Journal of Bacteriology and Virology 2014. Vol. 44, No. 2 p.162 – 169 http://dx.doi.org/10.4167/jbv.2014.44.2.162
Microbial Profile of the Stomach: Comparison between Normal Mucosa and Cancer Tissue in the Same Patient
Incheol Seo, Bijay Kumar Jha, Seong-Il Suh, Min-Ho Suh and Won-Ki Baek* Department of Microbiology, School of Medicine, Keimyung University, Daegu, Korea
Gastric cancer is the third most common cancer and the third most frequent cause of cancer mortality in Asia. It is predicted that gastric cancer will remain an important cause of death at least during the next half century because of the increasing number of new cases in an aging population. However, little has been revealed about the role of gastric microbes and their reaction to gastric cancer. In this study, we identified differences in the microbial communities between gastric cancer and normal gastric mucosa by comparing the microbiomes of tissues from the same patients. The clustering analysis results showed different bacterial communities between normal gastric mucosa and gastric cancer. A comparison of bacterial communities at the species level revealed that Helicobacter pylori was significantly reduced in cancer tissue compared to that in normal gastric mucosa in the same patient. A comparison at the genus level showed that Propionibacterium spp., Staphylococcus spp., and Corynebacterium spp. had significantly reduced populations in cancer tissue, whereas Clostridium spp. and Prevotella spp. had significantly increased populations in cancer tissue.
Key Words: Gastric cancer, Helicobacter pylori, Microbiome
INTRODUCTION
The stomach was considered sterile for a long time because of the acidic environment until Helicobacter pylori was found in the 1980s (1). Since then, H. pylori have been identified as a major causative agent of gastric adenocar- cinoma (2, 3). In addition, the presence of microorganisms besides H. pylori has been demonstrated based on culture- dependent methods. New non-culturable microbes have been discovered through DNA-based approaches (4, 5).
However, we have limited knowledge about the stomach microbiome, as these organisms are highly diverse between patients and among experimental studies (6). Contamination
of ingested microorganisms from the mouth and throat also makes study difficult (7).
Gastric cancer is the third most common cancer and the third most frequent cause of cancer mortality in Asia (8).
Many reports are available about the mechanism by which H. pylori induces gastric adenocarcinoma (9), but little is known about the role of other microorganisms in stomach cancer incidence (10). Moreover, not much is known about changes in the microbial communities including H. pylori after the occurrence of gastric cancer. Many studies have focused on changes in the microbiome due to increased gastric pH from proton-pump inhibiting drugs (11, 12) or atrophic gastritis as a result of H. pylori infection (13).
Dicksved et al. investigated gastric microbiota from 10
162
Original Article
Received: April 11, 2014/ Revised: May 23, 2014/ Accepted: May 26, 2014
*Corresponding author: Won-Ki Baek, M.D., Ph.D. Department of Microbiology, Keimyung University School of Medicine, 1095 Dalgubeol-daero, Dalseo-Gu, Daegu, 704-701, Korea.
Phone: +82-53-580-3843, Fax: +82-53-580-3788, e-mail: [email protected]
○CCThis is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/license/by-nc/3.0/).
patients with gastric cancer and compared them with gastric microbiota from five dyspeptic control patients who had normal gastric mucosa morphology (14). However, because of the high diversity in the gastric microbiomes between individuals, it was difficult to assess the effect of gastric cancer on the microbiome. Therefore, in this study, we identified differences in microbial communities between gastric cancer and normal gastric mucosa by comparing them from the same patients using a culture-independent method.
MATERIALS AND METHODS Data source
Thirty-two RNA sequencing data of gastric cancer and adjacent normal gastric mucosa from 16 patients in Korea
were obtained from NCBI Sequence Read Archive with accession number SRP014574. Detailed information about the patients and samples were referenced from the publi- cation by Yoon (15), and described in Tables 1 and 2. From the analysis of normal gastric mucosa in 16 patients, 11 were considered H. pylori carriers whose H. pylori bit score accounted for more than 10% of the total bit score;
thus, the sequence analysis results of those 11 patients were used for further statistical testing.
Bacterial sequence detection from the raw data The computational subtraction method was applied to detect the pathogen from sequence data (16, 17) with a modification. Raw data sequences were aligned to the hg19 human reference genome (NCBI) using TopHat (18). A total of 62~88% of all reads was mapped to the human
Table 1. Clinical information for the samples used in the analysis.
Patient
IDa Sexa Agea Locationa Size (cm)a Histologic subtypea
(WHO classification) Laurena Stagea HP (%)b Selectc
43 M 51 U 7.0 WD Gastric IIIb 2.7
80 M 56 L 5.0 PD Diffuse IIIb 1.3
87 M 53 U 8.0 MD Diffuse IIIb 0
95 M 65 L 6.0 WD Intestinal IIIa 31.1 O
119 M 48 L 10.0 WD Intestinal IIIa 11.7 O
130 M 57 L 6.5 MD Intestinal IIb 0
134 M 57 L 8.5 WD Intestinal IIIb 31.8 O
135 M 37 L 6.0 Mucinous Intestinal IIb 92.0 O
136 M 55 L 4.5 MD Diffuse IIIa 17.5 O
195 M 67 U 6.0 Papillary Intestinal IIIa 30.4 O
236 F 72 L 6.5 MD Mixed IIa 68.1 O
849 M 64 L 5.4 PD Intestinal Ib 63.8 O
859 F 60 L 5.0 MD Intestinal Ib 74.8 O
882 F 75 L 3.5 MD Intestinal Ia 57.4 O
889 F 66 L 8.7 MD Intestinal IIa 56.6 O
917 M 73 L 7.9 MD Intestinal IIa 6.5
Information marked with 'a' were obtained from 'Yoon et al., Comprehensive genome- and transcriptome-wide analyses of mutations associated with microsatellite instability in Korean gastric cancers, Genome Res 23 (2013) 1109-1117. HP(%)b, percentage of Helicobacter pylori population from entire blast results; Selectc, selected patients for analysis accroding to the presence of Helibacter pylori in the normal gastric mucosa; U, upper third; L, lower third; WD, tubular adenocarcinoma and well differentiated; MD, tubular adenocarcinoma and moderately differentiated; PD, tubular adenocarcinoma and poorly differentiated.
Table 2. Summary of the sequence statistics.
SRA Runa Patient
IDb Sampleb Total
readsc Unmapped
with hg19d Unmapped
read (%)e Contigsf Matching with
bacteriag Above thresholdh SRR546237 43 Normal 52,088,890 13,775,741 26.4 98,994 1,722 246 SRR546236 43 Tumor 52,088,890 12,407,493 23.8 33,734 388 49 SRR825140 80 Normal 43,269,450 9,637,598 22.3 41,315 514 172 SRR546238 80 Tumor 42,996,076 12,289,307 28.6 69,175 5,567 532 SRR546239 87 Normal 44,471,834 14,136,479 31.8 93,583 1,227 190 SRR546240 87 Tumor 49,814,912 12,612,740 25.3 70,767 3,171 227 SRR825143 95 Normal 49,357,002 10,892,291 22.1 52,590 613 295 SRR825141 95 Tumor 46,801,172 8,781,017 18.8 39,856 138 69 SRR546226 119 Normal 52,088,890 14,958,865 28.7 62,186 2,739 327 SRR546225 119 Tumor 55,022,224 16,085,525 29.2 35,455 135 58 SRR825136 130 Normal 46,515,690 9,967,511 21.4 32,930 142 75 SRR825135 130 Tumor 49,181,346 10,449,965 21.2 34,317 117 58 SRR546227 134 Normal 52,400,004 12,290,105 23.5 31,281 500 97 SRR546228 134 Tumor 55,022,224 13,180,350 24.0 34,657 252 91 SRR546230 135 Normal 52,088,890 11,573,353 22.2 68,630 1,766 997 SRR546229 135 Tumor 52,266,668 10,269,450 19.6 47,252 166 43 SRR546232 136 Normal 54,755,556 14,774,615 27.0 66,951 845 177 SRR546231 136 Tumor 53,511,112 10,454,368 19.5 26,586 166 66 SRR546233 195 Normal 53,511,112 12,381,166 23.1 55,855 305 107 SRR546234 195 Tumor 52,088,890 14,704,878 28.2 47,228 499 94 SRR825139 236 Normal 41,204,902 9,277,569 22.5 44,692 857 498 SRR825137 236 Tumor 43,956,792 6,135,987 14.0 20,651 192 89 SRR801424 849 Normal 58,124,992 10,104,003 17.4 139,959 216 54 SRR801425 849 Tumor 63,880,118 7,752,423 12.1 27,819 90 6 SRR801427 859 Normal 59,019,350 10,010,433 17.0 114,625 356 104 SRR801426 859 Tumor 56,227,424 6,982,330 12.4 86,906 188 57 SRR801428 882 Normal 63,549,430 9,720,473 15.3 119,616 355 150 SRR801429 882 Tumor 59,102,148 7,322,912 12.4 75,218 354 137 SRR801430 889 Normal 44,498,444 7,450,469 16.7 88,745 412 122 SRR801431 889 Tumor 59,655,378 7,622,162 12.8 25,708 881 134 SRR801432 917 Normal 62,755,892 7,906,885 12.6 74,454 382 131 SRR801433 917 Tumor 63,306,020 8,839,289 14.0 131,114 413 162 SRA Runa, Run number of each sample in the NCBI Sequence Read Archive; Information marked with 'b' were obtained from 'Yoon et al., Comprehensive genome- and transcriptome-wide analyses of mutations associated with microsatellite instability in Korean gastric cancers, Genome Res 23 (2013) 1109-1117. Total readsc, Total number of sequenced reads; Unmapped with hg19d, Number of unmapped reads after aligning with hg19 human reference genome using TopHat; Unmapped read (%)e, Percentage of unmapped reads to total sequenced reads; Contigsf, Number of contigs de novo assembled by Vetvet; Matching with bacteriag, Number of contigs which were matched with bacterial reference genomes; Above thresholdh, Number of contigs with E-value ≤1 × E-40 and identity ≥95% to the reference sequence among matched contigs with bacterial reference genomes.
genome from each sample (Table 2). The unmapped reads were assembled by Velvet (19) to produce a longer sequence that allowed a strict match with the bacterial genome. The assembly produced 20,651~139,959 contigs from each sample. A bacterial reference database was built using 2,646 sets of the bacterial reference genome (NCBI) to match the bacterial sequences from the assembled reads.
Then, the contigs were aligned to the bacterial sequences using NCBI's nucleotide megablast against the bacterial
reference database. Only one query with the lowest E-value and highest bit score was extracted from each matched contig to the reference. Among the extracted queries, hits with E-values > 1 × E-40 and identity < 95% to the reference sequence were removed. The total bit score was calculated and used in the analysis by summing all bit scores of the remaining hits according to genus and species.
Statistical analysis
Bray-Curtis dissimilarity was used with R statistical software and the Vegan package (20) for the cluster analysis.
The Wilcoxon signed-rank test was performed by pairing results of the cancer and normal samples from the same patient to analyze differences in the microbiomes between gastric cancer and normal gastric mucosa. P < 0.05 was considered significant.
RESULTS
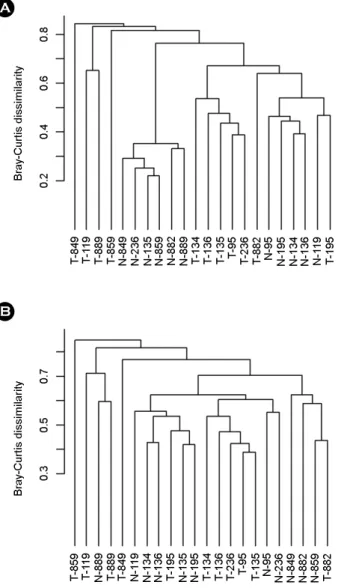
Twenty-two RNA-sequencing samples were used for statistical analysis to find differences in the microbiomes between gastric cancer and adjacent normal gastric mucosa from 11 patients who had a high H. pylori sequence number in normal mucosa from the results of de novo assembly among 16 patients. A total of 350 bacterial species were detected from all 32 samples (data not shown). The clustering analysis results for the bacterial populations obtained by BLASTing the assembled non-host sequences from each sample showed differences in the bacterial communities between normal gastric mucosa and that of gastric cancer (Fig. 1). A similar clustering result was observed when the H. pylori sequences were removed from the blast results to exclude statistical error because H. pylori was the dominant bacterium in the 11 selected patient samples.
Comparing bacterial communities at the species level, H.
pylori decreased significantly in cancer tissue compared to that in normal gastric mucosa from the same patient (Table 3, Fig. 2). Comparison at the genus level after removing the H. pylori sequences from the blast results showed that Propionibacteirum spp., Staphylococcus spp., and Coryne- bacterium spp. decreased significantly in cancer tissue A
B
Figure 1. Clustering of the samples based on bacterial com- position at the species level. Bacterial composition is different between normal gastric mucosa and gastric cancer. Y-axis represents the Bray-Curtis dissimilarity. (A) Whole bacterial species were used for analysis including Helicobacter pylori. (B) Clustering analysis result after excluding H. pylori.
mucosa, whereas Clostridium spp. and Prevotella spp.
increased significantly in cancer tissue mucosa (Fig. 3 and 4).
DISCUSSION
The incidence and mortality of gastric cancer continues to decrease worldwide, but it is predicted that gastric cancer will remain an important cause of death at least during the next half century because of the increasing number of new
cases in an aging population (21, 22). The role of H. pylori in gastric cancer tumorigenesis has been studied extensively.
In contrast, little has been revealed about the role and reaction of other gastric microbes in patients with gastric cancer. Information about the stomach microbiome has been accumulating continuously. The increased use of Table 3. Microbiota with different populations between normal
gastric mucosa and that of gastric cancer.
Genera p-value
Helicobacter pylori 0.000977
Propionibacterium spp. 0.013672
Staphylococcus spp. 0.014433
Clostridium spp. 0.034611
Prevotella spp. 0.036032
Corynebacterium spp. 0.044011
Significantly different microbiota in the population by comparing normal gastric mucosa and gastric cancer from the same patient.
p-value for Helicobacter pylori was analyzed by comparing all strains at the species level. Other bacterial genera were analyzed at the genus level after removing all H. pylori sequences from the blast results. The Wilcoxon signed-rank test was applied for statistical analysis.
Figure 2. Relative abundances of Helicobacter pylori in normal gastric mucosa and that of gastric cancer. Normal gastric mucosa has a higher abundance of Helicobacter pylori than that of gastric cancer. Y-axis represents the relative proportion of H. pylori to all bacterial species from each sample. 'N' denotes normal gastric mucosa, 'T' denotes gastric cancer. Numbers are identification codes for each sample; the same identification code means they were obtained from the same patient.
A
B
C
Figure 3. More abundant bacterial genera in normal gastric mucosa than that of gastric cancer. Presence of (A) Propioni- bacterium spp., (B) Staphylococcus spp., and (C) Corynebacterium spp. in normal gastric mucosa and that of gastric cancer. Y-axis represents the relative proportion of each bacterial genus to all bacterial genera from each sample. 'N' denotes normal gastric mucosa, 'T' denotes gastric cancer. Numbers are identification codes for each sample; the same identification code means they were obtained from the same patient.
gastroendoscopy for diagnostic and therapeutic purposes and advances in high-throughput sequencing methods have enabled a considerable amount of genetic information to be obtained in a short time (1, 23). Despite these advances, information about the gastric cancer microbiome has been insufficient until now. Therefore, in this study, we investi- gated the microbiome by matching non-host sequences from sequencing data of the human stomach to the bacterial reference genomes to elucidate differences in the micro- biomes between normal gastric mucosa and that of gastric cancer.
The samples we used were obtained by gastrectomy with immediate harvest of specimens under an aseptic condition (15). Thus, they were free from contamination through the biopsy channel of the throat and mouth (1).
Furthermore, we included all non-culturable bacteria for
investigation because the analysis was performed based on bacterial sequences. However, the purpose of sample pre- paration and sequencing was not aimed at assessing the bacterial sequence. Nevertheless, several studies have demonstrated that this approach is valid to study micro- organisms (16, 17, 24, 25). Moreover, this is the first study that has considered differences in the microbiomes between gastric cancer and normal mucosa of the stomach from the same patient using a culture-independent method rather than analyzing samples obtained from different patients with gastric cancer and healthy volunteers. Thus, we provide valuable insight about changes in the gastric cancer microbiome.
We only included samples from H. pylori carriers who had the H. pylori sequence in their normal gastric mucosa to focus on the relationships between H. pylori and other microbiota in the mucosa. H. pylori was absent or was significantly reduced in cancer tissue mucosa (Fig. 2). This result is consistent with a study showing a decrease in the H. pylori population in cancer tissue by measuring the relative abundance of terminal restriction fragments (14).
Changes in the mucosal environment makes it difficult for H. pylori to colonize cancer tissue because gastric adeno- carcinoma originates from the mucosa and becomes hypo- chlorhydric (26), whereas H. pylori colonizes gastric mucosa due to chemotaxis driven by pH (27, 28).
According to the clustering results of Fig. 1A, after removing H. pylori (Fig. 1B), the normal mucosa samples were clustered with normal mucosa samples, and gastric cancer samples were clustered with gastric cancer samples rather than normal to cancer in the same patient. Normal gastric mucosa had larger populations of Propionibacterium spp., Staphylcoccus spp., and Corynebacterium spp. than those of gastric cancer (Fig. 3). Propionibacterium in- habiting the stomach should be highly resistant to acidic conditions (29), but growth of Staphylococcus and Coryne- bacterium is suppressed in an acidic environment (30).
Some evidence indicates that elevated gastric pH due to H.
pylori-induced inflammation predisposes colonization by environmental microbiota (31). Moreover, ammonia and bicarbonate produced by H. pylori from urea can be used A
B
Figure 4. More abundant bacterial genera in gastric cancer than those in normal gastric mucosa. Presence of (A) Clostridium spp. and (B) Prevotella spp. in normal gastric mucosa and that of gastric cancer. Y-axis represents the relative proportion of each bacterial genus to all bacterial genera from each sample. 'N' denotes normal gastric mucosa, 'T' denotes gastric cancer. Numbers are identification codes for each sample; the same identification code means they were obtained from the same person.
as substrates by other bacteria (32). In contrast, Clostridium and Prevotella were denser in gastric cancer mucosa than that in normal mucosa of the stomach (Fig. 4). This result was consistent with previous reports showing that patients with gastric carcinoma harbor higher numbers of Clostridium (32) and Prevotella (14). Clostridium increases significantly as gastric pH increases (11), and Clostridium preferentially accumulates in tumors (33). Furthermore, Prevotella can be affected by the presence of H. pylori because Prevotella spp.
are found only in H. pylori-negative gerbils (13). Therefore, we concluded that the changes in the mucosal layer due to gastric cancer such as increased pH are a major factor involved in the changes of the gastric cancer microbiome.
Further investigation is needed to determine whether the existence of H. pylori between normal gastric mucosa and gastric cancer affects composition of the gastric microbiome.
The influence of H. pylori on the composition of stomach microbiome remains controversial (4, 34, 35).
The microbial species obtained from the 32 samples seemed to contain normal flora of the mouth and throat, bacteria from the environment and food, and major stomach bacteria, as discussed above. Further research on the gastric cancer microbiome will be needed to identify the involve- ment of microorganisms in suppressing or inducing gastric cancer. This may help in the prevention and treatment of gastric cancer by comparing the gastric cancer microbiome with that of normal gastric mucosa.
REFERENCES
1) Yang I, Nell S, Suerbaum S. Survival in hostile territory:
the microbiota of the stomach. FEMS Microbiol Rev 2013;37:736-61.
2) No authors listed. Schistosomes, liver flukes and Helico- bacter pylori. IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Lyon, 7~14 June 1994. IARC Monogr Eval Carcinog Risks Hum 1994;
61:1-241.
3)Parsonnet J, Friedman GD, Vandersteen DP, Chang Y, Vogelman JH, Orentreich N, et al. Helicobacter pylori infection and the risk of gastric carcinoma. N Engl J Med 1991;325:1127-31.
4) Bik EM, Eckburg PB, Gill SR, Nelson KE, Purdom EA, Francois F, et al. Molecular analysis of the bacterial microbiota in the human stomach. Proc Natl Acad Sci U S A 2006;103:732-7.
5) Hu Y, He LH, Xiao D, Liu GD, Gu YX, Tao XX, et al.
Bacterial flora concurrent with Helicobacter pylori in the stomach of patients with upper gastrointestinal diseases. World J Gastroenterol 2012;18:1257-61.
6) Maldonado-Contreras A, Goldfarb KC, Godoy-Vitorino F, Karaoz U, Contreras M, Blaser MJ, et al. Structure of the human gastric bacterial community in relation to Helicobacter pylori status. ISME J 2011;5:574-9.
7)Stearns JC, Lynch MD, Senadheera DB, Tenenbaum HC, Goldberg MB, Cvitkovitch DG, et al. Bacterial biogeography of the human digestive tract. Sci Rep 2011;1:170.
8) Albarracin VH, Pathak GP, Douki T, Cadet J, Borsarelli CD, Gärtner W, et al. Extremophilic Acinetobacter strains from high-altitude lakes in Argentinean Puna:
remarkable UV-B resistance and efficient DNA damage repair. Orig Life Evol Biosph 2012;42:201-21.
9)Wroblewski LE, Peek RM Jr. Helicobacter pylori in gastric carcinogenesis: mechanisms. Gastroenterol Clin North Am 2013;42:285-98.
10)Wang ZK,Yang YS. Upper gastrointestinal microbiota and digestive diseases. World J Gastroenterol 2013;19:
1541-50.
11) Amir I, Konikoff FM, Oppenheim M, Gophna U, Half EE. Gastric microbiota is altered in oesophagitis and Barrett's oesophagus and further modified by proton pump inhibitors. Environ Microbiol 2013.
12)Williams C, McColl KE. Review article: proton pump inhibitors and bacterial overgrowth. Aliment Pharmacol Ther 2006;23:3-10.
13)Osaki T, Matsuki T, Asahara T, Zaman C, Hanawa T, Yonezawa H, et al. Comparative analysis of gastric bacterial microbiota in Mongolian gerbils after long- term infection with Helicobacter pylori. Microb Pathog 2012;53:12-8.
14)Dicksved J, Lindberg M, Rosenquist M, Enroth H, Jansson JK, Engstrand L. Molecular characterization of the stomach microbiota in patients with gastric cancer and in controls. J Med Microbiol 2009;58:509-16.
15) Yoon K, Lee S, Han TS, Moon SY, Yun SM, Kong SH,
et al. Comprehensive genome- and transcriptome-wide analyses of mutations associated with microsatellite instability in Korean gastric cancers. Genome Res 2013;
23:1109-17.
16)Isakov O, Modai S, Shomron N. Pathogen detection using short-RNA deep sequencing subtraction and assembly. Bioinformatics 2011;27:2027-30.
17)Xu Y, Stange-Thomann N, Weber G, Bo R, Dodge S, David RG, et al. Pathogen discovery from human tissue by sequence-based computational subtraction.
Genomics 2003;81:329-35.
18) Trapnell C, Pachter L, Salzberg SL. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 2009;
25:1105-11.
19) Zerbino DR, Birney E. Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res 2008;18:821-9.
20) Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB, et al. vegan: Community Ecology Package. 2013 [cited 2014 Mar 3]. Available from:
http://CRAN.R-project.org/package=vegan/.
21)Leja M, Wex T, Malfertheiner P. Markers for gastric cancer premalignant lesions: where do we go? Dig Dis 2012;30:268-76.
22) Yeh JM, Hur C, Schrag D, Kuntz KM, Ezzati M, Stout N, et al. Contribution of H. pylori and smoking trends to US incidence of intestinal-type noncardia gastric adenocarcinoma: a microsimulation model. PLoS Med 2013;10:e1001451.
23) Loman NJ, Misra RV, Dallman TJ, Constantinidou C, Gharbia SE, Wain J, et al. Performance comparison of benchtop high-throughput sequencing platforms. Nat Biotechnol 2012;30:434-9.
24) Feng H, Taylor JL, Benos PV, Newton R, Waddell K, Lucas SB, et al. Human transcriptome subtraction by using short sequence tags to search for tumor viruses in conjunctival carcinoma. J Virol 2007;81:11332-40.
25)Rathi B, Sarangi AN, Trivedi N. Genome subtraction for novel target definition in Salmonella typhi. Bio- information 2009;4:143-50.
26) Lauwers GY, Srivastava A. Gastric preneoplastic lesions and epithelial dysplasia. Gastroenterol Clin North Am 2007;36:813-29.
27)Amieva MR, El-Omar EM. Host-bacterial interactions in Helicobacter pylori infection. Gastroenterology 2008;
134:306-23.
28) Salama NR, Hartung ML, Müller A. Life in the human stomach: persistence strategies of the bacterial pathogen Helicobacter pylori. Nat Rev Microbiol 2013;11:385 -99.
29)Delgado S, Suárez A, Mayo B. Identification, typing and characterisation of Propionibacterium strains from healthy mucosa of the human stomach. Int J Food Microbiol 2011;149:65-72.
30) Giannella RA, Broitman SA, Zamcheck N. Gastric acid barrier to ingested microorganisms in man: studies in vivo and in vitro. Gut 1972;13:251-6.
31) Oh JD, Kling-Bäckhed H, Giannakis M, Engstrand LG, Gordon JI. Interactions between gastric epithelial stem cells and Helicobacter pylori in the setting of chronic atrophic gastritis. Curr Opin Microbiol 2006;9:21-7.
32)Wu WM, Yang YS, Peng LH. Microbiota in stomach:
New insights. J Dig Dis 2014;15:54-61.
33)Forbes NS. Engineering the perfect (bacterial) cancer therapy. Nat Rev Cancer 2010;10:785-94.
34) Tan MP, Kaparakis M, Galic M, Pedersen J, Pearse M, Wijburg OL, et al. Chronic Helicobacter pylori infection does not significantly alter the microbiota of the murine stomach. Appl Environ Microbiol 2007;73:1010-3.
35)Martin ME, Bhatnagar S, George MD, Paster BJ, Canfield DR, Eisen JA, et al. The impact of Helico- bacter pylori infection on the gastric microbiota of the rhesus macaque. PLoS One 2013;8:e76375.