학 술 논 문
138
3T3-L1 지방전구세포의 지방분화에서 멜라토닌의 영향
이정근 · 이영훈 · 김지현*
연세대학교 보건과학대학 의공학부
Effect of Melatonin on Differentiating 3T3-L1 Preadipocytes
Jeongkun Lee, Yeong Hun Lee, and Chi Hyun Kim*
Department of Biomedical Engineering, Yonsei University
(Manuscript received 11 May 2020 ; revised 19 June 2020 ; accepted 22 June 2020)
Abstract: Adipocytes are the main constituent of adipose tissue. Understanding the molecular basis of adipogenesis is pivotal to finding the therapeutic targets for treatment of obesity. Melatonin is associated with obesity and its mech- anism is currently under intensive investigation. The objective of this study was to investigate the effect of melatonin on adipogenesis in differentiating preadipocytes. 3T3-L1 preadipocytes were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 5% calf serum at 37
oC with 5% CO
2in a humidified incubator. Differentiation was induced using DMEM with 10% fetal bovine serum supplemented with MDI two days after cell confluence (day 0).
Cells were treated with 0, 10 and 100 μM melatonin on either day 0 or day 5. 72 hours after each treatment, lipid accumulation was measured by oil red O staining. Proteins were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to membranes. As a result, lipid accumulation decreased with mel- atonin treatment. ERK pathway, activated when differentiation is induced, also decreased with an increase in melatonin concentration. Furthermore, the expression of key adipogenic factors, C/EBP α, C/EBPβ, and PPARγ, were reduced by melatonin treatment. These results imply that melatonin may inhibit the process of adipogenesis and may have a role as a new anti-obesity agent.
Key words: 3T3-L1 preadipocyte, Adipogenesis, Cell death, Differentiation of adipocyte, Melatonin
I. 서 론
비만은 에너지 대사의 불균형으로 인한 지방 조직의 과잉 발달을 특징으로 하는 대사 질환이다. 지방 조직의 과도한 증식은 지방세포의 인슐린 민감성을 감소시키고 혈중 콜레 스테롤 농도를 증가시켜 제 2형 당뇨나 고지혈증, 동맥경화 등 심혈관계 질환 등을 유발하며[1,2] body mass index(BMI)를 기준으로 30 이상일 시 비만으로 정의한다[3]. 비만의 유발은 생활습관과 밀접한 관련이 있다. 정적인 생활과 육류 위주의 식 습관에 의한 에너지의 소비량과 섭취량의 불균형은 널리 알
려진 요인 중의 하나이다[4]. 최근에는 수면 패턴과 비만 유 병률 간의 상관관계를 규명하기 위한 연구들도 진행 중이다. 본 연구들에 의하면 하루 수면 시간이 6시간 이내인 사람의 경우 비만과 그 합병증의 유병률이 높게 나타났다[5,6]. 그러나 수면 부족이 비만을 유발하는 정확한 기전은 아직 불명확하다.
지방생성(adipogenesis)은 지방세포로 분화하기 전 단계인 지방전구세포가 성숙한 지방세포로 분화하는 일련의 과정으로, 과 도한 지방생성은 비만의 주요 특징인 지방 조직의 과잉 발달로 이어진다[7]. 지방생성은 세포 내 다양한 성장 인자들과 사 이토카인(cytokine), 호르몬의 영향을 받으므로 여러 생화 학적 방법을 통하여 지방생성의 기전을 이해하고, 이를 억 제하여 비만 치료에 활용하는 연구가 진행 중이다[8-10]. 지 방생성을 조절하는 것으로 알려진 주요 전사 인자들로 peroxisome proliferator-activated receptor gamma (PPARγ) 와 CCAAT/enhancer-binding proteins (C/EBPs)가 있으 며[7], 이 인자들의 발현 정도를 비교함으로써 지방생성의 Corresponding Author : Chi Hyun Kim
Department of Biomedical Engineering, College of Health Sciences, Yonsei University, Wonju, Gangwon-do, 26493, Korea Tel: +82-33-760-2785
E-mail: [email protected]
이 연구는 교육부의 재원으로 한국연구재단의 지원을 받아 수행된 기
초연구사업임(NRF-2015R1D1A1A01060699).
139 발생 유무를 판단할 수 있다.
또한 지방세포의 수를 줄여 과잉 증식을 예방하기 위한 방법으로서 지방세포의 세포자살(apoptosis)이나 자가포식 작용(autophagy)을 유도할 수 있다[11]. 세포자살은 세포 괴사(necrosis)와는 구분되는 programmed cell death로서, 세포 위축, 세포막 blebbing, 염색질 응축 및 DNA 분절을 통하 여 불필요한 세포 및 조직을 제거하는 세포 기전이며[12], cysteine-aspartic proteases (caspase) family 에 의해 조 절된다. 자가포식작용은 불필요한 세포나 세포소기관을 가 수분해하고 그 잔해를 재활용하는 기전으로, mechanistic target of rapamycin (mTOR) signaling 을 통해 생성된 자가포식소체(autophagosome)의 격리 작용에 의해 이루어지 며 Beclin, LC3 등의 marker를 통해 발생 여부를 파악할 수 있다[13].
멜라토닌(melatonin)은 송과선(pineal gland)에서 분비 되어 생물의 일주기를 조절하는 호르몬으로, 낮에 합성되고 밤에 분비되어 수면을 유도한다[14]. 생물의 일주기와 여러 세포 및 조직들의 상관관계를 규명하기 위하여 in vitro 환 경에서 멜라토닌을 이용한 실험이 진행 중이다. 지방세포에 관한 연구로서, 3T3-L1 지방전구세포에 멜라토닌을 다양한 농도와 시간 조건으로 투여하여 세포의 증식률과 지방생성 정도를 분석한 실험이 존재한다[15,16]. 해당 연구결과에 따 르면 멜라토닌의 투여 농도 및 시간에 따라 지방전구세포의 증식 및 지방생성이 촉진되었으며, 지방세포의 생존에는 크게 기여하지 않았다. 그러나 이 연구들은 각각 3T3-L1을 분화 시키지 않거나 혹은 분화를 마친 상태에서 실험을 진행하였 으며, 지방세포의 분화 단계에 따른 멜라토닌의 영향을 분 석한 연구는 미비한 실정이다. 그 예로, 지방세포로 분화가 가능한 간엽줄기세포(mesenchymal stem cell)에 멜라토닌을 투여했을 시에는 위의 결과와 달리 지방생성이 억제되는 결 과가 나타났으며[17], 이는 멜라토닌이 지방세포에 주는 영 향은 그 농도, 처리 시간뿐만 아니라 세포의 분화 단계와 관련이 있다는 것을 암시한다. 따라서 멜라토닌과 지방세포 간의 뚜 렷한 상관관계를 규명하기 위하여 보다 다양한 측면의 연구가 이루어져야 한다.
우리는 일주기 조절 호르몬인 멜라토닌과 지방세포의 분화 기전 간의 상관관계를 분석하기 위하여, 분화 중인 3T3-L1 지방전구세포에 멜라토닌을 0, 10, 100 µM의 농도로 처리 하여 실험을 진행하였다. 분화 과정 중 멜라토닌 투여 기간은 induction media 로 배양하여 분화를 유도하는 분화 초기의 induction 단계와, growing media로 배양하여 세포를 성 숙시키는 분화 후기의 growing 단계로 정하였으며[7], 총 3 일 동안 멜라토닌의 영향을 받도록 하였다. 그 후 지방 과 립의 생성과 oil red O staining, 지방생성, 세포자살, 자 가포식작용 인자들을 분석하였다.
II. 연구 방법
1. 세포 분화 및 멜라토닌 처리
세포는 mouse-derived 3T3-L1 지방전구세포(Korean Cell Line Bank, Korea) 를 사용하였으며, 계대 배양은 1 주일에 한 번, confluence가 80% 이상 도달하였을 때 실 시하였다. 사용한 배지는 Dulbecco’s modified eagle’s medium (DMEM, Lonza, USA) 에 calf serum (CS, Gibco, USA) 5%, penicillin/streptomycin (Lonza) 1% 를 혼합하여 사용하였 다. 세포 배양에 사용된 모든 용액은 사용 전 water bath를 이 용하여 37
oC 로 맞춰 사용하였다. 세포 배양기는 온도 37
oC 에 이산화탄소 농도 5%로 고정하였다. 지방세포 분화는 6-well plate 에 세포를 100% confluence에 도달하도록 배양하여 실시하였다. Induction 단계인 처음 3일 동안 fetal bovine serum (FBS, Lonza) 이 10% 혼합된 DMEM (10% FBS/
DMEM) 에 1 µM dexamethasone (Dex, Sigma, St. Louis, MO, USA), 0.5 mm 3-isobutyl-1-methylxanthine (IBMX, Sigma), 그리고 10 µM insulin (Sigma)을 혼합하여 세포를 배 양하였다. 그 후 배지를 10% FBS/DMEM에 10 µM insulin을 혼합한 배지로 교체하여 2일 간 배양하였다. Growing 단 계인 마지막 3일 동안 배지를 10% FBS/DMEM으로 교체 하여 배양하였다. 멜라토닌은 dimethyl sulphoxide (DMSO, Sigma)에 희석하여, 배지에 투여하였을 때 최종 농도가 0, 10, 100 µM 가 되도록 하였다[15,16]. 그 후 induction 과 growing 단계에 각각 3일 동안 처리하였다[7].
2. Oil red O staining
배지를 제거하고 세포를 PBS로 세척한 뒤, 10% formalin으 로 실온에서 각각 10 분, 1 시간으로 총 두 번 배양하여 고정 하였다. 그 후 formalin을 PBS로 세척하여 완전히 제거한 뒤, 60% iso-propanol (Sigma) 로 2분 간 세척하였다. Oil red O (Sigma) 는 stock solution (0.35g oil red O in 100 ml isopropanol) 을 증류수에 6:4의 비율로 희석하여 사용하였 으며, 실온에서 세포에 10분 간 처리하여 염색하였다. 염색 한 후 증류수로 4번 세척한 뒤 현미경으로 관찰하였다.
3. Western Blot
분화가 모두 종료된 8일차 때, induction 단계와 growing 단계의 세포를 각각 수확하여 단백질을 추출한 뒤, BCA assay kit 을 이용하여 단백질 농도를 측정하였다. SDS-PAGE 는 acrylamide/bis의 농도를 running gel 9%, stacking gel 5%
로 설정하였으며, 80V의 전압으로 2 시간 동안 전기영동을
실시하였다. 전기영동 후, 100V의 전압에서 50분 동안 단
백질을 PVDF membrane으로 transfer한 후 shaker를 이
용, membrane을 1 시간 동안 5% skim milk로 blocking
140
하였다. 그 후 membrane을 1차 항체에 하룻밤 동안, 2차 항체에 1 시간 동안 배양하였다. 1차 항체는 BSA에 1:1000의 비율로 희석하여 사용하였으며, p-ERK (sc-7383, Santa Cruz, USA), ERK (sc-93, Santa Cruz), C/EBP α (#2295, Cell Signaling, USA), C/EBP β (#3087, Cell Signaling), PPARγ (#2430, Cell Signaling), Caspase-3 (#9662, Cell Signaling), LC3 (#4108, Cell Signaling), Beclin (#3738, Cell Signaling), GAPDH (sc-25778, Santa Cruz) 등이 실험에 사용되었다. 2차 항체는 5% skim milk에 1:5000의 비율로 희석하여 사용하였다.
처리의 각 과정 사이에는 TBS-T용액을 사용하여 10분씩 네 번, 총 40분 간 membrane을 shaker로 세척하였다.
4. 통계 분석
감광하여 필름에 검출된 밴드는 Image J 프로그램을 이 용하여 밴드의 밝기를 측정 후, Graphpad Prism 5 프로 그램을 이용하여 그래프를 산출하였고, 같은 프로그램을 이 용하여 one-way ANOVA를 수행하였다. 결과 값은 평균 ±
표준편차(SD)로 표현하였고, 사후 검정은 Tukey’s test를 사용하였다. 유의 수준은 0.05로 설정하였다.
III. 연구 결과
먼저 oil red O staining을 이용하여 지방전구세포의 분화 과 정 중에 멜라토닌을 처리했을 때 지방 과립의 변화를 농도 별로 분석하였다(그림 1). Induction 단계의 세포에서는 지 방 과립이 발달하지 않아 큰 차이가 관찰되지 않았다. 반면 에, growing 단계의 세포는 멜라토닌을 투여한 실험군에서 지방 과립에 의한 염색이 대조군에 비해 면적이 적게 나타 남을 확인할 수 있었다. 또한 이 결과는 멜라토닌의 농도가 높을수록 두드러져, 100 µM 농도에서 대조군과 비교했을 때 약 20% 감소하여 가장 뚜렷한 변화를 보였다(p<0.01).
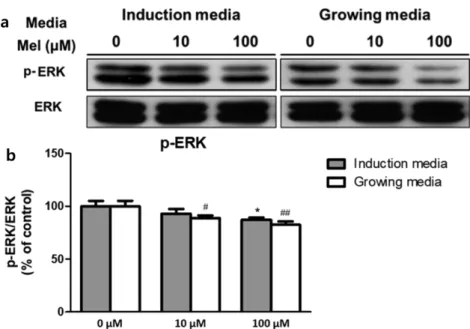
지방전구세포에 대한 멜라토닌의 영향이 ERK pathway를 경유하는지 알아보기 위해, ERK의 인산화 정도를 western blot으로 분석하였다(그림 2). 그 결과 두 단계 모두 ERK의 인
그림 1. 멜라토닌을 처리한 각 3T3-L1 실험군에 oil red O staining을 실시한 결과. (a) 광학현미경을 통해 ×200 배율로 확대한 사진 (scale bar = 100 µm), (b) 각 실험군의 지방과립 생성의 변화. (n=3, **P<0.01, vs control of growing media)
Fig. 1. The result of oil red O stained on each 3T3-L1 groups treated with melatonin. (a) Microscopy bright field images using a microscope with a magnification ×200 (scale bar = 100 µm), (b) lipid accumulation of growing media group. (n=3,
**P<0.01, vs control of growing media)
산화 형태인 p-ERK의 발현이 멜라토닌 농도에 따라 감소 141 하였으며, 100 µM에서 가장 큰 감소를 보였다. 분화 초기 인 induction media 처리 단계에서는 p-ERK의 발현이 대 조군과 비교했을 때 약 10% (p<0.05), 분화 후기인 growing media 단계에서는 약 20% (p<0.01) 감소하였다. 특히 이 결과는 분화 초기 단계보다 분화 후기 단계에서 두드러지게 나타났는데, 분화 초기에 멜라토닌을 10 µM 투여한 실험 군에서는 대조군과 뚜렷한 차이가 드러나지 않은 반면에, 분 화 후기에서는 10 µM 실험군에서도 p-ERK의 발현이 대 조군과 비교했을 때 약 10% 감소하였다(p<0.05).
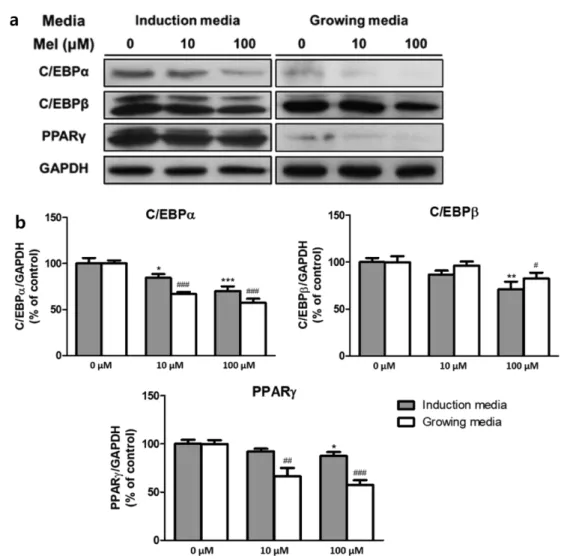
분화 중인 지방전구세포의 지방 생성 효과가 멜라토닌에 의해 촉진되었는지 혹은 억제되었는지 확인하기 위하여 C/
EBPα, C/EBPβ, 그리고 PPARγ 등 주요 지방 생성 인자들의 발 현 정도를 western blot으로 분석하였다(그림 3). 그 결과 세 인 자 모두 두 분화 단계에서 100 µM의 멜라토닌을 처리하였을 때 발현이 감소하였다. C/EBPα의 경우 가장 크게 감소하였는 데, 100 µM의 멜라토닌을 처리하였을 때 분화 초기 induction media 처리 단계에서는 대조군에 비해 약 30%(p<0.001), 분화 후기 growing media 처리 단계에서는 약 40%(p<0.001) 감소하였다. 또한 ERK pathway의 결과와 유사하게, induction 단계보다 growing 단계에서 C/EBPα와 PPARγ의 발현 감 소 효과가 더 크게 나타났으며 10 µM의 농도에서도 발현이 감 소였다. 대조군과 비교했을 때 10 µM의 농도에서 C/EBPα의 경우 약 30% (p<0.001), PPARγ는 약 30% (p<0.01)로 발
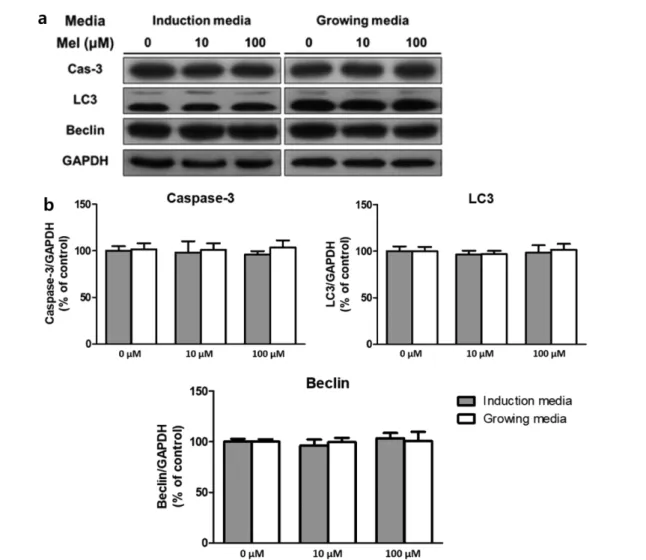
현이 감소하였다. 한편, C/EBPβ의 발현은 두 단계 모두에 서 멜라토닌 농도가 100 µM일 때만 감소하여, 대조군과 비 교했을 때 분화 초기 단계는 약 30% (p<0.01), 분화 후기 단계는 약 20% (p<0.05)로 감소하였다. 마지막으로, 분화 중인 지방전구세포의 세포자살과 자가포식작용이 멜라토닌에 의해 유도되는지 확인하기 위하여 세포자살 인자인 caspase-3 과 자가포식작용 인자인 LC3, Beclin의 발현을 western blot 으로 확인하였다(그림 4). 그 결과, 세 인자 모두 두 분화 단 계에서 멜라토닌에 의한 발현의 변화를 보이지 않았으며, 특히 caspase-3의 활성화를 확인할 수 있는 cleaved caspase-3는 검 출되지 않았다.
IV. 고찰 및 결론
지방 조직은 triacylglycerol의 저장과 분해를 통하여 체내 지 질 대사의 균형을 조절하며 인슐린의 작용을 받아 혈당을 조절한다[18]. 이러한 대사 기능은 지방 조직의 최대 2/3를 차지하는 지방세포에서 대부분 유래하며, 이 지방세포가 지 방생성을 통해 과도하게 발달하거나 증식하여 대사 조절 기 능을 상실할 때 비만이 유발된다[2]. 지방세포는 완전히 분화한 상태에서는 분열하지 않으며, 간엽줄기세포(mesenchymal stem cell)에서 분화한 지방전구세포 상태에서 분열한 뒤 성 숙을 거쳐 발달한다[3]. 따라서 비만의 기전을 이해하기 위 하여 지금까지 3T3-L1 세포주를 이용한 지방전구세포의 분화
그림 2. 멜라토닌 처리 후 western blot으로 측정한 3T3-L1 세포의 신호전달경로 활성. (a) western blot 밴드, (b) p-ERK의 발현 변화. (n=3,*P<0.05 vs control of induction media, #P<0.05, ##P<0.01 vs control of growing media)
Fig. 2. Activity of signaling pathway in 3T3-L1 cells measured with western blot analysis, after melatonin was treated. (a) Bands of western blot analysis, (b) Change of p-ERK expression. (n=3, *P<0.05 vs control of induction media, #P<0.05,
##P<0.01 vs control of growing media)
142
과정에 대한 연구가 이루어졌으며, 지방전구세포의 수를 감 소시키고 지방생성을 억제할 수 있는 호르몬이나 사이토카 인(cytokine), 그리고 외래 물질들을 발견하려는 연구가 진 행 중이다. 본 실험에서는 수면 시간의 저하에 의해 비만 유 병률이 증가한다는 연구 결과들에 근거하여[14,19,20], in vitro 환경에서 멜라토닌이 분화 중인 지방전구세포에 어떠 한 영향을 주는지 분석하기 위한 실험을 진행하였다.
지방전구세포의 분화 도중에 관찰할 수 있는 주요 형태학적 특성 중 하나로 세포 내의 지질 축적 증가가 있으며, 이는 지방세포의 주요 특성 중 하나이다. 따라서 분화 과정에 따른 지방 축적 정도를 확인함으로써 지방생성이 촉진되었는지 억제되었는지 알 수 있다. 본 연구에서 oil red O staining 결과를 통해 induction 단계에서는 멜라토닌 처리 여부와 관련없이 아직 지방 과립이 크게 발달하지 않았다는 것을
알 수 있었으며, 멜라토닌에 의한 형태학적 차이점 또한 발 견하지 못하였다. 반면 growing 단계에서는 지방 과립의 축 적이 일어났으며, 멜라토닌에 의해 과립의 면적이 감소하여 지방세포의 형태학적 변화가 억제된 것을 확인하였다.
이러한 외부 ligand에 의한 지방전구세포의 분화기전은 ERK pathway에 의해 그 신호가 핵으로 전달된다. ERK pathway는 p38, JNK 와 함께 mitogen activated protein kinases (MAPKs) pathway 를 구성한다. PPARγ, C/EBPs 등의 지방생성 인자들의 발현 또한 ERK pathway의 조절을 받으며[7], ERK pathway를 억제할 시 in vitro와 in vivo 모델 모두에서 지방생성이 억 제되는 것으로 알려져 있다[21]. 멜라토닌에 의한 각 분화 단계에서의 전사 인자의 변화를 살펴보았을 때, induction 단계와 growing 단계 모두에서 ERK pathway와 지방생성 인 자의 발현이 감소하였으며, 멜라토닌이 지방전구세포의 분
그림 3. 멜라토닌 처리 후 western blot으로 측정한 3T3-L1 세포의 지방생성인자 활성. (a) western blot 밴드, (b) C/EBPα, C/EBPβ, PPARγ의 발현 변화. (n=3, *P<0.05, **P<0.01, ***P<0.001 vs control of induction media, #P<0.05, ##P<0.01, ###P<0.001 control of growing media)Fig. 3. Activity of adipogenic factors in 3T3-L1 cells measured with western blot analysis, after melatonin was treated. (a) Bands of western blot analysis, (b) Change of C/EBPα, C/EBPβ, and PPARγ expression. (n=3, *P<0.05, **P<0.01, ***P<0.001 vs control of induction media, #P<0.05, ##P<0.01, ###P<0.001 control of growing media)
143
화를 억제한 것으로 보인다. 이는 oil red O staining에서 관찰 된 형태학적 결과를 뒷받침한다. 두 분화 단계 간의 멜라토 닌에 의한 차이를 비교해 보았을 때, 전체적으로 induction 단계보다 growing 단계에서 감소량이 더 크게 나타났으나 C/EBP β는 induction 단계에서 더 크게 감소하였다. 기존의 연 구에 따르면 이 지방생성 인자들은 세포의 분화 단계에 따라 단계적으로 발현하며, 분화 초기에는 C/EBPβ, δ가, 그 이후로 PPAR γ와 C/EBPα, β가 우세하게 작용하는 것으로 알려져 있다[7,18]. 따라서, 본 결과는 분화 초기에 PPARγ와 C/
EBP α가 C/EBPβ에 비해 상대적으로 약하게 발현하여 멜 라토닌의 영향을 덜 받은 것으로 판단된다.
한편, 해당 농도에서 멜라토닌에 의한 지방전구세포의 세 포자살과 자가포식작용은 일어나지 않았다. 멜라토닌은 전 형적인 항산화 물질이지만[14,22], mM 단위의 고농도로 투 여할 시 세포에 따라 산화 물질로도 작용하는 것으로 알려
져 있다[23-25]. 멜라토닌과 세포 생존 간의 연관성을 분석 한 선행 연구에 따르면, 멜라토닌을 몇 종류의 암세포와 신 경세포 등에 고농도로 투여함으로써 세포자살을 유도하는데 성공한 반면에[23-25], 다른 일반적인 세포에 멜라토닌을 투 여하는 경우에는 오히려 세포자살을 예방하는 결과 또한 나 타났다[26,27]. 또한 인간의 야갼 혈중 멜라토닌 농도는 20~40 세 성인 기준 80~120 pg/ml로[28], 1 nM을 넘지 못한다 는 것을 고려해 볼 때 멜라토닌에 의한 지방생성의 감소 효 과는 세포자살이나 자가포식작용과 무관하며 임상적으로도 이러한 기전을 통한 효과는 기대하기 어려울 것으로 보인다.
멜라토닌은 시상하부(hypothalamus)의 시교차 상핵 (suprachiasmatic nucleus) 에서 감지하는 빛의 유무에 의 해 분비량이 조절된다. 어두운 환경이 지속될수록 멜라토닌의 혈중 농도는 증가하며 하루 중 밤의 길이는 계절에 따라 바 뀌므로, 결국 하루 중 멜라토닌의 혈중 농도는 계절의 영향을
그림 4. 멜라토닌 처리 후 western blot으로 측정한 3T3-L1 세포의 세포자살 인자 활성. 각 농도별로 유의한 차이 없음. (a) western blot 밴드, (b) Caspase-3, LC3, Beclin의 발현 변화. (n=3)Fig. 4. Activity of programmed cell death factors in 3T3-L1 cells measured with western blot analysis, after melatonin was treated. (a) Bands of western blot analysis, (b) Change of caspase-3, LC3, and beclin expression (n=3)
144
받게 된다[14]. 지속적인 연구를 통해 계절과 일치하는 유 기체의 행동과 신진대사의 변화가 계절에 의한 멜라토닌의 농도 변화에 의해 조절된다는 것이 밝혀졌으며, 이는 결국 멜라토닌이 수면 패턴을 조절할 뿐만 아니라 지방 조직이나 내분비계에도 영향을 주어 체내 대사를 조절할 수 있다는 것을 의미한다[29,30]. 기존의 지방세포 연구에서, 멜라토닌을 투여하였을 때 지방전구세포나 간엽줄기세포의 지방생성 인 자들이 감소하였으며[17], 인슐린 민감성이 증가하거나 백 색지방의 갈변화가 관찰되는 등 지방세포의 지질 분해 기능 이 향상되어 체중 감량에 유리한 결과를 보였다[31]. 이러 한 결과들은 본 실험의 결과를 지지하고 있으며, 멜라토닌 이 지방 감소에 긍정적인 효과를 보일 수 있음을 나타낸다. 그 러나 in vitro 환경에서 멜라토닌의 농도가 생리학적 농도와 큰 차이를 보이고 있기 때문에, in vivo 환경에서 멜라토닌을 직접 투여하거나 일주기에 변화를 주었을 때 지방생성 억제를 통한 지방 감소 효과가 있는지 추가적인 연구가 필요할 것 으로 사료된다.
본 연구를 통하여 3T3-L1 지방전구세포의 분화 과정에서 각각 induction 단계와 growing 단계에 멜라토닌을 µM 단 위로 3일간 투여하였을 때 세포자살이나 자가포식작용의 효 과는 볼 수 없었으나, 지방생성의 주요 전사 인자들의 발현 감소를 통해 분화를 억제할 수 있음을 확인하였다. 또한 각 분화 단계에서 전사 인자의 발현에 차이가 존재하므로, 이에 따 라 멜라토닌에 의한 억제 효과도 induction 단계와 growing 단 계 간에 차이가 있음을 확인하였으며, growing 단계에서 억 제 효과가 더 크다는 것을 알 수 있었다. 멜라토닌은 지질 대사를 조절하는데 밀접한 관련이 있는 호르몬이므로, 멜라 토닌과 지방세포간의 연관성을 규명함으로써 생체 리듬 조 절이나 직접적인 멜라토닌 투여를 활용한 비만 치료법을 고 려해 볼 수 있을 것이다.
References
[1] Formiguera X, Cantón A. Obesity: epidemiology and clinical aspects. Best Pract Res Clin Gastroenterol. 2004;18(6):1125- 46.
[2] Kopelman PG. Obesity as a medical problem. Nature. 2000;
404(6778):635.
[3] Flegal KM, Carroll MD, Kit BK, Ogden CL. Prevalence of obesity and trends in the distribution of body mass index among US adults, 1999-2010. JAMA. 2012;307(5):491-7.
[4] Wellman NS, Friedberg B. Causes and consequences of adult obe- sity: health, social and economic impacts in the United States.
Asia Pac J Clin Nutr. 2002;11:S705-9.
[5] Itani O, Jike M, Watanabe N, Kaneita Y. Short sleep duration and health outcomes: a systematic review, meta-analysis, and meta-regression. Sleep Med. 2017;32:246-56.
[6] Loredo JS, Weng J, Ramos AR, Sotres-Alvarez D, Simonelli G, Talavera GA, Patel SR. Sleep Patterns and Obesity: Hispanic
Community Health Study/Study of Latinos Sueño Ancillar Study.
Chest. 2019;156(2):348-56.
[7] Lefterova MI, Lazar MA. New developments in adipogene- sis. Trends Endocrinol Metab. 2009;20(3):107-14.
[8] Kang MC, Ding Y, Kim HS, JeonYJ, Lee SH. Inhibition of Adipogenesis by Diphlorethohydroxycarmalol (DPHC) through AMPK Activation in Adipocytes. Mar Drugs. 2019;17(1):44.
[9] Kim KS, Jang MJ, Fang S, Yoon SG, Kim IY, Seong JK, Yang HI, Hahm DH. Anti-obesity effect of taurine through inhibition of adipogenesis in white fat tissue but not in brown fat tissue in a high-fat diet-induced obese mouse model. Amino Acids. 2019;51(2):245-54.
[10] Wang Y, Jones PJ. Conjugated linoleic acid and obesity con- trol: efficacy and mechanisms. Int J Obesity. 2004;28(8):941.
[11] Goldman S, Zhang Y, Jin SV. Autophagy and adipogenesis:
implications in obesity and type II diabetes. Autophagy. 2010;
6(1):179-81.
[12] Vermeulen K, Van Bockstaele DR, Berneman ZN. Apoptosis:
mechanisms and relevance in cancer. Ann Hematol. 2005;
84(10):627-39.
[13] Singletary K, Milner J. Diet, autophagy, and cancer: a review. Can- cer Epidemiol Prev Biomarkers. 2008;17(7):1596-610.
[14] Cipolla-Neto J, Amaral F, Afeche S, Tan D, Reiter RJ. Mel- atonin, energy metabolism, and obesity: a review. J Pineal Res. 2014;56(4):371-81.
[15] Kato H, Tanaka G, Masuda S, Ogasawara J, Sakurai T, Kizaki T, Ohno H, Izawa T. Melatonin promotes adipogenesis and mitochondrial biogenesis in 3T3?L1 preadipocytes. J Pineal Res. 2015;59(2):267-75.
[16] Zwirska-Korczala K, Jochem J, Adamczyk-Sowa M, Sowa P, Polaniak R, Birkner E, Latocha M, Pilc K, Suchanek R.
Influence of melatonin on cell proliferation, antioxidative enzyme activities and lipid peroxidation in 3T3-L1 preadipocytes-an in vitro study. J Physiol Pharmacol. 2005;56:91.
[17] Zhang L, Su P, Xu C, Chen C, Liang A, Du K, Peng Y, Huang D.
Melatonin inhibits adipogenesis and enhances osteogenesis of human mesenchymal stem cells by suppressing PPARγ expression and enhancing Runx2 expression. J Pineal Res.
2010;49(4):364-72.
[18] Smas CM, Sul HS. Control of adipocyte differentiation. Bio- chem J. 1995;309(Pt 3):697.
[19] Contreras-Alcantara S, Baba K, Tosini G. Removal of mela- tonin receptor type 1 induces insulin resistance in the mouse.
Obesity. 2010;18(9):1861-3.
[20] Reiter RJ, Tan DX, Korkmaz A, Ma S. Obesity and metabolic syndrome: association with chronodisruption, sleep depriva- tion, and melatonin suppression. Ann Med. 2012;44(6):564-77.
[21] Prusty D, Park B, Davis KE, Farmer SR. Activation of MEK/
ERK signaling promotes adipogenesis by enhancing peroxi- some proliferator-activated receptor γ (PPARγ) and C/EBPα gene expression during the differentiation of 3T3-L1 preadipocytes. J Biol Chem. 2002;277(48):46226-32.
[22] Reiter RJ, Tan Dx, Mayo JC, Sainz RM, Leon J, Czarnocki Z. Melatonin as an antioxidant: biochemical mechanisms and pathophysiological implications in humans. Acta Biochim Pol. 2003;50(4):1129-46.
[23] Bizzarri M, Proietti S, Cucina A, Reiter RJ. Molecular mech- anisms of the pro-apoptotic actions of melatonin in cancer: a review. Expert Opin Ther Targets. 2013;17(12):1483-96.
[24] Fernández A, Ordóñez R, Reiter RJ, González-Gallego J,
145
Mauriz JL. Melatonin and endoplasmic reticulum stress: relationto autophagy and apoptosis. J Pineal Res. 2015;59(3):292-307.
[25] Hong Y, Won J, Lee Y, Lee S, Park K, Chang KT, Hong Y.
Melatonin treatment induces interplay of apoptosis, autophagy, and senescence in human colorectal cancer cells. J Pineal Res.
2014;56(3):264-74.
[26] Fu Y, Tang M, Fan Y, Zou H, Sun X, Xu X. Anti-apoptotic effects of melatonin in retinal pigment epithelial cells. Front Biosci. 2012;17(4):1461-8.
[27] Pazar A, Kolgazi M, Memisoglu A, Bahadir E, Sirvanci S, Yaman A, Yeğen BÇ, Ozek E. The neuroprotective and anti- apoptotic effects of melatonin on hemolytic hyperbilirubin- emia-induced oxidative brain damage. J Pineal Res. 2016;60(1):
74-83.
[28] Karasek M, Winczyk K. Melatonin in humans. J Physiol Pharmacol. 2006;57:19.
[29] Favero G, Stacchiotti A, Castrezzati S, Bonomini F, Albanese M, Rezzani R, Rodella LF. Melatonin reduces obesity and restores adipokine patterns and metabolism in obese (ob/ob) mice. Nutr Res. 2015;35(10):891-900.
[30] Kayaba M, Park I, Iwayama K, Seya Y, Ogata H, Yajima K, Satoh M, Tokuyama K. Energy metabolism differs between sleep stages and begins to increase prior to awakening. Metabo- lism. 2017;69:14-23.
[31] Fernández Vázquez G, Reiter RJ, Agil A. Melatonin increases brown adipose tissue mass and function in Zücker diabetic fatty rats: implications for obesity control. J Pineal Res. 2018;
64(4):e12472.