Article
http://dx.doi.org/10.4217/OPR.2017.39.4.279
한국 서해 문치가자미(Marbled Sole, Pseudopleuronectes yokohamae)의 성숙과 성장
황선완
1,2· 황학빈
3· 황선도
4*1한국해양과학기술원 생태기반연구센터 (15627) 경기도 안산시 상록구 해안로 787
2한국과학기술연합대학원대학교 해양생물학과 (34113) 대전광역시 유성구 가정로 217
3서대문자연사박물관
(03718) 서울특별시 서대문구 연희로32길 51
4한국수산자원관리공단 서해지사 (54021) 전라북도 군산시 해망로 256
Maturity and Growth of Marbled Sole, Pseudopleuronectes yokohamae in the West Sea of Korea
Sun-Wan Hwang
1,2, Hakbin Hwang
3, and Sun-Do Hwang
4*1Marine Ecosystem and Biological Research Center, KIOST Ansan 15627, Korea
2Department of Marine Biology, Korea University of Science and Technology Daejeon 34113, Korea
3Seodaemun Museum of Natural History Seoul 03718, Korea
4Korea Fisheries Resources Agency Gunsan 54021, Korea
Abstract : We determined the maturity and growth of the Marbled sole, Pseudopleuronectes yokohamae based on monthly sampling in the West Sea of Korea from February 2009 to December 2010.
Determination of sex of P. yokohamae was by gonadal inspection, and age and growth were determined by analyzing the otolith. The biological minimum size of P. yokohamae female was 24.5 cm. The seasonal changes in the ratio of a translucent zone to an opaque zone revealed that the end of the translucent zone of the otolith were annuli formed in May once a year. We compared 8 type growth equations with several length-at-age data. By using the length-at-monthly age data of these, the calculated von Bertalanffy growth equations were L
t= 41.7(1 − e
−0.47(t + 0.24)) for females and L
t= 32.7(1 − e
−0.75(t + 0.01)) for males. We estimated that 24.5 cm was the total length of 2 year old P. yokohamae spawns in the cold water season from January to April, and grows to 8 years old for females and 6 years old for males in the West Sea of Korea.
Key words : Pseudopleuronectes yokohamae, Otolith microstructure, age, growth, maturity
*Corresponding author. E-mail : [email protected]
1. 서 론
문치가자미(Pseudopleuronectes yokohamae)는 가자미 목(Pleuronectiformes) 가자미과(Pleuronectidae)에 속하는 어류로 한국의 연근해, 황해, 일본의 홋가이도 이남 해역 및 동중국해 북부 해역에 분포하며, 수심 100 m 이내의 모래질이나 펄질의 바닥에서 주로 생활하는 저어류이다 (Masuda et al. 1984; Kim et al. 2005). 충남 태안군 백사 장 수협의 2008년 어획통계자료에 의하면, 문치가자미 어 획량은 81.5 MT로 가자미류 어획량(91.7 MT)의 88.9%
를 차지하여 한국의 서해 중부 연안에서 어획되는 가자미 류에서 양적으로나 상업적으로 중요한 위치를 차지하는 종이다.
문치가자미에 관해서는 난에서 치어기의 성장에 따른 형태 변화(김 등 1983), 식성(곽과 허 2003; 허 등 2012), 생식기구 및 개체군 동태(강 등 1985; 이 등 1985), 성숙 과 산란(Shafieipour et al. 2004; Kume et al. 2006; Tanda et al. 2008b; 서 등 2010; 김 등 2016) 및 자원량 해석(박 과 청 1991) 등의 다양한 연구가 보고될 정도로 관심대상 종이다. 특히, 문치가자미의 자원평가에 기초자료로 활용 되는 연령과 성장에 관한 논문이 다수 보고되었다 (Tianxiang and Shengmin 1986; Solomon et al. 1987; 김 등 1991; Dagang et al. 1992; Tanda et al. 1992, 2008a;
Tsujino et al. 1997; 박 1997; 문과 이 1999; Kume et al.
2006; Lee et al. 2009b; 남 2013).
그러나 이제까지 보고된 문치가자미의 연령과 성장에 관한 연구들은 대부분 1세 이하 자료가 부족하여 역추산 (back-calculation) 방법을 통해 저연령군의 체장을 추정하 였는데, 이와 같은 방법은 부화 이후 미성숙 시기의 체장 자료가 포함되지 않아 전체 생활사에 대한 성장양상을 반 영하기 어렵고, 성장계수(K)가 과소평가될 수 있는 단점 이 있다(Beckman et al. 1990; Rodriguez and Hammann 1997; Hwang et al. 2008a). 그러므로 문치가자미의 재가 입군의 상태 파악과 어린 개체 관리를 위해서는 고등어 (Hwnag et al. 2008a) 나 흰베도라치(Hwnag et al. 2008b) 에서와 같이 미성어를 포함한 전 생애의 성장양상을 살펴 보는 것이 필요하다.
또한, 이들 문치가자미의 연령과 성장에 관한 이전의 연구결과들 중에는 연륜의 기준인 윤문 설정을 불투명대 에서 투명대로 이행하는 경계로 한 경우(Tianxiang and Shengmin 1986; 김 등 1991; Lee et al. 2009a, 2009b; 남 2013) 와 투명대에서 불투명대로 이행하는 경계로 한 경우 ( 강 등 1985; Solomon et al. 1987; 박 1997; Kume et al.
2006; Tanda et al. 2008a)가 있어 일치성이 없다. 문치가 자미의 이석은 핵을 중심으로 불투명대가 형성되고 성장 하면서 투명대와 불투명대가 교대로 형성되는데, 윤문 판
독을 불투명대 끝으로 할 경우와 투명대 끝으로 할 경우 에 따라 연령 사정결과가 다르게 나타난다(김 등 1991).
결과적으로 윤문 판독과 설정 방법에 따라 하나의 연륜이 많거나 적어지는 결과가 발생하고, 이는 문치가자미의 자 원상태를 평가하고 관리하는데 오류를 범할 수 있음을 의 미한다. 따라서 연령 분석을 위한 윤문 판독 기준을 명확 히 할 필요가 있다.
본 연구에서는 문치가자미 성어의 연령-체장 자료에 문 과 이 (1999)에 의해서 추정된 부화 시 체장과 채집 시기 별 유어의 체장 자료를 추가하여 von Bertalanffy 성장모 델(von Bertalanffy 1938)의 매개변수를 추정하였으며, 이 결과를 문치가자미의 연령과 성장에 관한 기존의 국내외 연구 결과와 비교하였다.
2. 재료 및 방법
자료수집 및 처리
본 연구에 사용된 문치가자미는 2009년 2월부터 2010년 12 월까지 매월 1회 서해 중부해역에서 자망이나 기선저인 망에 포획되어 충남 태안군 백사장 수협위판장에 양륙된 어획물에서 무작위로 추출하여 수집하였다. 단, 2009년 9 월에는 백사장항에 어획물이 없어 대신 인천항에 양륙된 문치가자미를 수집하였다. 수집된 시료는 실험실로 운반 하여 체장(TL: total length, 전장)과 체중(BW: body weight) 및 생식소의 중량(GW: gonad weight)을 측정한 후, 생식소 발달 단계를 파악하고 이석을 추출하였다. 체 장, 체중 및 생식소 중량은 각각 0.1 cm, 1.0 g, 0.01 g 단 위로 측정하였으며, 생식소의 발달 단계는 5단계(미숙, 중 숙, 완숙, 방중, 방후)로 구분하였다(김 1997). 생식소의 중 량과 발달단계 분석은 2009년 2월부터 2010년 1월까지의 자료만을 대상으로 하였다.
연령형질로 추출한 이석은 표면의 유기물과 수분을 제
거하여 실리콘 이석 고정틀에 이석의 둥근 면이 아래로
향하게 올려놓고, 경화제를 첨가한 에폭시 수지(Castolite
Resin, BUEHLER, U.K.) 를 부어 약 2일간 굳힌 후 실리
콘 이석 고정틀을 제거하였다. 이와 같이 제작된 이석 시
료는 핵의 위치와 윤문 판독이 가능할 때까지 시상면으로
연마기(METASERVE 2000, BUEHLER, U.K.)를 이용하
여 갈아내고, 연마포에서 1 µm의 alumina powder로 광택
을 내었다. 이석의 미세구조는 광학현미경의 반사광을 이
용하여 관찰하였다(Fig. 1). 본 연구에서 윤문은 투명대에
서 불투명대로 이행하는 경계를 기준하였으며, 장반경
(OR: otolith radius)과 각 윤경(r
n)은 핵으로부터 성장이
빠른 전단부 방향으로 가장 긴 축을 기준으로 mm 단위로
측정하였다. 연령사정은 2회 이상 반복하였으며, 일치된
결과만 채택하였다.
자료 분석
산란시기를 파악하기 위해 월별 생식소숙도지수(GSI:
Gonadosomatic index)를 분석하였다. 생식소숙도지수는 GSI = GW/BW × 10
2(GW: 생식소 중량, BW: 체중)으로 구하였다.
2009 년 10월부터 2010년 5월 사이의 암컷의 체장별 성 숙개체(완숙, 방중, 방후 단계)의 비율을 이용하여 로지스 틱 곡선(logistic curve), M
L= 1/[1 + exp(a − bL)] (M
L: 군 성숙도, L: 체장, a와 b는 Ln(1/M − 1) = a − bL 회귀직선 에서 상수와 기울기)을 계산하고, 생물학적최소체장 (L
50) = a/b으로 추정하였다(김 1997).
이석 내에서 윤문이 형성되는 시기와 연간 형성 횟수를 파악하기 위하여 월별 연역지수(MI: Marginal index)는 MI = (OR − r
n)(r
n− r
n−1)
−1(OR: 장반경, r
n: 마지막 윤경,
r
n−1: n −1번째 윤경)으로 구하였다. 성장식은 von Bertalanffy 성장식(VBGE)을 이용하여 자료유형에 따른 4가지 성장 식, 즉 윤문 형성시의 체장을 이석장반경과 체장 간의 회 귀직선식(L
i= a + b × r
i) (L
i: i 번째 연륜이 형성되었을 때 의 체장, r
i: i 번째 연륜까지의 윤경) 및 Fraser-Lee의 식 ((L
i= (r
i/R) × (L − a) + a) (L: 채집 당시의 체장, R: 이석 반경, a: 체장과 이석 반경 간의 회귀관계식(L = a + b × R) 의 상수) (Bagenal and Tesch 1978)으로부터 역추산한 연 령-체장 자료, 채집 당시의 이석으로 사정한 연령(observed yearly age) 에 해당하는 체장 자료, 그리고 채집 당시의 연 령을 월령으로 환산한 나이(observed monthly age)-체장 자료를 이용하였다. 만약 문치가자미가 5월에 연륜이 형 성되고 11월에 채집된 개체가 3세로 연령이 판독되었다 면, 나이는 3.5세(= 3년 + 6개월/12개월)로 환산할 수 있 다. 또한, 이들 자료유형에 t = 0 일 때의 길이(L
0) 를 포함 하지 않았을 경우와 포함하였을 경우를 고려한 성장식을 각각 추정하였다. 부화 할 때(t = 0)의 평균 체장과 부화 후 2개월(t = 0.17), 3개월(t = 0.25) 및 4개월(t = 0.33) 된 유어의 평균 체장은 문과 이 (1999)의 결과를 인용하여 각 각 0.34 cm, 2.26 cm, 2.36 cm 및 3.82 cm를 사용하였다.
이상과 같이 8가지의 연령-체장 자료를 이용한 VBGE 의 매개변수는 통계 프로그램 R (R Core Team 2013)를 이용한 FishR (http://fishr.wordpress.com) 패키지의 비선 형회귀방법 중에서 von Bertalanffy 방법으로 추정하였다.
3. 결 과
체장 조성
본 연구에 측정된 문치가자미는 총 1,363 개체였으며, Fig. 1. Photographs show annual rings of Pseudopleuronectes
yokohamae caught in the Western Sea of Korea, under dark-field reflected light. (a) the translucent zone formed on margin of otolith in winter season (February), (b) the opaque zone formation in summer season (June). Otolith radius (OR) and annuli were measured from the core to the longest anterior margin. Each r
irepresents the annual radius
Fig. 2. Length frequency distribution of Pseudopleuronectes
yokohamae collected from the commercial catch
by a gill net and/or an otter trawl in the West Sea
of Korea from February 2009 to December 2010
암컷(881개체, 64.6%)이 수컷(482개체, 35.4%)에 비하여 차지하는 비율이 높았다(Fig. 2). 체장(TL, 전장) 범위와 평균 체장(±SD)은 암컷은 14.1~51.5 cm와 32.5 cm (±6.18 cm), 수컷은 15.5~49.0 cm와 28.2 cm (±4.37 cm)로, 암 컷이 수컷에 비하여 체장 범위가 넓고 큰 개체들이 많았 다(t-test, p < 0.05).
성 성숙과 생물학적최소체장
문치가자미의 GSI는 암컷과 수컷 모두 겨울에 높고 여 름에 낮은 뚜렷한 계절 변동을 보였다(Fig. 3). 암컷의 GSI 는 2월에 가장 높았고 3월에서 5월 사이에 시간에 따 라 급격히 감소하였다. 이후 6월부터 9월 사이에는 5월보 다 약간 낮은 수준으로 유지되다가 10월과 11월에 이전의 5월보다 높은 수준으로 증가하였고, 이어지는 12월과 익 년 1월에 급격하게 상승하였다. 수컷의 경우는 GSI의 최 대값이 나타난 시기가 암컷(2월)보다 한 달 빠른 1월인 것 을 제외하면, 전체적인 계절(월)적 변동은 암컷과 같은 양 상이었다.
연구기간 동안 수온이 높은 여름철에 문치가자미 암수 생식소의 미숙단계 비율이 높은 반면, 가을부터는 중숙의 비율이 증가하다가 겨울철인 1~2월에 전부 완숙 단계이 었고, 봄철인 4~5월에 방후 단계가 나타났다(Fig. 4).
이상의 결과에서 문치가자미의 산란기는 GSI가 높은 값을 나타내고 완숙과 방중 단계를 보이는 1~4월이며, 주 산란기는 2~3월로 추정된다.
문치가자미 암컷은 전장 20 cm에서 성숙한 개체가 출 현하기 시작하였으며, 개체군 중 50% 성숙 체장인 생물학 적최소체장(L
50) 은 24.5 cm이었다(Fig. 5).
Fig. 3. Monthly variations in the means and 95% CIs (vertical bars) of the gonadosomatic index (GSI) for Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February 2009 to January 2010
Fig. 4. Relative frequency of maturity stages by month for Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February 2009 to January 2010
Fig. 5. Scatter diagram of maturity (M
L) versus total
length (L) for female Pseudopleuronectes yokohamae
sampled in the West Sea of Korea from October
2009 to May 2010. The binary data were fitted to
the logistic function
연령과 성장
문치가자미 이석은 타원형으로 핵을 중심으로 불투명대 가 형성되어 있고, 바깥쪽으로 가면서 투명대와 불투명대 가 번갈아 나타났다(Fig. 1). 겨울철 이석에는 가장자리에
투명대가 형성되어 있었던 반면에 여름철 이석 가장자리 에는 불투명대가 나타났다. 한편, 투명대에서 불투명대로 이행하는 경계가 불투명대에서 투명대로 이행하는 경계보 다 뚜렷하였다.
이석 가장자리의 투명대 비율의 월별 변화 양상을 보
Fig. 6. Monthly variation in proportion having translucent zone and opaque zone in the otolith margin of Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February 2009 to January 2010
Fig. 7. Monthly variations in the mean of the marginal index of Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February 2009 to January 2010. Vertical bars represent the 95%
CIs
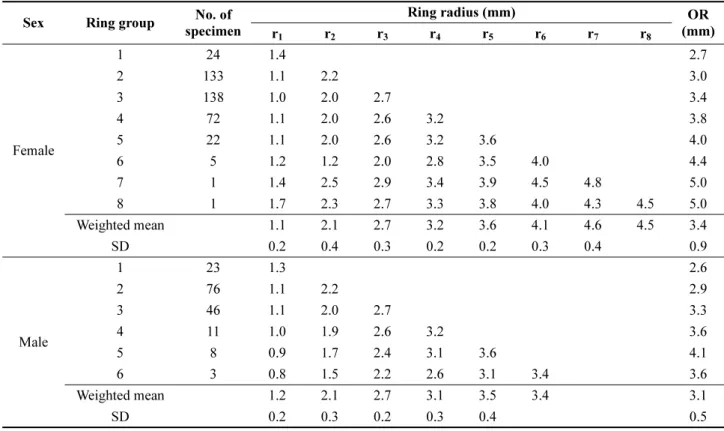
Table 1. Mean ring radius when the each annulus was formed in the otolith of Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February 2009 to December 2010
Sex Ring group No. of specimen
Ring radius (mm) OR
r
1r
2r
3r
4r
5r
6r
7r
8(mm)
Female
1 24 1.4 2.7
2 133 1.1 2.2 3.0
3 138 1.0 2.0 2.7 3.4
4 72 1.1 2.0 2.6 3.2 3.8
5 22 1.1 2.0 2.6 3.2 3.6 4.0
6 5 1.2 1.2 2.0 2.8 3.5 4.0 4.4
7 1 1.4 2.5 2.9 3.4 3.9 4.5 4.8 5.0
8 1 1.7 2.3 2.7 3.3 3.8 4.0 4.3 4.5 5.0
Weighted mean 1.1 2.1 2.7 3.2 3.6 4.1 4.6 4.5 3.4
SD 0.2 0.4 0.3 0.2 0.2 0.3 0.4 0.9
Male
1 23 1.3 2.6
2 76 1.1 2.2 2.9
3 46 1.1 2.0 2.7 3.3
4 11 1.0 1.9 2.6 3.2 3.6
5 8 0.9 1.7 2.4 3.1 3.6 4.1
6 3 0.8 1.5 2.2 2.6 3.1 3.4 3.6
Weighted mean 1.2 2.1 2.7 3.1 3.5 3.4 3.1
SD 0.2 0.3 0.2 0.3 0.4 0.5
면, 2009년 4월 이후 투명대 출현비율이 급격히 감소하고 불투명대가 증가하여 5월부터 9월까지 불투명대가 나타나 는 비율이 높았고, 10월부터는 다시 투명대의 비율이 점 점 증가하였다(Fig. 6). 이석 내 윤문이 형성되는 시기와 그 횟수 또는 주기성을 파악하기 위해 월별 MI의 변동을 분석한 결과(Fig. 7), 윤문은 5월경에 연 1회 형성되는 것
으로 판단되어 연령 판독 기준으로 사용할 수 있었다. 이 는 Fig. 6에서 보여지는 것과 같이 투명대와 불투명대 간 의 상대 비율에 급격한 변화가 일어나는 시기와 일치하여 투명대에서 불투명대로 이행하는 경계 즉, 투명대 끝을 연 륜이 형성되는 윤문으로 판명할 수 있었다. 이 이석의 윤 문을 기준으로 판독한 문치가자미의 최고 연령은 암컷과
Fig. 8. Relationship between otolith radius and total length of Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February to December 2010
Table 2. Mean back-calculated total length at age (L
i) derived by the simple regression of total length on otolith radius of Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February 2009 to December 2010 Sex Age
group
No. of specimen
Back-calculated total length (cm)
L
1L
2L
3L
4L
5L
6L
7L
8Female
1 24 24.8
2 133 23.4 28.5
3 138 22.9 27.5 30.7
4 72 23.4 27.5 30.3 33.1
5 22 23.4 27.5 30.3 33.1 34.9
6 5 23.9 23.9 27.5 31.2 34.4 36.7
7 1 24.8 29.8 31.7 34.0 36.3 39.0 40.4
8 1 26.2 28.9 30.8 33.5 35.8 36.7 38.1 39.0
Weighted mean 23.3 27.8 30.5 33.0 34.9 37.0 39.3 39.0
SD 1.1 1.9 1.4 1.1 0.9 1.3 1.6
Male
1 23 24.3
2 76 23.4 28.5
3 46 23.4 27.5 30.8
4 11 22.9 27.1 30.3 33.1
5 8 22.5 26.2 29.4 32.6 34.9
6 3 22.0 25.2 28.5 30.3 32.6 34.0
Weighted mean 23.4 27.9 30.5 32.5 34.3 34.0
SD 0.8 1.3 1.0 1.5 1.6
수컷에서 각각 8세와 6세로 사정되었고, 2세와 3세의 비 율이 각각 68.4%와 73.1%로 나타나 전체에서 차지하는 비율이 높았다. 이석의 핵으로부터 각 윤문까지의 평균 윤 경을 산출한 결과는 Table 1과 같다. 이석 장반경(R)과 체 장(TL) 간 상관관계(Fig. 8)을 이용하여 윤문이 형성되었 을 때의 체장을 역추산한 연령별 체장과 Fraser-Lee 방법 으로 역추산한 연령별 체장 결과를 각각 Table 2와 Table 3 에 나타내었다. 역추산 연령별 체장 자료를 VBGE에 적
용하여 t = 0 일 때의 길이(L
0) 를 포함하지 않았을 경우와 포함하였을 경우의 문치가자미의 성장 매개변수(L
∞, K, t
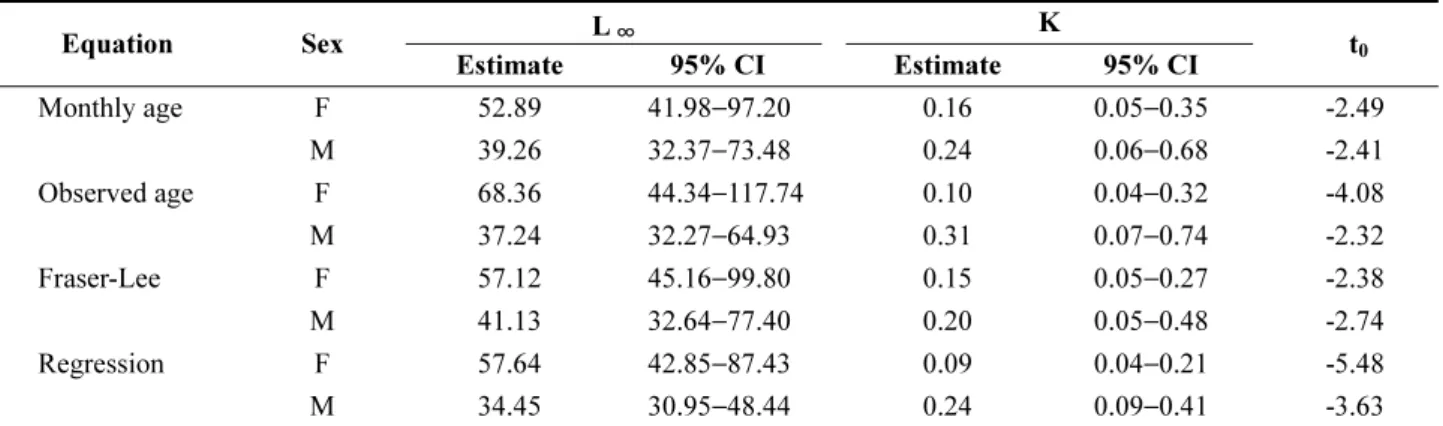
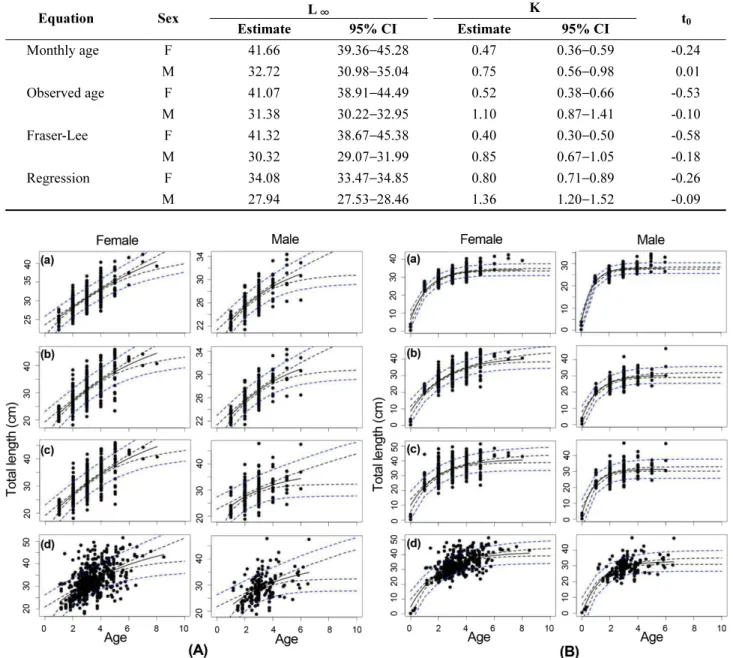
0) 를 Table 4와 Table 5에 각각 추정하고, 각각의 성장곡 선을 Fig. 9에 표현하였다.
문치가자미의 이론적 최대 체장(L
∞) 은 모든 성장 모델 에서 암컷(52.89~68.36 cm)이 수컷(34.15~41.13 cm)에 비하여 크게 추정된 반면에 성장계수(K)는 암컷(0.09~
0.16) 이 수컷(0.20~0.31)에 비하여 작게 추정되었다(Table
Table 3. Mean back-calculated total length at age (L
i) derived by the Fraser-Lee equation of total length on otolith radius of Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February 2009 to December 2010
Sex Age group
No. of Specimen
Back-calculated total length (cm)
L
1L
2L
3L
4L
5L
6L
7L
8Female
1 24 22.9
2 133 22.7 27.0
3 138 22.9 27.5 30.8
4 72 23.8 28.2 31.2 34.1
5 22 24.3 29.1 32.4 35.6 37.7
6 5 25.6 25.6 30.4 35.2 39.4 42.4
7 1 25.7 31.4 33.5 36.1 38.7 41.9 43.5
8 1 26.7 29.7 31.7 34.6 37.1 38.1 39.5 40.5
Weighted mean 23.1 27.6 31.0 34.5 38.0 41.7 41.5 40.5
SD 1.5 1.9 1.1 0.8 1.0 2.4 2.8
Male
1 23 20.9
2 76 21.9 25.5
3 46 22.2 25.3 27.8
4 11 22.0 25.4 28.0 30.2
5 8 21.3 24.0 26.3 28.7 30.3
6 3 22.8 26.7 30.5 32.8 35.5 37.2
Weighted mean 21.9 25.4 27.8 30.0 31.7 37.2
SD 0.7 0.9 1.7 2.1 3.7
Table 4. Estimate and 95% confidential intervals (CIs), calculated without length data (L
0) at t
0, for the von Bertalanffy growth parameters in four equations for Pseudopleuronectes yokohamae sampled in the West Sea of Korea
Equation Sex L
∞K
t
0Estimate 95% CI Estimate 95% CI
Monthly age F 52.89 41.98−97.20 0.16 0.05−0.35 -2.49 M 39.26 32.37 −73.48 0.24 0.06 −0.68 -2.41 Observed age F 68.36 44.34−117.74 0.10 0.04−0.32 -4.08
M 37.24 32.27−64.93 0.31 0.07−0.74 -2.32
Fraser-Lee F 57.12 45.16 −99.80 0.15 0.05 −0.27 -2.38 M 41.13 32.64 −77.40 0.20 0.05 −0.48 -2.74
Regression F 57.64 42.85−87.43 0.09 0.04−0.21 -5.48
M 34.45 30.95 −48.44 0.24 0.09−0.41 -3.63
4 and 5). 또한, 이론적 최대 체장(L
∞) 은 L
0값이 포함되지 않은 경우(암컷: 52.89~68.36 cm, 수컷: 34.45~41.13 cm) 가 포함된 경우(암컷: 34.08~41.66 cm, 수컷: 27.94~
32.72 cm) 에 비해 크게 추정된 반면에 성장계수(K)는 L
0값이 포함되지 않은 경우(암컷: 0.09~0.10, 수컷: 0.20~
0.24)가 포함된 경우(암컷: 0.40~0.80, 수컷: 0.75~1.36) 에 비하여 작게 추정되었다. 이론상 길이가 0 일 때의 시 간 t
0는 L
0값을 포함한 모델(암컷: −0.58 ~ −0.24, 수컷:
−0.18 ~ 0.01)은 “0”에 근사한 결과를 보였는데 비하여 L
0값을 추가하지 않은 모델(암컷: −5.48~−2.38, 수컷:
Table 5. Estimates and 95% confidential intervals (CIs), calculated with length data (L
0) at t
0and additional monthly age-length data during early life period which were refered in Moon and Lee (1999), for the von Bertalanffy growth parameters in four equations for Pseudopleuronectes yokohamae sampled in the West Sea of Korea
Equation Sex L
∞K
t
0Estimate 95% CI Estimate 95% CI
Monthly age F 41.66 39.36−45.28 0.47 0.36−0.59 -0.24
M 32.72 30.98−35.04 0.75 0.56−0.98 0.01
Observed age F 41.07 38.91 −44.49 0.52 0.38 −0.66 -0.53
M 31.38 30.22−32.95 1.10 0.87−1.41 -0.10
Fraser-Lee F 41.32 38.67−45.38 0.40 0.30−0.50 -0.58
M 30.32 29.07 −31.99 0.85 0.67 −1.05 -0.18
Regression F 34.08 33.47−34.85 0.80 0.71−0.89 -0.26
M 27.94 27.53−28.46 1.36 1.20−1.52 -0.09
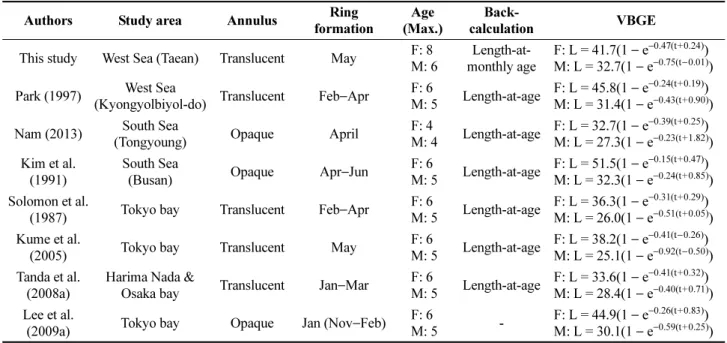
Fig. 9. von Bertalanffy growth curves (solid lines) and their 95% CIs and predicted intervals indicated by inner and
outer dotted lines, respectively derived from back-calculated length at age according to (a) simple regression
(SR), (b) Fraser-Lee equation (FR), (c) the observed length at age and (d) used the observed length at
converted monthly age for Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February
2009 to December 2010. ‘A’ represents the results of VBGE without L
0data, but ‘B’ includes L
0and additional
data during early life period referred in Moon and Lee (1999)
−363~−2.32)은 “0”과 큰 차이를 보였다. 이론상 최대 체 장(L
∞) 과 성장계수(K)의 95% 신뢰구간은 L
0값을 추가하 지 않은 모델이 L
0값을 추가한 모델에 비해 범위가 넓었 다. 한편, L
0값을 추가한 성장 모델에서 이론상 최대 체장 (L
∞)은 단순회귀직선을 이용한 성장식(암컷: L
∞= 34.08 cm, 수컷: L
∞= 27.94 cm) 을 제외하면, 암컷과 수컷은 각 각 41.07~41.66 cm 범위와 30.32~32.72 cm 범위로 성장 식간에 차이가 크지 않았다(Fig. 10).
4. 토 의
성성숙과 생물학적최소체장
황해 중서부 해역에 서식하는 문치가자미의 산란기는 월별 GSI 변화와 성숙단계의 월변화로부터 1월에서 4월 사이로 추정되었다. 또한, 본 연구해역을 포함하여 보다 북쪽에 위치한 보해(Bohai Sea)와 황해 북서부 해역에서 는 문치가자미의 산란이 주로 4월에 이루어진다(Dagang et al. 1992). 반면에 문치가자미는 남해안에 위치한 경남 남해도에서는 12월~1월(서 등 2010), 진해만 11월~12월 ( 문과 이 1999), 그리고 거제 연안에서는 12월~2월(김 등 2006) 에 산란하는 것으로 보고되었다. 또한, 위도상 더 남 쪽인 일본의 동경만(Tokyo Bay)에서는 12월에서 1월 사 이에 문치가자미가 산란하는 것으로 알려졌다(Kume et al. 2006). Aida (1991) 는 어류의 생식소 성숙은 수온이나 광주기와 밀접한 관련이 있다고 제시하였으며, Matsuyama et al. (1991)는 정어리(Sardinops melanosticus)의 GSI 변 동은 수온이 11
oC 로 낮아진 3월에 최대치에 도달하였다가 고수온기인 7월에서 9월 사이에 난소의 퇴화가 일어나 수 온이 생식소의 성숙에 중요한 역할을 한다고 증명하였다.
한편, 이 등 (1985)과 김 등 (2006)은 문치가자미의 GIS 변동과 관련하여 수온 하강이 보상적으로 생식소 성숙을 촉진하여 연중 가장 낮은 수온기에 산란이 이루어진다고 보고하였다.
남해안 문치가자미의 산란기는 11월~2월(10~15
oC) 인 반면, 서해안 문치가자미의 산란시기는 1월~4월(5
oC 내 외)로 상대적으로 수온이 낮은 시기에 산란하는 것으로 나타났고, 같은 시기라도 위도 차이에 따른 해양환경 특성 Fig. 10. Comparing four von Bertalanffy growth equations
for Pseudopleuronectes yokohamae sampled in the West Sea of Korea from February 2009 to December 2010. Parameters of equations were calculated with L
0(length at age 0), referenced in Moon and Lee (1999). F, M, BC indicates female, male and back-calculation, respectively
Fig. 11. Bimonthly variations in bottom water temperature at 7 NIFS (National Institute of Fisheries Science)’s serial oceanographic observation stations (left panel), and location map of NIFS’s serial oceanographic observation stations (right panel). The bottom water temperature in each stations was averaged yearly from 1988 to 2002.
Vertical bars represent standard error. The data were obtained from Korea Oceanographic Data Center of
NIFS (http://kodc.nifs.go.kr)
때문에 수온 역시 차이가 있었다(Fig. 11). 즉, 같은 종이 지만 위도에 따른 해역별 산란시기와 산란수온이 다른 것 을 알 수 있었다.
본 연구 해역에서 채집한 문치가자미 암컷의 생식소 월 변화에서 성숙단계에서 방후 단계가 출현하는 10월에서 다음해 5월까지의 개체를 대상으로 50% 군성숙 을 나타 내는 생물학적최소체장(L
50) 은 24.5 cm이었다(Fig. 5). 반 면에 남해안에 서식하는 문치가자미의 생물학적최소체장 은 남해도 문치가자미 19.8 cm (서 등 2010), 부산 문치가 자미 19.5 cm (강 등 1985), 통영 문치가자미 18.0 cm (남 2013) 로 18.0~19.8 cm 범위의 유사한 크기를 보였지만, 서해안 문치가자미의 생물학적최소체장(24.5 cm)과 큰 차 이를 보였다. 한편, 일본 동경만 문치가자미의 생물학적최 소체장은 13.3 cm로(Kume et al. 2006), 우리나라 문치가 자미의 값과는 더 큰 차이를 보여 해역마다 서식환경의 차이에 따른 다른 산란계군이 존재할 가능성을 제시할 수 있다.
연령과 성장
본 연구에서 서해 중부 연안 문치가자미 최대수명은 암 컷 8세, 수컷 6세까지 출현하였다(Table 6). 일본의 연구에 서 암컷은 6세, 수컷은 5세까지 출현하는 것이 일반적이 었다(Solomon et al. 1987; Kume et al. 2006; Tanda et al.
2008a; Lee et al. 2009a). 이석의 시상면(sagittal plane)과 횡단면(transversal plane)에서 각각 연륜을 판독했을 때, 고연령어에서는 연령 사정의 차이가 생길 수 있지만, 저연 령어에서는 성장 매개변수에 유의한 차이가 나타나지 않
았다(Lee et al. 2009a). 이석의 횡단면을 이용하여 연령을 판독한 남 (2013)의 경우에 문치가자미는 1~4세까지로 판독되었고, 대부분 1~2세의 저연령군에 해당되었다. 본 연구에서는 7세, 8세로 판독된 암컷이 1개체씩만 포함되 었기 때문에 연륜을 과소 측정하였더라도 성장식에서 추 정된 매개변수에는 큰 영향이 없었을 것으로 판단된다. 남 (2013) 의 연구결과와 본 연구에서 7세, 8세로 연령사정 된 암컷의 자료를 제외할 경우, 채집된 문치가자미의 최대연 령은 연륜을 판독하는 이석의 절단면 선택 방법과 채집 지역에 따라서 큰 차이가 나지 않는다.
이석(otolith)을 이용하여 어류의 연령을 사정하는데 있 어 연륜의 정확한 판독이 우선되어야 한다. 이석을 이용하 여 문치가자미의 연령을 사정한 기존의 연구들을 비교하 면, 불투명대에서 투명대로 넘어가는 경계를 연륜으로 판 독하기도 하였고(Tianxiang and Shengmin 1986; 김 등 1991; Lee et al. 2009a, 2009b; 남 2013), 투명대에서 불 투명대로 이행하는 경계를 연륜으로 판독하기도 하여 (Solomon et al. 1987; 박 1997; Kume et al. 2006; Tanda et al. 2008a), 동일 종에 대한 연령사정 기준이 달랐다. 김 등 (1991)은 윤문을 투명대에서 불투명대로 이행하는 경 계로 판독한 강 등 (1985)의 문치가자미 이석 시료를 대상 으로 불투명대에서 투명대로 이행하는 경계를 윤문으로 판독하여 강 등 (1985)이 제시한 최고 연령(암컷: 5세, 수 컷: 3세)보다 1세 많은 결과(암컷: 6세, 수컷: 4세)를 보고 하였다. 이는 윤문 판독 기준에 따라 연령이 달라지고 자 원 평가와 이를 기반으로 한 자원관리방안에 오류를 범할 수 있음을 의미한다. 본 연구에서 월별로 문치가자미의 이
Table 6. Comparison of von Bertalanffy growth equation (VBGE) in the present study and previous studies Authors Study area Annulus Ring
formation
Age (Max.)
Back-
calculation VBGE This study West Sea (Taean) Translucent May F: 8
M: 6
Length-at- monthly age
F: L = 41.7(1 − e
−0.47(t+0.24)) M: L = 32.7(1 − e
−0.75(t−0.01)) Park (1997) West Sea
(Kyongyolbiyol-do) Translucent Feb−Apr F: 6
M: 5 Length-at-age F: L = 45.8(1 − e
−0.24(t+0.19)) M: L = 31.4(1 − e
−0.43(t+0.90)) Nam (2013) South Sea
(Tongyoung) Opaque April F: 4
M: 4 Length-at-age F: L = 32.7(1 − e
−0.39(t+0.25)) M: L = 27.3(1 − e
−0.23(t+1.82)) Kim et al.
(1991)
South Sea
(Busan) Opaque Apr −Jun F: 6
M: 5 Length-at-age F: L = 51.5(1 − e
−0.15(t+0.47)) M: L = 32.3(1 − e
−0.24(t+0.85)) Solomon et al.
(1987) Tokyo bay Translucent Feb −Apr F: 6
M: 5 Length-at-age F: L = 36.3(1 − e
−0.31(t+0.29)) M: L = 26.0(1 − e
−0.51(t+0.05)) Kume et al.
(2005) Tokyo bay Translucent May F: 6
M: 5 Length-at-age F: L = 38.2(1 − e
−0.41(t−0.26)) M: L = 25.1(1 − e
−0.92(t−0.50)) Tanda et al.
(2008a)
Harima Nada &
Osaka bay Translucent Jan−Mar F: 6
M: 5 Length-at-age F: L = 33.6(1 − e
−0.41(t+0.32)) M: L = 28.4(1 − e
−0.40(t+0.71)) Lee et al.
(2009a) Tokyo bay Opaque Jan (Nov−Feb) F: 6
M: 5 - F: L = 44.9(1 − e
−0.26(t+0.83))
M: L = 30.1(1 − e
−0.59(t+0.25))
석을 관찰한 결과, 이석의 가장자리가 겨울철에는 투명대 가 형성되어 있었던 반면에 여름철에는 불투명대를 보였 다. 또한, MI가 5월에 최저값을 보였다(Fig. 7). 더욱이 월 별 GSI 변동으로부터 문치가자미의 산란기가 1~4월로 추정된 점을 고려하면(Fig. 3), 겨울철 산란을 마친 시기인 투명대에서 불투명대로 이행하는 경계를 윤문형성시기로 정하면 만 1년의 연령이 되기 때문에 합리적이다. 따라서 서해 문치가자미의 산란기가 4월까지 이어지고, 윤문형성 시기가 5월인 점을 고려하여 연령의 기준일을 5월 1일로 정하여도 무방하리라 판단된다. 이는 월령을 계산하는데 기준으로 사용하였다.
본 연구에서 VBGE을 적용하여 추정한 문치가자미의 성장식은 암수 간에 뚜렷한 차이를 나타내었다(Fig. 10).
성장 속도는 암컷과 수컷 모두 2세까지는 급격하게 증가 하다가 3세부터는 연령이 증가함에 따라 감소하였으며, 역추산한 체장은 암컷이 같은 연령의 수컷보다 유의하게 큰 것으로 나타났다(t-test, p < 0.05). 이러한 현상은 이 종 에 대한 기존의 국내외 연구 결과들에서도 나타났다(강 등 1985; Tianxing and Shengmin 1986; Solomon et al.
1987; 김 등 1991; Dagang et al. 1992; Tanda et al. 1992, 2008a; 박 1997; Kume et al. 2006; Lee et al. 2009b; 남 2013). 문치가자미와 같은 가자미과에 속하는 어류 중 도다 리(Pleuronichthys cornutus), 물가자미(Eopsetta grigorjewi), 돌가자미(Kareius bicoloratus), 갈가자미(Tanakius kihtarae), 참가자미(Pleuronectes herzensteini), 찰가자미(Microstomus achne) 등의 어류에서도 수컷에 비하여 암컷의 성장이 빠 른 것으로 나타나(Dagang et al. 1992; 남 2013), 가자미과 어류의 일반적인 현상으로 판단된다.
어류의 성장을 나타내는데 널리 이용되는 VBGE은 매 개변수 추정 방법에 따라 성장식도 달라진다(Gallucci and Quinn 1979; Schnute 1981; Francis 1988; Cailliet et al.
2006). VBGE 의 매개변수(L
∞, K, t
0) 추정에 자연에서 채 집한 자치어에서 추정한 부화 시 체장(L
0)이나 실험실에 서 사육하면서 부화할 때(t = 0)의 길이(L
0)를 추가시키면, 1 세 이하의 초기 성장을 성장식에 반영하게 되므로 전 생 애의 성장 양상을 나타낼 수 있다(Hwang et al. 2008a, 2008b). 본 연구에서는 문치가자미 성장식 추정에 문과 이 (1999) 가 제시한 부화 때(t
0) 의 길이(L
0) 와 부화 후 2개월 (t
0.17), 3 개월(t
0.25) 및 4개월(t
0.33) 된 유어의 길이(L
0= 0.34 cm, L
0.17= 2.26 cm, L
0.25= 2.36 cm, L
0.33= 3.82 cm) 를 이용함으로써 초기 성장을 포함한 전 생애의 성장식을 나타내었다. 본 연구의 문치가자미 VBGE에서 t
0는 L
0를 포함하지 않았을 경우에는 −5.48에서 −2.32의 범위로 “0”
에서 많이 벗어났고, 포함하였을 경우에는 –0.58에서 0.01 의 범위로 “0”에 가까운 값을 보였다(Table 4와 5). 또한, L
0를 포함하지 않는 성장식에서는 0세일 때 체장(L
0) 이
16 ~ 22 cm 범위로 계산되어 생물학적으로 상식적이지 않 는 과대치 결과가 도출되었다. 이와 같이 초기성장이 빠르 거나 최대수명이 작은 종에서는 t = 0 일 때의 길이인 L
0를 포함하는 경우와 그렇지 않은 경우에 이들 성장식간 차이가 크게 나타났다(Hwang et al. 2008a). 따라서 본 연 구에서 문치가자미의 성장식은 L
0를 추가시켜 초기 성장 을 포함한 전 생애의 성장 양상을 반영하는 것이 타당하 다고 판단된다. 1세 이전의 저연령어에 대한 자료는 어구 와 어업의 선택성에 따라 얻기 어려운 경우가 생긴다. 따 라서 본 연구와 같이 치어와 유어의 일령을 이용한 부화 시기 및 부화 시 체장 추정치, 어린시기의 월별 체장 자료 는 성장식에서 초기성장을 나타내는데 유용할 수 있다.
윤문 형성시기의 체장을 역추산하는 방법에 따른 성장 모델의 결과를 보면, Fraser-Lee 방법은 이론상 최대 체장 이 실측 체장을 이용한 방법과 월령으로 환산한 자료로 사용한 성장식은 큰 차이가 없었으나, 단순회귀직선식으 로 역추산한 체장을 이용한 성장식이 상대적으로 다르게 나타났다(Fig. 10 and Table 5). 따라서, 이석 반경(R)과 체장(L)의 관계에서 y 절편이 0과 유의하게 다른 경우에 는 Lee 현상을 보정한 Fraser-Lee의 방법으로 연륜 형성 시의 길이(length-at-yearly age)를 역추산할 것을 제안한 다. 또한, 현재 컴퓨터의 연산속도는 모든 실측자료를 포 함하더라도 충분히 계산이 가능하기 때문에 자료의 수가 적은 평균 체장 자료보다 자료의 수가 많은 실측 체장 자 료나 연령을 월로 환산한 월령별 체장(length-at-monthly age) 자료를 이용하는 방법이 통계적으로 더 타당하다는 추세다(Hwang et al. 2008a, 2008b).
서해 중부 연안에서 채집된 문치가자미의 이론상 최대 체장(L
∞) 은 단순회귀직선에 의한 성장식의 값을 제외하고 암컷 41.1~41.7 cm, 수컷 30.3~32.7 cm로 추정되었다.
김 등 (1991)과 남 (2013)은 남해에서 채집된 문치가자미 암컷과 수컷에 대하여 이론상 최대체장(L
∞) 이 각각 51.5 cm 와 32.3 cm 그리고 32.7 cm와 27.3 cm로 추정하 였다. 또한, 박 (1997)은 서해에서 채집된 문치가자미를 대상으로 이론상 최대체장(L
∞) 을 암컷과 수컷에 대하여 각각 45.8 cm와 31.4 cm로 추정하였다. 이렇게 이론상 최 대체장(L
∞)이 다른 것은 4세 이상의 고연령군의 시료 수 가 적어 성장모델을 잘 구현해내지 못했거나, 성장식 계산 방법의 차이, 또는 해역과 시기가 다르기 때문으로 판단된 다. 따라서 해역과 시기에 따른 성장양상의 차이를 알아보 기 위해서는 시료의 성질과 추정방법에서 동일함이 선결 되어야 할 것이다.
그럼에도 불구하고, 문치가자미 이석의 투명대 끝을 연
륜으로 판독한 연구결과만을 비교했을 때, 본 연구와 박
(1997) 의 VBGE의 이론적 최대체장은 암컷에서 40 cm 이
상, 수컷에서 30 cm 이상으로 비슷하게 계산되었으나, 일
본의 연구결과에 비해 대체로 크게 추산되었다(Table 6).
이상에서 본 바와 같이 문치가자미는 해역별로 산란기 와 성장 양상이 다르게 나타남으로써 해역별로 다른 계군 이 존재한다고 판단된다. 따라서 향후 문치가자미 자원을 평가할 때 해역별로 해당 계군의 생물학적 특성치를 사용 해야 함이 요구된다.
사 사
본 연구의 이석 시료 처리와 분석에 대한 해석에 도움 을 준 국립수산과학원 임양재 박사와 박지영 님, 그리고 원고 정리에 도움을 준 한국해양과학기술원 홍지민, 송민 주 연구원께 감사드립니다. 본 논문은 한국해양과학기술 원의 주요사업(PE99566, 미세플라스틱 발생 방지를 위 한 친환경 고압력 부표 개발)에서 일부 지원되었음을 밝 힙니다.
참고문헌