot.
carried out in a Bio-Rad Mini-Protein II Cell apparatus (Bio- Rad Co, Hercules, CA) using a discontinuous buffer system.
Aliquots (∼30 μg of protein) of cytosolic fraction of cells was loaded onto 12% SDS-polyacrylamide gel, and transferred onto polyvinylidene fluoride (PVDF, Gelman Lab, Ann Arber, MI, USA) membranes in transfer buffer (48 mM Tris, 39 mM glycine, 20% methanol). The membrane was blocked with blocking buffer containing 5% skim milk in 0.1% Tween 20 in PBS (PBS/T) overnight at 4°C. The antibody against GRP78 (200 μg/ml), obtained from Santa Cruz Biotechnology Inc.
(Santa Cruz, CA, USA), was diluted by 1:2,000 with dilution buffer (5% skim milk in PBS/T), added to the membrane, and incubated for 4 h at room temperature. After washing 6 times with PBS/T, blots were incubated in washing buffer (PBS/T) with horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature, followed by washing 8 times with PBS/T. Washed blots were exposed onto a film. Antibody against rat β-tubulin (Santa Cruz Co., Santa Cruz, CA) was used as a loading control.
7. Statistical analysis
Data were expressed mean±SD. The statistical analysis bet- ween groups was evaluated using a one-way ANOVA followed by Duncan’s multiple range tests. All the analysis was per- formed with SPSS 12.0 software (SPSS Co. Ltd., Chicago, IL, USA). A value of p<0.05 was regarded as statistically signi- ficant.
RESULTS 1. Overexpression of GRP78
To induce overexpression of GRP78 both MCF-7 and MDA- MB-231 cells were transfected with vector carrying grp78 gene construct. The transfectants showed the 1.5-fold increase in the level of GRP78 protein, compared to control cells, without any significant morphological change as shown in Fig. 1.
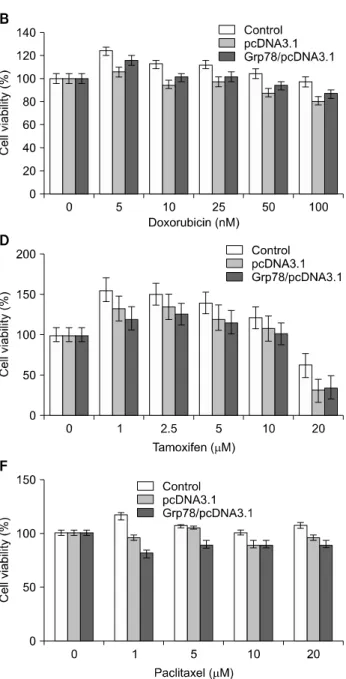
Fig. 2. Effect of GRP78 overexpression on cytotoxicity of anticancer agents for MCF-7 (A, C, E, G) and MDA-MB-231 (B, D, F) cells. Cells were plated into 6-well plate at the concentration 3×105 cells/plate. After incubation for 24 h, media were changed into the fresh one containing various concentrations of anticancer agent and incubated for 24 h, following viability assay by SRB method. Data are mean +/−
S.D. of triplicate assays per experiment.
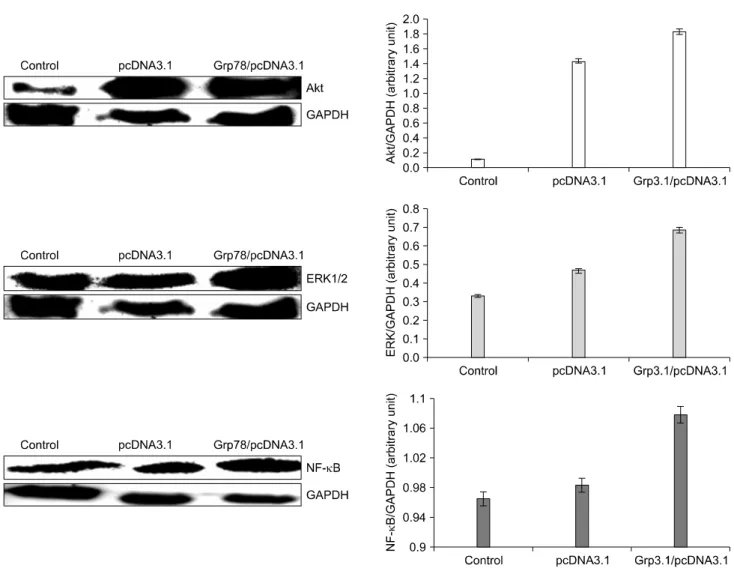
Fig. 3. The effect of GRP78 overexpression on the expression of Akt, ERK1/2, NF-κB in MCF-7 cells. Human mammary tumor MCF-7 cells were plated into plate (100 mm) at the concentration 3×105 cells/plate and incubated for 72 h. Homogenized cell lysates were prepared and the expression of Akt, Erk1/2, and NF-kappa-B was measured by Western blot.
2. Effect of overexpression of GRP78 on drug resistance
To determine the effect of GRP78 overexpression on activi- ties of anticancer drugs, mammary tumor cells (MCF-7 and MDA-MB-231) and their transfectants overexpressing GRP78 were exposed to doxorubicin, mitomycin C, paclitaxel, and tamoxifen, and then subjected to SRB assay for determining cell survival rates. As shown Fig. 2, overexpression of GRP78 conferred significantly enhanced resistance to doxorubicin in MCF-7 cells and in lesser extent in MDA-MB-231 cells. How- ever, increased GRP78 level in both mammary tumor cells did not affect antiproliferative activities of tamoxifen and paclitaxel in both cell lines and even tended to enhance drug sensitivity.
For instance, the growth of MDA-MB-231 cells was more efficiently suppressed by tamoxifen and paclitaxel when GRP78 was overexpressed, compared to control cells.
3. Effect of GRP78 overexpression on intracellu- lar levels of Akt, Erk1/2, and NF-κB
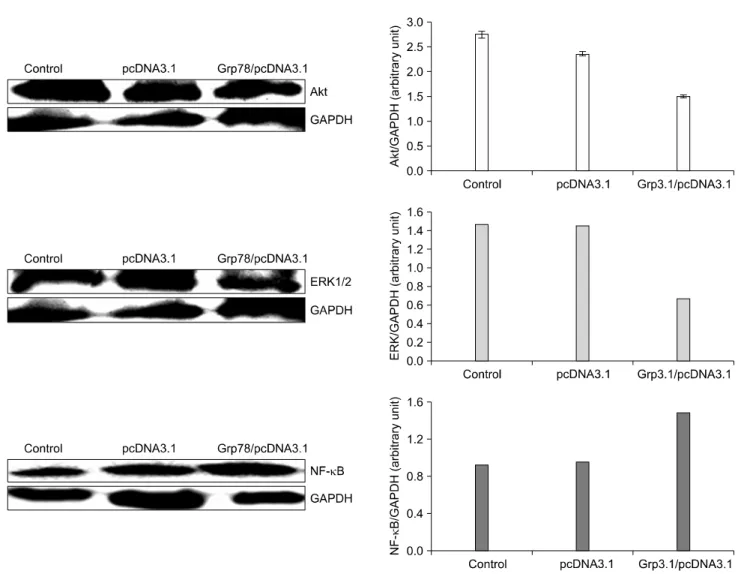
Tumor cells manage to survive via several survival mechani- sms such as MAPK/Erk and PI3K/Akt signaling pathways under stressed conditions. We examined whether overexpres- sion of GRP78 leads to activation of some cell survival pathways. As shown in Fig. 3 and 4, overexpression of GRP78 increased the expression of Akt, Erk1/2, and NF-kappaB in MCF-7 but downregulated Akt and Erk in MDA-MB-231 cells.
Fig. 4. The effect of GRP78 overexpression on the expression of Akt, ERK1/2, NF-κB in MDA-MB-231 cells. Human mammary tumor MDA-MB-231 cells were plated into plate (100 mm) at the concentration 3×105 cells/plate and incubated for 72 h.
Homogenized cell lysates were prepared and the expression of Akt, Erk1/2, and NF-κB was measured by Western blot.
DISCUSSION
Tumor cells usually developed altered protein profile from normal cells. Furthermore, treatment with antiproliferative agents causes extensive change in protein expression. Our pre- vious proteomic study demonstrated that GRP78, one of the heat shock proteins, was significantly upregulated in mammary tumor MCF-7 and MDA-MB-231 cells exposed to genistein, one of tyrosine kinase inhibitors and soy isoflavones.14) It has been reported that increased level of heat shock protein(s) lead to drug resistance of cancer cells.17,18) The present study showed that overexpression of GRP78 would not always increase re- sistance to anticancer drugs at least in mammary tumor models
used in the study. For instance, mammary tumor MDA-MB- 231 cells overexpressing GRP78 were more sensitive to tamoxi- fen and paclitaxel than control cells although the cells become more resistant to doxorubicin than the control. This observation is consistent with decreased levels of Akt and Erk in MDA- MB-231 cells with increased expression of GRP78 (Fig. 4). The factors known to induce GRPs include ER Ca2+ depletion, misfolded protein, reductive stress, glycosylation antagonists, hypoxia, glucose starvation, embryonic development.18) However, this is the first report that soy isoflavones could induce GRP 78 and thereby alter sensitivity to several anticancer drugs.
It is not clear how GRP78 upregulation caused enhanced sensitivity to some anticancer agents, in particular, tamoxifen.
It seems to be associated with decreased levels of Akt and Erk by overexpression of GRP78. Again, the mechanism how overexpression of GRP78 leads to downregulation of Akt and Erk needs to be further investigated.
ACKNOWLEDGEMENT
This work was supported by the Korea Research Foundation Grant, the Korean Government (KRF-2008-532-F00018 and F00069).
REFERENCES
1) Fernandez PM, Tabbara SO, Jacobs LK, Manning FCR, Tsangaris TN, Schwartz AM, Kennedy KA, Patierno SR.
Overexpression of the glucose-regulated stress gene GRP78 in malignant but not benign human breast lesions. Breast Cancer Res Treat 59, 15-26, 2006.
2) Schroder M, Kaufnam RJ. ER stress and the unfolded protein response. Mutat Res 569, 29-63, 2005.
3) Koumenis C, Wouters BG. "Translating" tumor hypoxia:
UPR-dependent and independent pathways. Mol Cancer Res 4, 423-436, 2006.
4) Kaufman RJ. Orchestrating the unfolded protein response in health and disease. J Clin Invest 110, 1389-1398, 2002.
5) Calderwood SK, Khaleque MA, Sawyer DB, Ciocca DR. Heat shock proteins in cancer. Trends Biochem Sci 31, 164-172, 2006.
6) Tiligada E. Chemotherapy: induction of stress responses.
Endocr-Relat Canc 13, 115-124, 2006.
7) Kim R, Emi M, Tanabe K, Murakami S. Role of the unfolded protein response in cell death. Apoptosis 11, 5-13, 2006.
8) Li J, Lee AS. Stress induction of GRP78/BiP and its role in cancer. Curr Mol Med 6, 45-54, 2006.
9) Virrey JJ, Dong D, Stiles C, Patterson JB, Pen L, Ni M, Schönthal AH, Chen TC, Hofman FM, Lee AS. Stress chaperone GRP78/BiP confers chemoresistance to tumor- associated endothelial cells. Mol Cancer Res 6, 1268-1275, 2008.
10) Sacchi CM, Schiaffonati L. The effect of etopside (VP-16) on mouse L fibroblasts: modulation of stress response, growth and apoptosis genes. Anticancer Res 16, 3659-3664, 1996.
11) Sugawara S, Takeda K, Lee A, Dennert G. Suppression of stress protein GRP78 induction in tumor B/C10ME eliminates resistance to cell mediated cytotoxicity. Cancer Res 53, 6001- 6005, 1993.
12) Chatterjee S, Hirota H, Belfi CA, Berger NA. Hypersensitivity to DNA cross-linking agents associated with up-regulation of glucose-regulated stress protein GRP78. Cancer Res 57, 5112- 5116, 1997.
13) Pyrko P, Shonthal AH, Hofman FM, Chen TC, Lee AS. The unfolded protein response regulator GRP78/BiP as a novel target for increasing chemosensitivity in malignant gliomas.
Cancer Res 67, 9809-9816, 2007.
14) Lim HA, Kim JH, Kim HJ, Sung MK, Kim MK, Yoon Park JH, Kim JS. Genistein induces glucose-regulated protein 78 in mammary tumor cells. J Med Food 9, 28-32, 2006.
15) Skehen P. New colorimetric cytotoxicity assay for anticancer- drug screening. J Nat Cancer Institute 82, 1107-1112, 1990.
16) Ausbel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Short protocols in molecular biology. 4th ed. Johns and Sons, Inc, New York, pp10-40∼10-48, 1999.
17) Reddy RK, Mao C, Baumeister P, Austin RC, Kaufman RJ, Lee AS. Endoplasmic reticulum chaperone protein GRP78 protects cells from apoptosis induced by topoisomerase inhi- bitors: role of ATP binding site in suppression of caspase-7 activation. J Biol Chem 278, 20915-20924, 2003.
18) Lee AS. The glucose-regulated proteins: stress induction and clinical applications. Trends Biochem Sci 26, 504-510, 2001.