약학회 지 제 48 권 제 6 호 352~357 (2004)
Yakhak Hoeji Vol. 48,No. 6
稱 學 舍 확
이노시톨 삼인산 수용체와 미토콘드리아에 의한 심방 근세포 Ca2+
신호전달의 조절
이향진 • 라스클리만* ■마틴모라드* • 우선희# 충 남 대 학 교 약 학 대 학 , * 미 국 조 지 타 운 대 학 교 의 과 대 학 약 리 학 교 실
(Received November 2,2004; Revised November 16,2004)
Regulation of Atrial Ca2+ Signaling by Inositol 1,4,5-Trisphosphate Receptor and Mitochondria
Hyang-Jin Lee, Lars Cleemann*, Martin Morad* and Sun-Hee Woo#
College of Pharmacy, Chungnam N ational University, Daejeon 305-764, Korea
^Department of Pharmacology, Georgetown University Medical Center, Washington, D C 20057 USA
Abstract — Atrial myocytes have two functionally separate groups of ryanodine receptors (RyRs): those at the periphery
colocalized with L-type Ca2+ channels (DHPRs) and those at the cell interior not associated with DHPRs. Ca 2+ current (ICa) directly gates peripheral RyRs on action potential and the subsequent peripheral Ca 2+ release propagates into the center of atrial myocytes. The mechanisms that regulate the Ca 2+ propagation wave remain poorly understood. Using 2-D confocalCa2+ imaging, we examined the role of inositol 1’4’5-trisphosphate receptor (IP 3R) and mitochondria on I Ca-gated local Ca2+
signaling in rat atrial myocytes. Blockade of IP 3R by xestospongin C (XeC) partially suppressed the magnitudes of I Ca-gated central and peripheral Ca2+ releases with no effect on I Ca. Mitochondrial staining revealed that mitochondria were aligned with =2-fxm separations in the entire cytoplasm of ventricular and atrial myocytes. Membrane depolarization induced rapid mitochondrial Ca2+ rise and decay in the cell periphery with slower rise in the center, suggesting that mitochondria may
immediately uptake cytosolic Ca2+, released from the peripheral SR on depolarization, and re-release the Ca 2+ into the cyto
sol to activate neighboring central RyRs. Our data suggest that the activation of IP 3R and mitochondrial Ca2+ handing on action potential may serve as a cofactor for the Ca2+ propagation from the DHPR-coupled RyRs to the DHPR-uncoupled
RyRs with large gaps between them.Keywords □ atrial myocyte, Ca2+ signaling, mitochondria, inositol 1,4,5-trisphosphate receptor
심장근세포의수축은활동전위(action potential)에의해유도 되는세포질내의 일시적 칼슘이온농도([Ca2+]c) 증가에의해 일 어난다
.
활동전위 발생 시세포막의 탈분극은전압의존성 L-형 Ca2+ 채널(L-type Ca2+ channel)을열리게하고,
열린채널들을 통해 소량의 Ca2+이세포 내로유입된다.
유입된 Ca2+에의해 세포근형질망(sarcoplasmic reticulum , SR) 막에있는 Ca2+ 유 리채널즉ryanodine receptors(RyRs)가열려 많은양의 Ca2+이세포질로유리된다
.1"4)
활동전위에 의해일어나는이러한세 포Ca2+ 신호전달(signaling)은그국부적인양상이 심실과심방 에서 현저히 다른데,
이는주로 세포막의 함몰구조 즉t-tubule 이심실에는존재하나심방근세포에는거의존재하지 않기 때#본논문에 관한 문의는 저자에게로
(
전화) 042-821-5924 (
팩스) 042-823-6566 (E-mail) [email protected]
문이다
.5)
따라서 세포주변부(periphery)에위치한SR은세포막 과접합(peripheral junction)을이루나,
세포내부(center)의SR 은그러한세포막접합을형성하지 않는다(non-junctional SR).51 이러한이유로주변접합부SR에있는RyRs는L-형Ca2+ 채널 과매우 가까이(약 1 0 nm ) 위치하지만,
세포 내부 SR에있는 RyRs는L-형Ca2+ 채널과의 접촉이 없다.61
심방근세포에서는활동전위 발생시세포막L-형Ca2+ 채널 이열려근처 접합부SR 막에 있는RyRs를먼저 열리게 한다
.
주변접합부RyRs에의해세포질로유리된 Ca2+은세포내부로 이동하여 이웃한RyRs를차례로자극하며 열리게 한다.
이러한 이유로활동전위 시에심방근세포에서는세포주변부에서 내부 로Ca2+이이동하는 "Ca2+ 전파 웨이브(propagation wave)”가 발생한다. 7
ᅵ9)
이심방근세포 Ca2+ 전파웨이브의속도는,
세포 막전압이 변하여 L-형Ca2+ 채널에 의해유입되는 Ca2+ 양이 달라져도일정하게유지되며Ca2+의단순확산속도보다느려서9>
352
미토콘드리아와이노시톨삼인산수용체에 의한심방킬슘조절
353
L
-형Ca2+
채널 이외에Ca2+
전파웨이브를조절하는다른세 포인자가존재할것으로 생각되고있다.
심방근세포의 요묘게는
RyRs
뿐만아니라IP3R
가존재하며 이들두단백질은세포주변부에서매우가까이위치한다고알려져
있다
.10)
또한심방근세포SR
근처에는미토콘드리아들이분포하며
,
최근미토콘드리아세 있는여러종류의이온수송체가세포질Ca2+
양을조절한다고보고되고있다.11_13)
따라서본연구에서는RyRs, IP3R,
미토콘드리아간의상호신호조절이Ca
2+을매개로 일어날수있다는가설과이상호신호조절이 탈분극시발생하는Ca2+
전파웨이브를조절한다는가설을검증하고자하였다.
실험 방법
단 일 심 방 근 세 포 분 리
심방 근세포의분리는
Woo
등9>의 방법으로시행하였다.
즉,
흰쥐수컷(Sprague-Dawley,
체중2()()~300 g
)을sodium pento- barbital(150m&acg,
복강투여)로마취한후심장을적출하였다-
심장의 대동맥을Langendorff
관류장치의cannular
에연결하여 고정한후대동맥안으로OmM [Ca2+]-Tyrode
용액((단위: mM) 137 NaCl, 5.4 KC1, 10 HEPES, 1 MgCl2, 10 Glucose, pH 7.4)1
:37°C
에서5
분간관류시켰다.
이어서OmM [Ca2+]-Tyrode
용액에collagenase(Roche, 1.4mg/m
/)와protease(Type XIV Sigma, 0.16 mg/m
/)를포함한용액을12
분간관류시킨다음마 지막으로0.2 mM [Ca
2+]을포함한Tyrode
용액을약7
분간관 류시켜사용된 효소를제거하였다.
심장을Langendorff
장치로 부터분리한후심방을취하여25 mm petri dish
에담긴0.2 mM [Ca2+]-Tyrode
용액에 두었고,
조직을 여러 번chopping
한早,mechanical dispersion
을가하였다.
분리된세포들은실험에 사 용되기 전까지실온에서0.2 mM [Ca2+]-Tyrode
용액에 보관하 였다.
세 포 질 및 미 토 콘 드 리 아 의
Ca2+
영 상 화Ca2+
영상화는Woo
둥9>의방법으로 시행하였다.
즉, Zeiss Axiovert TV135
도립현미경(x 40, water immersion objective)
에 연결된2-D laser scanning confocal microscopy system (Noran Odyssey XL
)을사용하였다. Argon
이온레이저를이용 하여488nm
또는544 nm
의빛으로fluo-3
와rhod-2
형광지시 약을각각excitation
시켰고>510 nm
및>570
의형광emis
sion
을PMT(Hamamatsu, NJ, USA
)를 이용하여 각각 감지하 였다. y
방향의scanning
속도는30Hz(Fig. 5)
또는240 Hz (Fig. 1
)로 조정하였다. Workstation(Indy, Silicon Graphics)
의Intervision 2D acquisition system
을'
이용하여 영상을기록하였 고자체 개발한”Con2”
컴퓨터 프로그램(Visual Basic 6.0
으로 작성)의pixel
측정법으로형광영상을정량하였다. Fluo-3
는세포질
Ca2+
농도측정을위해patch pipette
용액에첨가하여 세 포내로직접투석시켰으며,rhod-2
는미토콘드리아Ca
2+을측정 하기 위해AM
형태(5|iM
,30
분)로세포 외부에서 처리하여 세 포내로도입하였다. Ca2+
농도는6ᅵ무런처리를하지 않았을때 의바탕 형광강도(F
0)에대해각이미지의 형광강도(F
)를표준 화한값즉,F/I
^로나타내었다.
미토콘드리아영상측정
심실과 심방근세포에 미토콘드리아를선택적으로 염색하는
MitoTracker 0range(200 nM, Molecular Probes
)를포함한정상Tyrode
용액을약30
분간 처리한후 영상을측정하였다.
영상 은confocal laser scanning microscope system(LSM 510, Carl Zeiss, X40, oil immersion, argon laser; KBSI
)을 사용하 여Zeiss LSM image analyzer
로기록하였으며,
영상의 분석은OLYSIA BioReport(Olympus
)로시행하였다(Fig. 4).
Patch-clamp
방법심방근세포를 이용한
patch-clamp
의시행은Ham ill
등14>의 방법에 따라whole cell mode
에서 시행하였다.
심방근세포 내 에Cs
+이 풍부한(Cs+-rish) pipette internal solution
((단위: mM) 0.4 Fluo-3, 110 CsOH, 110 Aspartic Acid, 5 NaCl, 20 TEA-C1, 10 HEPES, 5 Mg-ATP; CsOH
를 이용하여pH
를7.2
로보정)을투석 시켰으며pipette
의저항은2.5-3.5MQ
이었다. L
-형Ca2+
전류를기록하기위해K+
및Na+
채널을zero [K +]0 (Cs
+으로교환) Tyrode
용액과tetrodotoxin(30 |iM
>을사용하여 각각 억제하였다. Patch clamp amplifier(PC-l, Dagan Co
.)로clamp
를시행하였고 전류를모니터링 하였다.
전류의 기록과 전압고정 프로토콜은
A/D converter
를통해pCLAMP software
(version 9, Axon Instruments
)로수행하였다.
단일 세포외부에 특정약물을처리할때에는고속자동용액교환장치를사용하였다.
통계
실험결과는평균
±
표준치•로 표시하였으며 유의성은Student's Mest
를사용하여 표시하였다.
실험 결 과 및 고 찰
세포질
Ca2+
유리에 대한IP3R
억제제효과활동전위에 의해 유도되는심방근세포의
Ca2+
전파 웨이브 가심방세포에 존재하는IP 3 R
에의해조절되는지알아보기 위 하여IP 3 R
에대한선택적 억제제xestospongin C(XeC
)를처리 하여 고찰하였다.
먼저 정상조건에서L
-형Ca2+
전류를막전압 을-80mV
에서+60mV
로고정하여 유도하였을 때(Fig. 1A)
심 방근세포의주변부(PERI
)와중심부(CEN
)에서 각각Ca
2+의변354
이향진•
리스클리만.
마틴모라드.
우선희A +60 mV
-80 mV
5pA /pF
B
20 ms
F / R
Fig. 1 — Effect of xestospongin C on atrial local Ca2+ releases. (A) Superimposed membrane currents elicited by the de
polarizing pulse (upper trace) in the presence or abscence of xestospongin C (XeC, 5
m-M). (B) Superimposed peri
pheral (PERI) and central (CEN) local Ca2+ transients, measured from the confocal Ca2+ images (240 Hz), on depolarization in the control condition and after incubation with XeC. (Inset) Image of pixel masks that were used to measure peripheral (two domain in the cell border) and central Ca2+ signal of atrial cell area.
화
C’Ca2+ transients
1’)가관찰되었다(Fig. IB). 5fiM XeC
가포함 된세포외액을동일세포에처리한후같은전압변화를일으켰 을때L
-형Ca2+
전류에는별변화가없었으나(Fig. 1A)
세포주변부와내부에서 유리되어중가하는
Ca2+
양이현저히 감소되었음을
.
관찰하였다(Fig. IB). XeC
로처리한7
개의 심방근세포에 서유사한 결과를관찰하였으며,
그평균Ca2+ transient
억제 효과는 세포주변부에서41±3.2%,
세포내부에서49±5.1
%로 각각관찰되어통계적으로유의한효과를보였다(n=7,
P<0.01).Ic
a는control
과XeC
처리 시 유사하게 관찰되었다(pA/pF:
Control, 1.12±0.22; XeC, 0.97±0.18
,n=7, P>0.05).
이결과 는막전압의 탈분극시SR RyRs
에의한Ca2+
유리뿐만아니라IP 3 R
에의한Ca
2+의유리도일부기여함을나타낸다.
쥐심방근세포에서
IP 3 R
는세포주변부RyRs
와근접하여 있으며 내부에는거의 존재하지 않는다고알려져 있다
.10) XeC
에의해억제되는내부
Ca2+
유리는(Fig. IB, CEN) XeC
에의해줄 어든 세포주변부Ca
2+에의해내부로 확산되는Ca2+
양이 감소하여 내부
RyRs
의활성에 영향을주었기 때문으로사료된다.
주변부
RyRs
에의해 먼저중가한Ca
2+이 직접 또는간접적인 기전에 의해IP 3 R
를자극하여SR
로부터더많은Ca2+
유리를 일으켰으리라고추정되며이러한Ca2+
유리 촉진과정은주변부RyRs
과바로이웃하는 세포내부RyRs
사미에 존재하는卿10)
을극복하고
Ca2+
전파웨이브가성공적으로일어날수있는하나의기전이될것으로사료된다
.
심 근 세 포 내 미 토 콘 드 리 아 의 분 포
활동전위에 의해유도되는심방근세포의
Ca2+
전파웨이브 가심방세포에존재하는미토콘드리아께의해조절되는지 알아Fig. 2 Mitochondrial staining in rat ventricular myocyte. (A) Raw cell image. (B) Confocal visualization of mitochondria in the whole myocyte. (C) Magnified image from the boxed area in panel (B).
Fig. 3 Mitochondrial staining in rat atrial myocyte. (A) Raw cell image. (B) Confocal visualization of mitochondria in the whole myocyte. (C) Magnified image from the boxed area in panel (B).
J. Pharm. Soc. Korea
미토콘드리아와이노시톨삼인산수
§•
체에의한심방킬슘조절355
보기 위해 미토콘드리^•를 특이적으로 염색하는
MitoTracker
Orange
를사용하여심근세포내미토콘드리아의 분포를고찰하였다
.
관찰된미토콘드리아는심실과심방근세포모두에서 세 포전면에규칙적으로분포되어 있었다(Fig. 2, 3).
심실근세포 에서 미토콘드리아는세포의longitudinal direction
0■로배열되 어있었으며(Fig. 2B, C) transverse direction0-?-
미토콘드리아Fig. 4 - Regular arrangement of mitochondria within the atrial myocyte. (A) Lower trace indicates fluorescence intensity measured by transverse line scanning (see the single straight line). Mitochondria were aligned in a longitudinal direction with 2-(im interval throughout the cytoplasm except nucleus. (B) Lower traces indicate fluorescence intensities measured by longitudinal line scanning (see the two straight lines). Note that the signal is denser around the nucleus ("N"). In each 2-(im domain mitochondrial signal was divided by double peaks.
사이에 어둡게보이는밴드가약
2 |am
간격의 규칙적인배열을 이루고있음을관찰하였다(Fig. 2C,
화살표).
심방근세포에서도 심실과유사하게 미토콘드리아가longitudinal
방향의 규칙적인 배열을이루고있었고(Fig. 3B) 2 nm
간격의transverse band
로 관찰되었다(Fig. 3C).
어둡게보이는이transverse band
는미토 콘드리아사이에 위치하는SR
일가능성이 가장크다.5>
Longitudinal direction
0■로배열된심방미토콘드리아형광신 호를측정하면 약2 nm
마다신호가중가됨을관찰할수 있는데
(Fig. 4A),
이는미토콘드리아사이에같은방향으로근섬유가규칙적으로 배열되기 때문이다
.5)
심방근세포핵의 가장자리에 는형광신호가보다강하•게기록되었고이는미토콘드리아의 높 은밀도를나타낸다(Fig. 4B,
화살표).
탈분극에의해유도된 미토콘드리아내
Ca2+
양의 변화Longitudinal direction
으로규칙적인배열을 이루는 미토콘드 리아는세포주변부에서 내부로이동하는Ca
2+에 대한물리적인 장애 벽으로작용한다.
기존 결과들에 의하면 이미토콘드리아 내벽에는 다양한이온 수송체가존재한다고 알려져 있고,15)
특 히최근에는그중Ca
2+을수송하는단백질들이 세포질Ca2+
농 도를 생리학적 조건에서 빠른 속도로조절한다고 보고된 바 있다12 ’13)
탈분극에 의해유도되는세포질
Ca
2+의 전파웨이브기"
미토 콘드리아에 의해 조절되는지 알아보기 위해 미토콘드리아 내Ca2+
농도([Ca
2+]m)가 탈분극에 의해 변화되는지조사하였다.
단일심방근세포에 고농도
K+(25mM
)을 포함한용액을 외부에단시간
(500 ms)
빠른속도로(용액 교환속도:
약50 ms)
처 리하여 세포탈분극을 일으키고이와동시에 측정된 미토콘드리아
Ca2+
형광영상으로부터 세포주변부와내부의 형광을 측정하여변화를관찰하였다
.
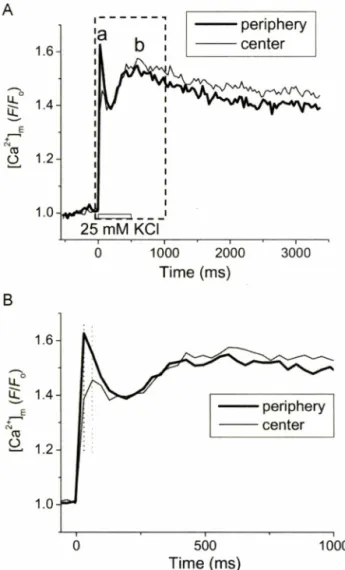
탈분극직후세포주변부[ C a ^ #
일 시적으로 급격히중가하였다가 감소하였고(Fig. 5A, a),
약250 ms
후부터 다시 서서히 중가하였다(Fig. 5A, b).
세포 중심부[Ca2^ #
주변부에비해약30 ms
지연되어 최고치에 도달하였 고(Fig. 5B,
점선참조)
다시 소량 감소후서서히 중가하였다.
초기 탈분극직후
[Ca2+]„^
증가는중심부에 비해주변부에서훨씬 컸고
(
스F/F 0 :
주변,0.69±0.11;
중심, 0.35±0.06
,n=14,
P<0.05)
후기 느린 속도의 증가는두 지역에서 유사한 정도로나타났다
(AF/F 0 :
주변, 0.52±0.12;
중심,0.55±0.08, n=14, P>0.05).
초기 미토콘드리아Ca2+
농도의 중가는 탈분극시L-
형Ca2+
전류에 의해 주변부SR
로부터 세포질로 유리된Ca2+
이미토콘드리아내로들어가기
(uptake)
때문이다.
이는주변부[Ca
2+]m가세포내부보다현저히빠르고많이증가한것으로알 수있다.
그후[Ca
2+]m가감소하는것은Ca
2+이미토콘드리아 밖으로다시 유출된 것을 나타내며,이Ca2+
유출은 주변부 세 포질Ca
2+의증가뿐만아니라내부의Ca
2+을증가시킬 수있다.
356
이향진.
라스클리만.
미틴모라드 우선희B 1.0
25 m M KCI
0 1000 2000
T im e (m s)
3000
0 500 1000
T im e (m s)
Fig. 5 - Effect of high K + pulse on mitochondrial Ca2+ in rat atrial myocyte. (A) Peripheral and central Ca2+ transients measured from 2-D Rhod-2 confocal fluorescence images (30 Hz). (B) Expanded recording of Ca2+ transients for the time marked by the box in panel (A).
특히주변부
[Ca2+] ^
감소폭이현저히큰데이는초기uptake
양이 많은것과관련이 있을것이다.
이차적으로느리게 일어나는 증가는 이차적인 세포질
Ca2+
중가에 기인할 것이다
.
실제 활동전위 지속기에 상응하는생리학적 조건에서는1
단계로 빠르게 일어나는[Ca2+]m transient(Fig. 5Aa)
만해당된 다. 250 ms
탈분극후변하는[Ca2+]m^r
지속되는탈분극에 의 해주로작동하는세포막Na+-Ca2+ exchanger
에의한Ca2+
유 리과정일기능성이 가장크다.
결 론
본연구결과는심방근세포의흥분-수축연결과정
(excitation- contraction coupling)
시 일어나는Ca2+
신호전달 과정을SR
RyRs
뿐만아니라IP3R
가조절하고있다는증거이며,
심방근세포내
Ca2+
중가가미토콘드리o
■ᅡ에의해조절되고 있음을또한 보여준다.
세포내부전면에 분포하는근섬유의 일률적인수축이곧심근의 수축력을결정하기 때문에심방세포내부
Ca2+
중가는심근의 수축력에직접적인 영향을준다
.
본연구는심방근세포에 현저하게 존재하는
ip3r
및미토콘드리아가 세포질Ca2+
증가에기여하고있음올보여주고있다.
이러한새로운Ca2+
조절기전은
IP 3 R
와미토콘드리아의중요한생리학적 기능을보 여줄뿐만아니라심장부전의 새로운발병 기전일수있으므 로,심장질환약물개발에 대한새로운표적 분자로써의응용가 치를가질것으로사료된다.
감사의 말씀
이논문은
2003
년도한국학술진흥재단의 지원에의하여 연구되었으며
(KRF-2003-042-E00009),
이에감사드립니다.
공촛점현 미경시스템을제공하여준한국기초과학지원연구원(KBSI)
과,
사 용을자세히보조해주신한국기초과학지원연구원이지원박사 님께감사드립니다.
문 헌
1) Fabiato, A .: Simulated calcium current can both cause calcium loading in and trigger calcium release from the sarcoplasmic reticulum of a skinned canine cardiac Purkinje cell. /. Gen.
Physiol. 85, 291 (1985).
2) Barcenas-Ruiz, L. and Wier, W. G. : Voltage dependence of intracellular [Ca2+]j transients in guinea pig ventricular myocytes. Circ. Res. 61, 148 (1987).
3) Cannell, M. B.
,Berlin, J. R. and Lederer, W. J. : Effect of membrane potential changes on the calcium transient in single rat cardiac muscle cells. Science 238, 1419 (1987).
4) Nabauer, M.
,Callewaert, G., Cleemann, L. and Morad, M. : Regulation of calcium release is gated by calcium current, not gating charge, in cardiac myocytes. Science 244,800 (1989).
5) Franzine-Armstrong, C., Protasi, E and Ramesh, V : Shape, size, and distribution of Ca2+ release units and couplons in skeletal and cardiac muscles. Biophys. J. 77,1528 (1999).
6) Carl, S. L., Felix, K., Caswell, A. H.
,Brandt, N. R., Ball, W. J.
Jr., Vaghy, R L., Meissner, G. and Ferguson, D. G. : Immunolocalization of sarcolemmal dihydropyridine receptor and sarcoplasmic reticular triadin and ryanodine receptor in rabbit ventricle and atrium. J. Cell Biol. 129(3),672 (1995).
7) Huser, J., Lipsius, S. L. and Blatter, L. A. : Calcium gradients during excitation-contraction coupling in cat atrial myocytes. J.
Physiol 494(Pt 3), 641 (1996).
8) Kockskamper, J., Sheehan, K. A., Bare, D. J., Lipsius, S. L., Mignery, G. A. and Blatter, L. A. : Activation and propagation
fM
o CD A
£
o
J. Pharm. Soc. Korea
미토콘드리아와이노시톨심인산수용체에 의한심방칼슘조절