화이트 퀴노아의 발아에 따른 이화학적 특성, In Vitro 전분 소화율 및 항산화 활성
고혜경․이영택 가천대학교 식품생물공학과
Effects of Germination on the Physicochemical Properties, In Vitro Starch Digestibility, and Antioxidative Activity of
White Quinoa (Chenopodium quinoa Willd.)
Hye-Kyung Goh and Young-Tack Lee
Department of Food Science and Biotechnology, Gachon University
ABSTRACT White quinoa (Chenopodium quinoa Willd.) seeds were subjected to germination at 25°C for 2 days and subsequent freeze-drying, and the effects of germination on the physicochemical properties, in vitro starch digesti- bility and antioxidative activity were evaluated. Germination decreased the crude protein, crude fat, and crude ash contents but increased slightly the total carbohydrate content, including starch. The total phenolic contents in water and 80% ethanol extract of quinoa were increased during a 48 h germination period, whereas the total flavonoid contents were decreased slightly. The pasting properties determined using Rapid Visco Analyser (RVA) showed that germination process decreased the pasting viscosities of quinoa significantly, due to the increased amylolytic activities.
In vitro starch hydrolysis (%) of germinated quinoa was somewhat higher than that of raw quinoa. The antioxidative
activity (1,1-diphenyl-2-picrylhydrazyl assay) of germinated quinoa was slightly lower than that of raw quinoa. These results suggest that bioactive components and biological activities may vary according to the structural parts of germinat- ing quinoa seed.
Key words: quinoa, germination, physicochemical properties, in vitro starch digestibility, antioxidative activity
Received 18 September 2018; Accepted 29 October 2018 Corresponding author: Young-Tack Lee, Department of Food Sci- ence and Biotechnology, Gachon University, Seongnam, Gyeonggi 13120, Korea
E-mail: [email protected], Phone: +82-31-750-5565
서 론
퀴노아(Quinoa,
Chenopodium quinoa
Willd.)는 남아메 리카의 안데스 지역이 원산지로 재배되어 내려오는 명아주 과 식물로서 재배적인 적응성이 뛰어나고 우수한 영양 및 기능성으로 인해(1) 전 세계적으로 관심이 높은 식량자원 중 하나이다. 퀴노아 종실은 평균 직경 1~2.5 mm로 쌀보다 작고 둥글납작한 모양을 하고 있으며 화본과 곡류와 구성 성질과 쓰이는 용도가 유사하여 유사 곡류(pseudocereal grain)로 불린다(2,3). 퀴노아 종자의 색은 흰색, 회색, 노란 색, 빨간색 또는 검은색 등으로 다양하다(1). 현재 국내 시중 에 유통되고 있는 퀴노아 제품은 대부분은 수입산이지만 최 근에 국내에서도 퀴노아가 일부 재배되어 생산이 이루어지 고 있다.퀴노아 종자는 일반 곡류보다 많은 단백질, 지질, 섬유소,
무기질을 함유하고 있으며, 특히 아미노산 및 지방산 조성이 매우 뛰어나다(4,5). 퀴노아의 종실은 종피(pericarp), 외배 유(perisperm), 내배유(endosperm), 배아(embryo)로 구 성되어 있으며, 배아가 외배유를 띠처럼 둘러싸고 있는 것이 일반 곡류와 다르다(6). 외배유는 주로 전분으로 이루어져 있고 내배유와 배아에는 단백질, 지질, 무기질, 섬유소가 상 대적으로 많이 함유되어 있다(7). 퀴노아에는 밀단백질인 글 루텐(gluten) 성분이 없기 때문에 글루텐 알레르기를 가진 사람들에게 밀, 호밀, 보리와 같이 글루텐을 함유하는 곡류 를 대체할 수 있는 작물의 하나로도 관심을 받고 있다(8).
퀴노아에는 또한 비타민 A, E, B1, B2, B3 등 각종 비타민과 Ca, P, K, Mg, Fe 등 미네랄 함량이 높으며(9), 폴리페놀, 플라보노이드, 피토스테롤, 토코페롤, 스쿠알렌 등 다양한 생리활성물질을 상당량 함유하고 있다(10,11).
퀴노아는 주로 종실을 이용하고 있으며 쌀과 같이 그대로 끓여서 조리해 섭취하거나 밀과 같이 분쇄하여 가루 형태로 빵류, 면류, 과자류, 스낵류, 수프류 등 다양한 가공제품에 활용할 수 있다(12). 곡류는 발아에 의해 종실에 생화학적, 이화학적 변화를 초래하는데, 발아 시에 가수분해 효소가 활성화되고 이는 전분, 비전분 다당류, 단백질을 분해함으로
써 조직이 연화되어 식감이 개선되며, 종실 내 각종 효소가 활성화되어 생체 기능성 물질의 함량이 증가하는 것으로 (13) 알려져 있다. 또한 발아가 진행되면서 단백질의 소화율 이 높아지고 무기질의 생체 이용률이 증진되며 대사과정에 서 2차 대사산물에 의한 항산화 활성의 증가 등 생리활성을 기대할 수 있다(14). 본 연구에서는 화이트 퀴노아를 이용하 여 발아처리 시에 퀴노아의 이화학적 특성,
in vitro
전분 소화율 및 항산화 활성에 미치는 영향을 조사하고자 하였다.재료 및 방법
재료
본 실험에 사용한 퀴노아는 페루산 화이트 퀴노아(white quinoa)이며 시중에서 구입하여 4°C에 보관하면서 실험에 사용하였다.
발아
정선된 퀴노아를 상온(25°C)에서 3시간 수침한 후 25°C, 습도 70∼95%의 조건으로 식물생장상(Hanbaek Science, Bucheon, Korea)에서 2일간 발아시켰다. 발아과정 중의 퀴 노아를 동결건조기(FD-5508, Ilshinlab, Yangju, Korea)를 사용하여 48시간 동결건조 시켰으며 발아된 싹 부분은 제거 한 후 Cyclotec 1093 sample mill(Tecator, Hoganas, Swe- den)을 사용하여 분쇄한 뒤 분석을 위한 시료로 사용하였다.
일반성분 분석
퀴노아 시료의 수분, 조단백, 조지방, 조회분, 전분 함량은 각각 AACC 방법(15) 44-16, 46-13, 30-10, 08-01, 76- 11에 의해 분석하였다.
총 폴리페놀 및 플라보노이드 함량
퀴노아 시료에 시료 중량의 10배에 해당하는 증류수 또는 80%(v/v) 에탄올을 용매로 사용하여 상온에서 2시간 추출 한 다음 원심분리(15,000 rpm, 30분) 하였다. 퀴노아 추출 물을 사용하여 총 폴리페놀 함량의 분석은 Folin-Denis(16) 방법에 준하여 실험하였으며, 총 플라보노이드 함량은 Zhishen 등(17)의 방법에 따라 측정하였다. 표준물질로 각 각 gallic acid(Sigma-Aldrich Co., St. Louis, MO, USA)와 quercetin(Sigma-Aldrich Co.)을 0~200 ppm 농도로 조 제하여 작성한 검량선에 의해 총 폴리페놀과 총 플라보노이 드 함량을 구하였다.
α-Amylase 활성
퀴노아의 α-amylase 활성은 α-amylase assay kit(Mega- zyme, Wicklow, Ireland)을 사용하여 McCleary와 Shee- han의 방법(18)으로 측정하였다. 퀴노아 시료 0.3 g을 100 mL 정용플라스크에 넣고 1% sodium chloride+0.02%
calcium chloride+0.02% sodium azide 용액으로 정용하
였으며 20°C에서 15분 동안 효소를 추출한 후 원심분리 (1,000×g, 10분) 하였다. BPNPG7(blocked
p
-nitro- phenyl maltoheptaoside)을 포함하는 α-amylase 기질용액 0.2 mL를 테스트 튜브에 넣고 희석된 효소 추출물 0.2 mL를 첨가하여 40°C에서 10분간 반응시켰으며 반응 후 생성된p
-nitrophenol에 의한 용액의 흡광도를 400 nm에서 측정 하였다. 1 Unit/g의 효소 활성은 1분 동안 BPGPN7으로부 터p
-nitrophenol 1 micromole을 생성하는 데 필요로 하는 효소의 양으로 정의되며 Ceralpha Unit으로 표시되었다.Rapid Visco-Analyzer 측정
퀴노아의 호화양상을 신속점도측정계(Rapid Visco Ana- lyzer, Newport Scientific, Sydney, Australia)를 사용하 여 점도 변화로 측정하였다. 즉, 퀴노아 가루 3.5 g(14%
m.b.)을 증류수에 분산시켜 25 mL로 조제한 시료를 RVA cup에 넣고 50°C에서 1분간 유지한 후 분당 12.16°C 속도 로 95°C까지 증가시켰으며 95°C에서 2.5분간 유지한 후 다시 11.84°C의 속도로 50°C로 냉각시켜 측정하였다. 이로 부터 호화개시온도(initial pasting temperature), 최고점도 (peak viscosity), 최저점도(trough, 95°C에서 2.5분 후의 점도), 50°C로 냉각 후의 최종점도(final viscosity)를 측정 하였다.
α-Amylase
in vitro전분 소화율 측정
퀴노아 시료의
in vitro
α-amylase 전분 소화율은 Xue 등 (19)의 방법을 변형하여 측정하였다. 시료 0.5 g에 0.04%(w/v) NaCl을 포함하는 0.05 M sodium phosphate buf- fer(pH 6.9) 용액 50 mL를 넣고 37°C 항온수조에 넣어 10 분간 유지했으며 이 용액에 0.2 mL α-amylase(504 U/mL) 를 넣어 37°C에서 반응시켰다. α-Amylase 효소액은 por- cine pancreatic α-amylase(Sigma-Aldrich Co.)로부터 조 제하였다. 효소반응 중 30, 60분의 간격으로 0.2 mL 용액을 취하여 생성된 환원당을 3,5-dinitrosalicylic acid 시약을 사용한 비색법으로 흡광도를 측정하였다. 표준당으로 mal- tose를 사용하였으며 전분의 가수분해율(%)은 [standard curve로부터 환산된 maltose 함량(mg)/ 전분 함량(mg)]×
100으로 계산하였다.
퀴노아 추출물의 DPPH 소거능
퀴노아 시료의 항산화 활성은 Blois(20)의 방법을 변형한 DPPH(1,1-diphenyl-2-picrylhydrazyl) 자유라디칼 소거 능 측정방법에 의해 평가하였다. 퀴노아 추출물 0.2 mL에 1×10-4 M DPPH 용액(에탄올에 용해) 0.8 mL를 넣고 10초 간 진탕 후 실온에서 정확히 10분 동안 방치한 다음 525 nm에서 흡광도를 측정하였다. 바탕시험인 시료 무첨가구는 시료 대신 증류수를 사용하여 측정하였다. DPPH 자유라디 칼 소거능은 [1-(시료첨가구의 흡광도/무첨가구의 흡광 도)]×100으로 나타내었다.
Table 1. Proximate composition1) of white quinoa during germination (%) Moisture Crude protein2) Crude fat Crude ash Total carbohydrate3) Raw quinoa
Germination (24 h) Germination (48 h)
7.02±0.96a4) 2.23±0.50b 2.28±0.34b
14.98±0.36a 12.20±0.21c 13.27±0.42b
6.16±0.08a 5.19±0.17b 5.28±0.08b
2.38±0.02a 1.75±0.04c 1.86±0.06b
76.48±0.20c 80.86±0.21a 79.60±0.37b
1)Dry weight basis except moisture.
2)Nitrogen×6.25.
3)% Carbohydrate=100%-(% crude protein+% crude fat+% crude ash).
4)Values are means±SD of triplicate determinations. Values with different letters in a column are significantly different by Duncan’s multiple range test (P<0.05).
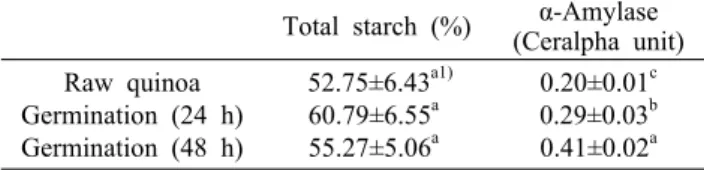
Table 2. Total starch content and α-amylase activity of white quinoa during germination
Total starch (%) α-Amylase (Ceralpha unit) Raw quinoa
Germination (24 h) Germination (48 h)
52.75±6.43a1) 60.79±6.55a 55.27±5.06a
0.20±0.01c 0.29±0.03b 0.41±0.02a
1)Values are means±SD of triplicate determinations. Values with different letters in a column are significantly different by Dun- can’s multiple range test (P<0.05).
통계처리
통계처리는 SAS program(SAS Institute, Cary, NC, USA)을 이용하여 평균과 표준편차(mean±SD)로 제시하였 고 Duncan’s multiple range test를 이용하여 유의적인 차 이를 분석하였다.
결과 및 고찰
발아 퀴노아의 일반성분
화이트 퀴노아의 발아에 따른 일반성분의 변화를 분석한 결과는 Table 1과 같다. 퀴노아 원곡은 조단백질 14.98%, 조지방 6.16%, 조회분 2.38%, 총탄수화물 76.48%로 분석 되었다. 이는 퀴노아의 일반성분이 조단백질 9.1~16.7%, 조지방 4.0~8.5%, 조회분 1.0~3.8% 범위로 보고한 결과 (1,5)의 범위 내에 있었다. 퀴노아를 24시간 발아시켰을 때 퀴노아의 조단백질, 조지방, 조회분 함량은 각각 12.20%, 5.19%, 1.75%로 감소하는 것으로 나타났다. 48시간 발아 시에는 각각 13.27%, 5.28%, 1.86%로 나타나 2일간의 발 아기간 중에 조단백질, 조지방, 조회분 함량은 감소하였으 며, 이에 따라 총탄수화물 함량은 증가한 것으로 분석되었 다. 발아시간에 따라서는 발아 24시간 비해 발아 48시간에 서 조단백과 조지방의 함량이 다소 미미하게 증가하는 경향 을 보여주었다. 이는 퀴노아가 발아 초기에 단백질과 지질이 발아 중에 생합성된 분해효소들에 의해 먼저 분해되어 사용 되기(21,22) 때문으로 판단되었다.
발아 퀴노아의 전분 함량 및 α-amylase 활성
퀴노아 시료의 발아에 따른 전분 함량과 α-amylase 활성 을 측정한 결과는 Table 2와 같다. 퀴노아 원곡의 전분 함량 은 52.75%로 퀴노아의 주성분인 전분 함량을 30~70% 사
이로 보고한(23) 범위 안에 있었다. 본 실험에서 퀴노아는 발아 1~2일 동안에 전분 함량이 약간 높게 나타났는데 이는 발아 초기에 단백질, 지방, 무기질 등 일반성분의 함량 감소 가 전분보다 상대적으로 더 컸기 때문으로 판단되었으며, 발아 중 전분 함량의 감소는 발아 36시간 이후부터 관찰되 었다는 보고(24)와 유사하였다. 퀴노아는 발아 전에 α- amylase의 활성이 미미하였으나 2일간 발아가 진행됨에 따 라 증가하는 추세를 나타냈다. Hager 등(24)은 퀴노아를 침 지한 후 발아 시에 α-amylase의 활성이 72시간에서 가장 높게 나타났으며, 배아를 분리한 뒤 α-amylase 활성을 측정 한 결과에서는 침지 후 12∼48시간에서 최대인 이후로 급격 히 감소함에 따라 α-amylase가 배아에서 합성되어 외배유 로 이동한다고 보고하였다. 또한 퀴노아는 발아 중 α-amy- lase의 활성이 25°C 발아온도에서 발아 36시간까지 증가하 였으나 그 이후에는 감소한다고 하였다(25).
발아 퀴노아의 총 페놀 및 총 플라보노이드 함량
발아에 따른 퀴노아의 총 페놀 및 총 플라보노이드 함량을 분석한 결과는 Table 3에 나타나 있다. 증류수와 80% 에탄 올을 용매로 하여 추출한 퀴노아 원곡의 총 페놀 함량은 각 각 1.37 mg/g, 1.08 mg/g이었으며, 이는 퀴노아에 포함된 폴리페놀 함량을 0.4∼8.64 mg/g으로 보고한 결과(26-28) 의 범위 내에 있었다. 증류수를 용매로 하여 추출한 퀴노아 의 총 페놀 함량은 발아 24시간 후에 1.22 mg/g으로 다소 감소하다가 발아 48시간 후에 1.42 mg/g으로 증가하는 양 상을 보였다. 퀴노아를 에탄올 용매로 추출하여 총 페놀 함 량을 측정하였을 때 발아 24시간, 발아 48시간은 각각 1.24, 1.33 mg/g으로 계속 증가하였다. 이는 퀴노아 메탄올 추출 물의 총 페놀 함량이 sprouting에 의해 증가하는 경향이 있 다는 보고(26)와 유사하게 나타났다.
퀴노아 원곡의 총 플라보노이드 함량은 증류수와 80%
에탄올 추출물에서 각각 0.76 mg/g, 0.35 mg/g이었으며 퀴노아의 플라보노이드 함량은 약 0.1∼1.9 mg/g의 범위로 보고된(11,27,28) 바 있다. 발아 퀴노아의 총 플라보노이드 함량은 증류수 추출 시 발아 24시간에 0.57 mg/g, 발아 48 시간에 0.61 mg/g으로 원곡보다 약간 감소한 값을 주었다.
에탄올 추출 시에도 원곡에 비해 발아 24시간, 48시간에 각각 0.16, 0.13 mg/g으로 증류수 추출보다 낮은 값을 보이 며 전반적으로 감소하는 추세를 보여주었다. Carciochi 등
Table 3. Total phenolic and flavonoid contents1) of white quinoa during germination (mg/g)
Total phenolic Total flavonoid
Water 80% ethanol Water 80% ethanol
Raw quinoa Germination (24 h) Germination (48 h)
1.37±0.09a2) 1.22±0.02b 1.42±0.01a
1.08±0.26a 1.24±0.28a 1.33±0.20a
0.76±0.14a 0.57±0.09b 0.61±0.07ab
0.35±0.08a 0.16±0.06bc 0.13±0.07c
1)Dry weight basis.
2)Values are means±SD of triplicate determinations. Values with different letters in a column are significantly different by Duncan’s multiple range test (P<0.05).
Table 4. Rapid Visco-Analyzer (RVA) pasting properties of white quinoa during germination Pasting temp.
(°C)
Viscosity (RVU)1)
Peak Trough Break down Final Setback Raw quinoa
Germination (24 h) Germination (48 h)
69.83±0.42a2) 69.88±1.28a 70.13±6.48a
141.17±2.00a 130.82±0.42a 82.71±10.21b
129.42±1.50a 117.67±0.59a 76.13±11.80b
11.75±0.50a 13.17±0.00a 6.59±1.59b
179.88±3.46a 173.84±0.58a 116.00±13.92b
50.46±1.96a 56.17±0.00a 39.88±2.13b
1)Trough=minimum viscosity after the peak, breakdown=peak viscosity minus trough viscosity, setback=final viscosity minus peak viscosity.
2)Values are means±SD of triplicate determinations. Values with different letters in a column are significantly different by Duncan’s multiple range test (P<0.05).
Table 5. Relative starch digestibility1) in vitro of white quinoa during germination
Hydrolysis time (min)
30 60
Raw quinoa Germination (24 h) Germination (48 h)
31.68±2.47a2) 31.25±2.00a 35.80±2.10a
59.87±2.09b 58.30±1.60b 65.66±0.97a
1)Measured as starch degree of hydrolysis (%) with α-amylase after 30 and 60 min of incubation at 37°C.
2)Values with different letters in a column are significantly dif- ferent by Duncan’s multiple range test (P<0.05).
(27)은 퀴노아의 총 페놀 함량과 총 플라보노이드 함량이 3일간의 발아기간 중 유의적으로 증가한다고 보고하여 본 실험의 결과와는 차이를 보여주었다. 한편 발아현미의 경우 발아에 따라 폴리페놀 함량은 약간 증가하나 플라보노이드 함량은 감소한다고 보고된(29) 바 있다. Kim 등(30)에 의하 면 총 7종의 현미를 발아 전후로 생리활성물질을 비교하였 을 때 대부분 발아 후 총 플라보노이드 함량이 감소한다고 보고하였으며, 이는 재배 품종, 생육하는 환경 조건 및 성숙 도, 수확 후 관리 등의 차이가 영향을 미치는 것으로 설명한 바 있다. 본 실험에서 발아 퀴노아의 총 플라보노이드 함량 이 원곡에 비해 낮게 나타난 이유는 발아 중 퀴노아를 동결 건조 하여 싹을 제거한 다음 분석에 사용했기 때문으로 판단 되며, 발아 후 퀴노아의 부위에 따라 총 페놀과 플라보노이 드 함량에 차이가 있기 때문으로 사료되었다.
발아 퀴노아의 RVA 호화 특성
퀴노아 원곡과 발아에 따른 퀴노아 가루의 호화양상을 신 속점도측정기(RVA)로 측정한 결과는 Table 4에 나타나 있 다. 퀴노아의 호화개시온도는 발아가 1~2일 진행되면서 큰 차이가 없었다. 퀴노아 가루의 RVA 점도는 발아가 진행되 면서 감소하는 추세를 나타내었으며, 특히 발아 48시간에 발아 퀴노아 가루의 peak, trough, final 점도가 현저하게 감소하였다. 노화의 정도를 나타내는 setback 점도 역시 발 아가 진행함에 따라 감소하였다. 발아에 따라 RVA 점도가 감소하는 이유는 발아 시에 생성되고 활성화된 전분 분해효 소에 의하여 전곡립 내에 전분이 다소 분해되면서 호화에 따른 페이스트 점도를 감소시키기 때문으로 판단되었다. 이 에 따라 발아에 따른 퀴노아의 α-amylase 효소 활성도 증가 가 RVA 페이스트 점도를 감소시키는 주요 요인임을 확인할 수 있었다.
발아 퀴노아의
in vitro전분 소화율
발아 퀴노아의 전분 소화율을 pancreatic α-amylase를 이용하여
in vitro
가수분해에 따른 전분 가수분해율(%)로 측정한 결과는 Table 5와 같다. 전분의in vitro
가수분해율 에서 차이는 전분의 출처, 입자크기, amylose/amylopectin 비율, 결정성 정도 및 형태, amylose-lipid 복합체, 식이섬 유와 관련한 전분의 분포, 항영양소, α-amylase 저해제, 가 공조건 등 다양한 요인에 의해 영향을 받으며 인체의 식후 혈당 반응과 관계가 있어(31) 건강과 영양적인 관점에서 중 요하다. 퀴노아 원곡과 발아 24시간까지는 30분 경과 시의 전분 가수분해율이 각각 31.68%, 31.25%였으며, 60분이 경과되었을 때는 59.87%, 58.30%로 나타나 전분 가수분해 율에 큰 차이가 없는 것으로 나타났다. 하지만 발아 48시간 퀴노아 시료에서는 전분 가수분해율이 30분 후 35.80%, 60 분 후 65.66%로 나타나 다소 증가하였다. 이는 발아 퀴노아 의 α-amylase 활성 분석 결과와 연관하여 발아 시에 생성된 효소 활성이 전분의 가수분해율에 영향을 주었기 때문으로 판단되었다.Table 6. DPPH radical scavenging activity of white quinoa dur- ing germination (%) Water extract 80% ethanol extract Raw quinoa
Germination (24 h) Germination (48 h)
21.68±5.05ab1) 16.15±3.82bc 12.38±3.36c
20.02±3.68a 19.47±2.47a 16.53±2.27a
1)Values are means±SD of triplicate determinations. Values with different letters in a column are significantly different by Dun- can’s multiple range test (P<0.05).
발아 퀴노아의 DPPH 라디칼 소거능
발아 퀴노아 시료를 증류수와 80% 에탄올 두 가지 용매 를 사용하여 추출한 뒤 추출물의 DPPH 자유라디칼 소거능 을 측정한 결과는 Table 6과 같다. 증류수로 추출하였을 때 원곡, 발아 24시간 및 48시간에서 각각 21.68%, 16.15%, 12.38%로 나타나 발아 퀴노아가 퀴노아 원곡보다 DPPH 라 디칼 소거능이 감소함을 나타냈다. 또한, 에탄올 추출 시에 도 원곡, 발아 24시간 및 48시간에서 각각 20.02%, 19.47%, 16.53%로 나타나 발아 퀴노아 추출액에서 DPPH 라디칼 소 거능이 더 낮은 것으로 나오지만 증류수를 이용하여 추출하 였을 때보다는 감소율이 낮은 것을 알 수 있다. 그러나 퀴노 아의 DPPH 소거능은 3일간의 발아기간 중에 증가하는 것 으로 보고된(27) 바 있다. 한편 Kim 등(32)에 의하면 DPPH 라디칼 소거능은 벼 에탄올 추출물에서 발아 전 22.95%가 발아 후 31.32 %로 증가하였지만 왕겨와 현미 부분에서는 발아 후 감소하였으며, 총 항산화력은 에탄올 추출물에서 발아 전과 후에 큰 차이를 보이지 않았으나 싹 부분에서는 높게 나타났다고 보고한 바 있다. 본 실험에 사용한 화이트 퀴노아 시료는 발아 중 싹 부분을 제거하는 과정을 거친 후 분석에 사용하였기 때문에 DPPH 라디칼 소거능에서 그 차 이를 보이는 것으로 사료되었다.
요 약
화이트 퀴노아(white quinoa)의 발아처리가 퀴노아의 일반 성분, 총 페놀 및 플라보노이드 함량, α-amylase 활성, RVA 호화양상 등 이화학적 특성과
in vitro
전분 소화율 및 항산 화 활성에 미치는 영향을 조사하였다. 퀴노아는 2일간의 발 아기간 중에 일반성분(조단백질, 조지방, 조회분 함량)은 감 소하였으며 전분을 포함한 총탄수화물 함량은 다소 증가하 였다. 발아 퀴노아의 호화 특성을 신속점도측정계(RVA)로 측정한 결과 RVA 점도는 2일간 발아에 따라 점도가 점차 감소하였으며, 발아에 따른 amylase 효소 활성으로 인한 전분의 분해가 점도에 영향을 미치는 것으로 나타났다.퀴노 아의in vitro
전분 가수분해율은 발아에 따라 증가하는 경향 을 보여주었다. 퀴노아의 총 페놀 함량은 80% 에탄올 추출 시 발아 2일까지 약간 증가하였으나 총 플라보노이드 함량 과 DPPH 라디칼 소거능에 의한 항산화 활성은 감소하는 경 향을 보여주었다. 본 연구를 통해 발아 퀴노아는 부위에 따라서 기능성 성분 함량 및 항산화 활성 등 생리활성에 차이 가 있음을 제시해 주었다.
REFERENCES
1. Vega-Galvez A, Miranda M, Vergara J, Uribe E, Puente L, Martinez EA. 2010. Nutrition facts and functional potential of quinoa (Chenopodium quinoa willd.), an ancient Andean grain: a review. J Sci Food Agric 90: 2541-2547.
2. Belton PS, Taylor JRN. 2002. Pseudocereals and less com- mon cereals. Springer-Verlag Berlin Heidelberg, Berlin, Germany. p 93-122.
3. Abugoch James LE. 2009. Quinoa (Chenopodium quinoa Willd.): composition, chemistry, nutritional, and functional properties. Adv Food Nutr Res 58: 1-31.
4. Lee JH. 2007. New beneficial crops amaranth and quinoa for food nutritional source. Food Industry and Nutrition 12 (2): 29-36.
5. Nowak V, Du J, Charrondiere UR. 2016. Assessment of the nutritional composition of quinoa (Chenopodium quinoa Willd.). Food Chem 193: 47-54.
6. Ando H, Chen YC, Tang H, Shimizu M, Watanabe K, Mit- sunaga T. 2002. Food components in fractions of quinoa seed. Food Sci Technol Res 8: 80-84.
7. Prego I, Maldonado S, Otegui M. 1998. Seed structure and localization of reserves in Chenopodium quinoa. Ann Bot 82: 481-488.
8. Alvarez-Jubete L, Arendt EK, Gallagher E. 2010. Nutritive value of pseudocereals and their increasing use as functional gluten-free ingredients. Trends Food Sci Technol 21: 106- 113.
9. Koziol MJ. 1992. Chemical composition and nutritional evaluation of quinoa (Chenopodium quinoa Willd.). J Food Compos Anal 5: 35-68.
10. Navruz-Varli S, Sanlier N. 2016. Nutritional and health ben- efits of quinoa (Chenopodium quinoa Willd.). J Cereal Sci 69: 371-376.
11. Hirose Y, Fujita T, Ishii T, Ueno N. 2010. Antioxidative properties and flavonoid composition of Chenopodium qui- noa seeds cultivated in Japan. Food Chem 119: 1300-1306.
12. Goh HK, Lee YT. 2017. Effects of heat treatments on phys- icochemical properties and in vitro biological activities of quinoa (Chenopodium quinoa Willd.). J Korean Soc Food Sci Nutr 46: 688-694.
13. Hubner F, Arendt EK. 2013. Germination of cereal grains as a way to improve the nutritional value: a review. Crit Rev Food Sci Nutr 53: 853-861.
14. Laus MN, Cataldi MP, Robbe C, D’Ambrosio T, Amodio ML, Colelli G, De Santis G, Flagella Z, Pastore D. 2017.
Antioxidant capacity, phenolic and vitamin C contents of quinoa (Chenopodium quinoa Willd.) as affected by sprout- ing and storage conditions. Ital J Agron 12: 63-68.
15. AACC. 2000. Approved methods of the AACC. 10th ed.
American Association of Cereal Chemists, St. Paul, MN, USA.
16. Folin O, Denis W. 1912. On phosphotungstic-phosphomo- lybdic compounds as color reagents. J Biol Chem 12: 239- 243.
17. Zhishen J, Mengcheng T, Jianming W. 1999. The determi- nation of flavonoid contents in mulberry and their scaveng- ing effects on superoxide radicals. Food Chem 64: 555-559.
18. McCleary BV, Sheehan H. 1987. Measurement of cereal α-amylase: A new assay procedure. J Cereal Sci 6: 237-251.
19. Xue Q, Newman RK, Newman CW. 1996. Effects of heat treatment of barley starches on in vitro digestibility and glu- cose responses in rats. Cereal Chem 73: 588-592.
20. Blois MS. 1958. Antioxidant determinations by the use of a stable free radical. Nature 181: 1199-1200.
21. Makinen OE, Hager AS, Arendt EK. 2014. Localisation and development of proteolytic activities in quinoa (Chenopo- dium quinoa) seeds during germination and early seedling growth. J Cereal Sci 60: 484-489.
22. Park SH, Morita N. 2004. Changes of bound lipids and com- position of fatty acids in germination of quinoa seeds. Food Sci Technol Res 10: 303-306.
23. Li G, Zhu F. 2018. Quinoa starch: structure, properties, and applications. Carbohydr Polym 181: 851-861.
24. Hager AS, Makinen OE, Arendt EK. 2014. Amylolytic activ- ities and starch reserve mobilization during the germination of quinoa. Eur Food Res Technol 239: 621-627.
25. Rosa M, Hilal M, González JA, Prado FE. 2004. Changes in soluble carbohydrates and related enzymes induced by low temperature during early developmental stages of qui- noa (Chenopodium quinoa) seedlings. J Plant Physiol 161:
683-689.
26. Alvarez-Jubete L, Wijngaard H, Arendt EK, Gallagher E.
2010. Polyphenol composition and in vitro antioxidant ac- tivity of amaranth, quinoa buckwheat and wheat as affected
by sprouting and baking. Food Chem 119: 770-778.
27. Carciochi RA, Manrique GD, Dimitrov K. 2014. Changes in phenolic composition and antioxidant activity during ger- mination of quinoa seeds (Chenopodium quinoa Willd.). Int Food Res J 21: 767-773.
28. Hemalatha P, Bomzan DP, Rao BVS, Sreerama YN. 2016.
Distribution of phenolic antioxidants in whole and milled fractions of quinoa and their inhibitory effects on α-amylase and α-glucosidase activities. Food Chem 199: 330-338.
29. Sung J, Lee J, Oh SK, Lee JS, Choi WS. 2013. Changes in phytochemical content and antiproliferative activity of germinated Geunnun and Ilpum rice varieties. J Korean Soc Food Sci Nutr 42: 1157-1161.
30. Kim DJ, Oh SK, Yoon MR, Chun AR, Choi IS, Lee DH, Lee JS, Yu KW, Kim YK. 2011. The change in biological activities of brown rice and germinated brown rice. J Kore- an Soc Food Sci Nutr 40: 781-789.
31. Hoover R, Zhou Y. 2003. In vitro and in vivo hydrolysis of legume starch by α-amylase and resistant starch formation in legumes-a review. Carbohydr Polym 54: 401-417.
32. Kim HY, Hwang IG, Kim TM, Park DS, Kim JH, Kim DJ, Lee J, Jeong HS. 2011. Antioxidant and angiotensin convert- ing enzyme I inhibitory activity on different parts of germi- nated rough rice. J Korean Soc Food Sci Nutr 40: 775-780.