Effects of dietary lipid sources on apoptotic and immune gene expression in head kidney of olive flounder (Paralichthys olivaceus)
Deokhwe Hur, Sang-Min Lee, Suhee Hong†8)
Department of Marine Biotechnology, Gangneung Wonju National University, Gangneung 210-702, South Korea
It can be hypothesized that dietary fatty acids can modulate immune responses in fish by inducing apoptosis of immune cells since dietary polyunsaturated fatty acid (PUFA) increase apoptosis by oxygen radicals generated by peroxidation. Thus we examined the effects of deferent dietary lipid sources such as squid liver oil (FO), linseed oil (LO) and soybean oil (SO) on oxidation (Cytochrome C oxidase; COS), apoptosis (TNF- Scinderin like) and immune (IL-1 and NKEF) gene expression in the main immune organ (head kidney) in olive flounder (Paralichthys olivaceus) by Q-PCR analysis after feeding diets containing each oil (5%) for 15 weeks. Linseed oil and soybean oil were chosen to compare n-3 or n-6 enriched vegetable oils, respectively. Consequently, COS, TNF-α and Scinderin like gene expression was increased in SO group, indicating the induction of oxidation and apoptosis. Meanwhile, no significant difference was found in immune gene expression. In conclusion vegetable oils containing n-3 PUFA like linseed oil seems to be more suitable lipid source than soybean oil for replacement of fish oil in flounder since n-6 PUFA in SO leads to activation of apoptosis pathways within the cellular damage in head kidney.
Key words :Olive flounder, Dietary lipid, Immune, Gene expression
High dietary fat levels commonly constitute fish diets as an important energy source for protein sparing and to decrease nitrogenous losses (Aksnes et al., 1996).
Fish oils have been used as main fat sources for fish feeds. Fish has higher amount of polyunsaturated fatty acids (PUFA) than mammals and PUFAs are major components of cellular membrane in fish. But there has been a concern that fish oil and fish meals are getting reduced (Barlow et al., 2001). The limited availability of fish resources has made researchers to seek sustainable alternative supplies of fish oils for the future development of aquaculture. A large number of vegetable oils
†Corresponding author : Suhee Hong
Tel : +82-33-640-2852 , Fax : +82-33-640-2955 E-mail : [email protected]
including palm, sunflower, borage, linseed, rapeseed, olive oil and soybean oil have been experimentally used as possible alternatives to fish oils (Torstensen et al., 2000; Tocher et al., 2001; Tocher et al., 2003; Rodríguez et al., 2002; Montero et al., 2005; Mourent et al., 2005).
Even though the substitution of fish oil to vegetable oils has been extensively studied and looked successful, it was known that use of vegetable oils can affect the fatty acid composition of fish tissue and plasma since vegetable oils mainly contain n-6 PUFA (linoleic acid) unlike fish oil (Torstensen, 2004). Fish oils are known to be rich sources of the n-3 PUFA like eicosapentaenoic acid (20:5n-3, EPA) and docosahexaenoic acid (22:6n-3, DHA) (Halliwell and Chirico, 1993). Cell membranes
in fish contain more n-3 PUFA than n-6 PUFA (Sargent et al., 1999). According to Pike (1999), the ratio of n-3/n-6 in most fish oils is 5-6:1 while the ratio in vegetable oils is 0-0.3:1 (Pike, 1999).
It is also known that dietary fatty acids can modulate immune responses and lipid related disorder in fish (Calder, 2002). It has been previously demonstrated in other studies that the modulation of dietary n-3/n-6 ratio by addition of vegetable oils or purified n-3 sources has affected resistance of fish to pathogens (Salte et al., 1988; Erdal et al., 1991; Waagbo et al., 1993;
Thompson et al., 1996). In contrast it was reported in Atlantic salmon that there were no significant differences between the fish oil and vegetable oil (rapeseed oil) groups in the gene expression of TNF- and IL-1 and in respiratory burst response (Seierstad et al., 2009).
However, they had a look at the indirect effect of dietary fatty acid after incubating head kidney cells with plasma from fish oil or vegetable oil fed fish. The direct effect of n-6 PUFA in fish diet has not yet been extensively studied at the gene expression level.
When investigating alternative oil sources main consideration was given to the growth performance, carcass nutrient profile and general health while the cytotoxicity of n-6 PUFA in fish was rarely studied.
In mammals, it is known that peroxidation of n-6 PUFA can cause apoptosis by damaging DNA. Ghosh et al.
(2006) demonstrated that consumption of a high fat diet rich in n-6 PUFA for only 4 weeks instigates mitochondrial nitrosative damage and causes cardiac dysfunction (Ghosh et al., 2006). Many recent studies have identified n-6 PUFA to cause mitochondrial damage and activation of transcription factors in the
vasculature (Toborek et al., 2002; Toborek et al., 1996;
Saraswathi et al.,2004; Pepe et al., 2005). In this study, therefore, the effect of n-6 PUFA on the expression of apoptosis and oxidative stress related gene i.e. Tumor necrosis factor- (TNF-), Cytochrome C oxidase (COS) and Scinderin like gene (ScinL) was assessed in fish.
There are two pathways of apoptosis. Type I is initiated from the cell surface death receptors (Fas, TNF family receptors) and involves activation of caspase-8, a cysteine protease cleaving at the C-side of aspartic acid (Budihardjo et al, 1999). The delivery of signals through the cell surface death receptor Fas or other receptors of the TNF family has emerged as a major pathway in the elimination of unwanted cells under physiological and disease conditions (Nagata, 1997).
Type II involves changes in mitochondrial integrity initiated by various effectors like Ca2+, reactive oxygen species (ROS), Bax, or ceramide, leading to the release of cytochrome c and activation of caspase 9 (Kadenbach et al., 2004). While TNF- is involved in type I pathway, ScinL are involved in type II pathway.
In this study it was also studied if n-3 PUFA enriched vegetable oil can affect those gene expressions since some vegetable oils contain high concentrations of n-3 PUFA than n-6 PUFA and they also can be used to substitute fish oil. Moreover, along with apoptosis related gene expression, the expression of immune related gene i.e. IL-1 and NK cell enhancing factor (NKEF) was also examined in this study. For this we have prepared fish diets containing fish oil, soybean oil or linseed oil to assess the effect of n-6 or n-3 PUFA enriched vegetable oils, on apoptosis and immune gene
expression in head kidney of flounder after feeding for 15 weeks.
Materials and methods
Experimental diets
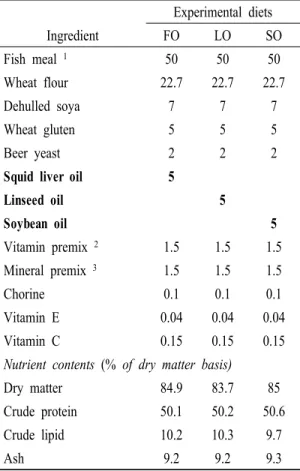
Three experimental diets were formulated to contain:
(i) 5% squid liver oil, (ii) 5% linseed oil, and (iii) 5%
soybean oil (Table 1). For the preparation of the experimental diets, calculated quantities of air-dry ingredients were thoroughly mixed in Hobart electric mixer. Experimental diets were made using a laboratory pellet machine after 30-40 g of water was mixed with 100 g mixture of ingredients, and dried overnight at room temperature. All diets were stored at -30°C until use.
Fish and feeding trial
Olive flounder Paralitichthys olivaceus were sourced from Uljin Marine Hatchery (Uljin-Kun, Korea). Olive flounder weighing around 8.5g were kept in circular 200L tanks at the Marine Biology Center for Research and Education at Gangneung-Wonju National University (Gangneung, Korea). The tanks were supplied with re-circulating seawater at an ambient temperature of approximately 20.9±1.2°C with 12/12h illumination and fish were fed a commercial pellet diet for two weeks prior to feeding the experimental diets. Groups of 40 fish were randomly distributed into nine 50L tanks. Each experimental diet was assigned to triplicate tanks. Fish were fed twice daily (9:00 and 17:00) for 15 weeks from June 17 to September 30, 2008. During each feeding, feed was offered by hand four to five times until satiation was reached. Fish were starved for 24h
Experimental diets
Ingredient FO LO SO
Fish meal 1 50 50 50
Wheat flour 22.7 22.7 22.7
Dehulled soya 7 7 7
Wheat gluten 5 5 5
Beer yeast 2 2 2
Squid liver oil 5
Linseed oil 5
Soybean oil 5
Vitamin premix 2 1.5 1.5 1.5
Mineral premix 3 1.5 1.5 1.5
Chorine 0.1 0.1 0.1
Vitamin E 0.04 0.04 0.04
Vitamin C 0.15 0.15 0.15
Nutrient contents (% of dry matter basis)
Dry matter 84.9 83.7 85
Crude protein 50.1 50.2 50.6
Crude lipid 10.2 10.3 9.7
Ash 9.2 9.2 9.3
1 Provided by Fisheries Co-op Feeds Co., Ltd. Gyeongsangnam province, Korea
2 Vitamin premix, contained the following amount which were diluted in cellulose (g/kg mix): L-ascorbic acid, 200;
DL-a-tocopheryl acetate, 20; thiamin hydrochloride, 5;
riboflavin, 8; pyridoxine hydrochloride, 2; niacin, 40;
Ca-D-pantothenate, 12; myo-inositol, 200; D-biotin, 0.4; folic acid, 1.5; p-aminobenzoic acid, 20; menadione, 4, retinyl acetate, chloecalciferol, 0.003; cyanocobalamin, 0.003.
3 Mineral premix, contained the following ingredients (g/kg mix): NaCI, 7; MgSO4ㆍ7H2O, 105; NaH2PO4ㆍ2H2O, 175;
KH2PO-l, 224; CaH4(PO4)2ㆍH2O, 140; Ferric citrate, 17.5;
Ca-lactate, 21.8; ZnSO4 . 7H2O, 2.8; ; CuCl,
Table 1. Composition and proximate analysis of the experiment diets (% of dry matter basis)
prior to sampling and anesthetized with MS222 (Woogene B&G, Korea). Fish of Each tank was bulk weighed at the beginning and end of the experiment.
Feed consumption, mortalities and feeding behavior were recorded daily.
Analysis of fatty acid composition of the experimental diets and flounder dorsal muscle
To analyze fatty acid composition of flounder body after feeding the diets for 15 weeks, all fish were killed and stored at -25°C in a freezer after removing head kidney for gene expression study. Chemical composition of the experimental diets and fish carcasses were determined using the following Association of Official Analytical Chemists procedures: dry matter by drying in an oven at 105°C for 24 h; crude protein (NX 6.25) by the Kjeldahl method after an acid digestion method using an Auto Kjeldahl System (Buchi, Switzerland);
crude lipid by ether extraction; ash by incineration in a muffle furnace at 550°C for 6 h and crude fiber by Fibertec automatic analyzer (Tecator, Sweden). Fatty acids in diets and fish carcasses were analyzed by using a HP-5890 II gas chromatograph (Hewlett-Packard, USA) with flame ionization detector, equipped with capillary column (HP, USA). The data were subjected to one-way analysis of variance (ANOVA) using the SPSS program version 7.5. Significant difference (P<0.05) among the means was determined by Duncan’s multiple range test.
Sample collection for gene expression study Three fish were taken randomly from the triplicate tanks of each treatment after 15 weeks of feeding and sacrificed using MS222 (Woogene B&G, Korea). Thus a total of nine fish were collected per treatment. All six groups were sampled on a particular day. Blood
was drawn from the caudal vein of individual fish and serum samples were obtained by centrifugation. Head kidney was removed and immediately frozen in liquid nitrogen, followed by storage at -80℃ until RNA extraction.
RNA extraction and first-strand cDNA synthesis Total RNA was extracted using Trizol Reagent (Invitrogen, USA) following the manufacturer’s protocol. RNA concentration was determined by optical density reading at 260 nm using NanoDrop (Thermo, USA), and integrity was verified by ethidium bromide staining of 28 S and 18 S ribosomal bands on a 1%
agarose gel. Four microgram of total RNA from each sample was reverse-transcribed using MMLV reverse transcriptase (Promega, USA) and oligo dT primers (Promega, USA). RNA samples and cDNA were stored at -80℃ and -20℃, respectively, until further use.
Analysis of immune related gene expression by real time PCR analysis
Real time PCR was performed using SYBR Green II and an ABI real time PCR 7300 Amplification Detection System (Applied Biosystems, USA). Flounder HSC70 (heat shock cognate 70, NCBI GenBank Accession No. gi39979268) was used as a housekeeping gene to normalize the potential variations in RNA loading (Table 2). Real time specific primers for target genes (Cytochrome C oxidase subunit III; NCBI GenBank Accession No.
gi9695366, Tumor necrosis factor; NCBI GenBank Accession No. gi7670053, Interleukin 1 beta; NCBI GenBank Accession No. gi19911214, Natural killer enhancing factor; NCBI GenBank Accession No. gi63002582,) were designed (Table 2). The 20㎕ real time PCR reactions
Gene or Clone similar to: GenBank accession no. Primer name Primer sequence Cytochrome C oxidase subunit III gi9695366 COS3 F 5'-CTTCGTTGCCACTGGTTTCC-3'
COS3 R 5'-TCATGCGGCTGCTTCGA-3'
Tumor necrosis factor- gi7670053 TNF F 5'-AGGAGGCAGCGGAAAAACA-3'
TNF R 5'-TAGGCGTCCTCCTGACTCTTCT-3'
Scinderin like gene This lab ScinL F 5'-CCCTCCGCTGTGGATGATC-3'
ScinL R 5'-GTGCGGCAGTGGTGAAGAGCA-3'
Interleukin-1 gi19911214 IL-1 F 5'-GACAGTGAGATGGTGCGATTTC-3'
IL-1 R 5'-ACCATCACTGGCCTGTTGTCT-3' Natural killer enhancing factor gi63002582 NKC F 5'-GACTCACACTTCTCCCATTTTGC-3'
NKC R 5'-TCTTCCTTCAGGACGCCATAG-3'
Heat shock cognate 70 gi39979268 HSC70 F 5'-CCGCACCCAACACCTAAAGT-3'
HSC70 R 5'-CTGTTGCCCTGGTCATTGG-3' Table 2. Primer lists used for real time PCR analysis
contained 10㎕ SYBR Green II real time PCR Master Mix (TaKaRa, Japan), 0.4㎕ of each forward and reverse primer and 2㎕ of cDNA. After denaturing by preincubation for 60 s at 95℃; cDNA was amplified for 40 cycles of denaturation for 15 s at 95℃, annealing for 15 s at 56℃ and extension for 23 s at 72℃. Dissociation- curve analysis was conducted using the ABI real time PCR 7300 Amplification Detection System Software to confirm that there was only one amplified product. All reaction in all cases amplifications were specific and no amplification was observed in negative controls by dissociation-curve analysis. The relative expression of target gene was estimated based on threshold cycle number (Ct). The Ct for each sample was determined by the ABI Sequence Detector Program. Each transcript level was normalized on the basis of the quantification of the HSC70 gene. Relative expressions of all target genes in all cDNAs were calculated by 2-ΔΔCT method (Kenneth and Thomas, 2001). ΔΔCT = (CT,Target-CT,HSC70)Experimental
group-(CT,Target-CT, HSC70)Control group. The results were presented as median with standard deviation. The data were analyzed using the non-parametric Kruskal Wallis test. Differences were considered statistically significant when P < 0.05.
Results
Fatty acid composition of experimental diets The fatty acid compositions of the experimental diets (Table 3) were consistent with the fatty acid compositions in oil sources. SO diet contained a very high concentration (21.9%) of linoleic acid (18:2n-6) while LO diet showed higher concentration of linolenic acid (18:3n-3, 14.6%) than other two diets. Meanwhile, FO diet contained higher concentrations of other n-3 fatty acids such as EPA (20:5n-3) and DHA (22:6n-3) compared to SO or LO diets. Consequently, SO diets contained high proportion of n-6 PUFA while LO and FO diets
Fatty acids 1 Diets
FO LO SO
C14:0 3.1 2.5 2.6
C14:1 0.2 0.2 0.2
C16:0 21.0 18.1 19.9
C16:1 4.7 3.7 3.9
C17:0 0.4 0.5 0.5
C18:0 3.7 3.7 3.8
C18:1n-9 13.2 14.2 14.4 C18:2n-6 12.1 14.6 21.9
C18:3n-3 1.1 13.6 2.4
C18:4n-3 1.3 1.0 1.0
C20:1n-9 2.3 0.8 0.9
C20:3n-3 0.8 0.5 0.5
C20:4n-3 0.4 0.4 0.4
C20:5n-3 14.9 11.6 12.0
C22:2n-6 0.7 0.6 0.6
C22:3n-3 0.3 0.2 0.2
C22:5n-3 3.0 2.5 2.7
C22:6n-3 16.7 11.3 12.0
n-3HUFA2 36.1 26.5 27.8
1 Some minor fatty acids (<0.1 g/100 g fatty acids) are not shown.
2 Highly unsaturated fatty acid (C≥20)
Table 3. Fatty acids composition (% of total fatty acids) of the experimental diets
contained high levels of n-3 PUFA.
Growth and feed utilization
It was observed in this study that different oil formulations of diets did not significantly affect growth performances including weight gain, specific growth rate, feed efficiency and daily feed intake (data not shown).
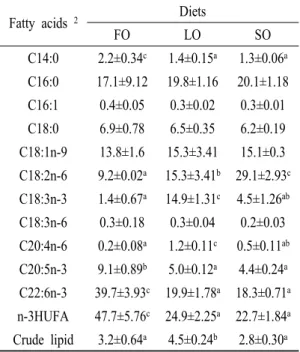
Fatty acids and crude lipid composition
Fatty acid compositions in the muscle were shown
to be strongly influenced by dietary treatments (Table 4) since the muscle of fish fed with the SO diet contained a significantly high concentration (29.1%) of linoleic acid (18:2n-6) while less in LO and FO groups. The muscle of flounder fed with the LO diet had significantly high concentration of linolenic acid (18:3n-3, 13.5%) and FO diet was shown to increase the concentration of other n-3 fatty acids such as EPA (20:5n-3) and DHA (22:6n-3) in muscle. n-3 HUFA was significantly higher in the muscle of flounder fed FO diet. Crude lipid concentration was significantly lower in LO group than other two groups.
Fatty acids 2 Diets
FO LO SO
C14:0 2.2±0.34c 1.4±0.15a 1.3±0.06a C16:0 17.1±9.12 19.8±1.16 20.1±1.18 C16:1 0.4±0.05 0.3±0.02 0.3±0.01 C18:0 6.9±0.78 6.5±0.35 6.2±0.19 C18:1n-9 13.8±1.6 15.3±3.41 15.1±0.3 C18:2n-6 9.2±0.02a 15.3±3.41b 29.1±2.93c C18:3n-3 1.4±0.67a 14.9±1.31c 4.5±1.26ab C18:3n-6 0.3±0.18 0.3±0.04 0.2±0.03 C20:4n-6 0.2±0.08a 1.2±0.11c 0.5±0.11ab C20:5n-3 9.1±0.89b 5.0±0.12a 4.4±0.24a C22:6n-3 39.7±3.93c 19.9±1.78a 18.3±0.71a n-3HUFA 47.7±5.76c 24.9±2.25a 22.7±1.84a Crude lipid 3.2±0.64a 4.5±0.24b 2.8±0.30a
1 Values (mean ± S.E. of four replications) in the same column not sharing a common superscript letter are significantly different (P<0.05).
2 Some minor fatty acids (<0.1 g/100 g fatty acids) are not shown.
Table 4. Fatty acids (% of total fatty acids) and crude lipid composition of dorsal muscle in flounder fed experimental diet for 15 weeks1
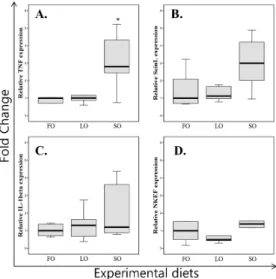
Effect of dietary fat composition on gene expression in head kidney
The relative expression of COS3 (Fig. 1) gene was significantly higher in SO feeding groups. The COS3 mRNA levels of SO were approximately 2-fold higher than mRNA levels of FO and LO. Even though the lowest expression was identified in LO feeding group, there was no statistical difference.
Fig. 1. Real time PCR analysis of COS3 expression in the kidney of the different sources of lipid fed flounder.
FO, fish oil; LO, linseed oil; SO, soybean oil. The gene expression levels were quantified by real time PCR analysis after normalized by HSC70 and expressed as a fold of the FO group. A fold of 1 indicates no difference and a relative expression below 1 indicates down regulation of gene expression compared to FO group.
Significant difference (P<0.05) from the FO group is indicated by an asterisk on top of the bar.
The SO group showed a significantly increased TNF gene expression (Fig. 2A). The ScinL mRNA levels in SO group were approximately 3-fold higher than in FO and LO groups (Fig. 2B). But there was no significantly difference between experimental groups. There are no significantly differences in both gene expression compared with FO group. There was no significant difference in IL-1 (Fig. 2C) and NKEF (Fig. 2D) gene expression among the different experimental groups.
Fig. 2. Real time PCR analysis of TNF (A), ScinL (B), IL-1 (C) and NKEF (D) gene expression in the kidney of the different sources of lipid fed flounder. FO, fish oil;
LO, linseed oil; SO, soybean oil. The gene expression levels were quantified by real time PCR analysis after normalized by HSC70 and expressed as a fold of the FO group. A fold of 1 indicates no difference and a relative expression below 1 indicates down regulation of gene expression compared to FO group. Significant difference (P<0.05) from the FO group is indicated by an asterisk on top of the bar.
Discussion
In this study, it was demonstrated that the composition of dietary fatty acids have influenced the lipid composition of fish body and apoptosis related gene expression in head kidney. Muscle of flounder fed with SO diet was rich in 18:2n-6, whereas those of fish fed with LO diet were rich in 18:3n-3. Muscle of flounder fed with SO and LO diets had lower levels of 20:5n-3 (DHA) and 22:6n-3 (EPA) in comparison to those of flounder fed with FO diet. This is in agreement with other studies in other fishes that the fatty acid composition of muscle reflected the fatty acid composition of the diets (Reinitz and Yu, 1981; Hardy et al., 1987;
Thomassen and Røsjø, 1989; Greene and Selivonchick, 1990; Arzel et al.,1994; Guillou et al., 1995).
High level of apoptosis related gene expression in SO group may be related to the effect of n-6 PUFA on apoptosis. In this study, oxidative stress and damaged cell death were observed by COS3, TNF and ScinL gene expression using real time PCR analysis. COS3 measurements revealed higher expression in the SO groups than in the FO and LO dietary groups. COS3 gene expression can be a marker for oxidative stress within a cell oxidative phosphorylation which is a metabolic pathway that uses energy released by the oxidation of nutrients to produce adenosine triphosphate (ATP). Although oxidative phosphorylation is a vital part of metabolism, it produces reactive oxygen species such as superoxide and hydrogen peroxide. These reactive oxygen species and their reaction products, such as the hydroxyl radical, are very harmful to cells, as they oxidize proteins and cause mutations in DNA (Rattan, 2006; Valko et al., 2007).
Especially n-6 PUFA undergoes peroxidative pathways initiated by hydrogen abstraction followed by oxygen attack on the generated lipid alkyl radical (Cosgrove et al., 1987). Several reports suggest that 18:2n-6 can act as a potent prooxidant in endothelial cells in culture.
For example, 18:2n-6 enhances radical adduct formation in endothelial cells exposed to iron-induced oxidative stress (Alexander-North et al., 1994) and increases peroxisomal -peroxidation (Hennig et al., 1990), a pathway that leads to the production of H2O2. Degradation of 18:2n-6 via peroxidative pathways leads to formation of highly cytotoxic products, such as linoleic acid hydroperoxides (LOOH) or 4-hydroxy-2-(E)-nonenal
(Tamura and Tamura, 1991). LOOH is known as highly cytotoxic lipid peroxide (Kaneko et al., 1987; Kaneko et al., 1988) and also a reactive oxygen species (Matsuo and Kaneko, 2000).
In human, evaluation of some epidemiological studies as well as experimental data linked dietary intake of n-6 PUFA such as linoleic acid to increased risks for cancers of the breast, colon and possibly prostate, particularly in combination with low intake of n-3 PUFA or monounsaturated fatty acids (Bartsch et al., 1999).
In male wistar rats, it was demonstrated that consumption of a high fat diet rich in n-6 PUFA for only 4 weeks instigated mitochondrial nitrosative damage and causes cardiac dysfunction at high after loads (Ghosh et al., 2006). In vitro study, linoleic acid promotes apoptosis and necrosis of Raji cells. The mechanism of cell death induced by these fatty acids seemed to involve mitochondrial depolarization, lipid accumulation, and overexpression of C-MYC and P53 (Maria et al., 2005).
In fish species, the high level of hepatocyte vacuolization and the isolated points of necrosis were observed in liver of gilthead seabream fed with diets containing high levels of n-6 PUFA (Robaina et al., 1998).
The increased activity of TNF and ScinL in the high n-6 PUFA group suggested the induction of apoptosis and increased incidence of cell death. Scinderin like gene (ScinL) is the closest homologue to gelsolin which is a prominent substrate of caspase-3 sequential activation of caspases plays a central role in the execution-phase of cell apoptosis, in vitro (Kothakota et al., 1997; Kamada et al., 1998).
Besides apoptosis inducing effect the linoleic acid (18:2n-6) can be involved in immune and inflammatory
response after elongated to longer n-6 PUFA like arachidonic acid (20:4n-6) that is converted to eicosanoids.
The eicosanoid family acts as inflammatory mediators (prostaglandins, leukotrienes and related metabolites) and through these regulates the intensity and duration of inflammatory and immune responses (Calder, 2002).
Since soybean oil contains high concentration of linoleic acid (Table 3) it can be expected that the proinflammatory gene, especially, IL-1 gene expression would be increased in SO group. However, in this study, there were no significant differences in IL-1 and NKEF gene expressions in SO group. It was also reported previously that there was no significant difference in expression of cytokine genes between the three lipid groups such as fish oil, rapeseed oil, or their 1:1 blend in Atlantic salmon (Seierstad et al., 2009). The reason why high lenoleic acid content in fish diet didn’t affect the inflammatory gene expression in those fish can be explained by the fact that marine fish species, such as flounder (Izquierdo et al., 1992) and turbot (Owen et al., 1975), has little or no ability to desaturate shorter chain PUFAs to longer chain PUFAs. The common eicosanoids are made by oxidation of twenty-carbon essential fatty acids which are synthesized by ∆6 and
∆5 desaturase from the 18:2 n-6 PUFA (Kelley et al., 1998) and some fishes lack or have very little amount of the enzymes.
In summary, we conclude that a high fat diet enriched with n-6 PUFA leads to activation of apoptosis pathways within the cellular damage and without beneficial effects of immune responses. Considering internal factors such as cellular damage and oxidative stress, linseed oil seems to be more suitable lipid source than soybean oil for
replacement of fish oil in flounder.
Acknowledgement
This work was supported by the funds of the Sea Grant Program in Korea.
References
Aksnes, A., Hjertnes, T., Opstvedt, J., 1996. Effect of dietary protein level on growth and carcass composition in Atlantic halibut (Hippoglossus hippoglossus L.). Aquaculture 145, 225-233.
Alexander-North, L.S., North, J.A., Kiminyo, K.P., Buettner, G.R., Spector A.A., 1994. Polyunsaturated fatty acids increase lipid radical formation induced by oxidant stress in endothelial ce1ls.J. Lipid Res 35, 1773-1785.
Arzel, J., Martinez Lopez, F.X., Me’tailler, R., Ste’phan, G., Viau, M., Gandemer, G., Guillaume, J., 1994.
Effect of dietary lipid on growth performance and body composition of brown trout (Salmo trutta) reared in seawater. Aquaculture 123, 361-375.
Barlow, S.M., Pike, I.H., 2001. Aquaculture feed ingredients in year 2010: fish meal and fish oil. In: Nash, C.E., Julien, V. (Eds.), Aquavision ’98. 2nd Nutreco Aquaculture Business Conference, Stavanger, Norway, 13-15 May 1998. Nutreco Aquaculture, Stavanger, Norway 71-74.
Bartsch, H., Nair, J., Owen, R.W., 1999. Dietary polyunsaturated fatty acids and cancers of the breast and colorectum: emerging evidence for their role as risk modifiers. Carcinogenesis 20, 2209-2218.
Budihardjo, I., Oliver, H., Lutter, M., Luo, X., Wang, X., 1999.
Biochemical pathways of caspase activation during apoptosis, Annu. Rev. Cell Dev. Biol 15, 269-290.
Calder, P.C., 2002. Fatty acids and gene expression related to inflammation. Nestle Nutr. Workshop Ser. Clin.
Perform. Program 7, 19-40.
Cosgrove, J.P., Church, D.F., Pryor, W.A., 1987. The kinetics of the autoxidation of polyunsaturated fatty acids.
Lipids 22, 299-304.
Erdal, J.I., Evensen, O., Kaurstad, O.K., Lillehaug, A., Solbakken R., Thorud, K., 1991. Relationship between diet and immune response in Atlantic salmon (Salmo salar L.) after feeding various levels of ascorbic acid and omega-3 fatty acids.
Aquaculture 98, 363-379.
Ghosh, S., Kewalramani, G., Yuen, G., Pulinilkunnil, T., An, D., Innis, S.M., Allard, M.F., Wambolt, R.
B., Qi, D., Abrahani, A., Rodrigues, B., 2006.
Induction of mitochondrial nitrative damage and cardiac dysfunction by chronic provision of dietary omega-6 polyunsaturated fatty acids. Free Radic Biol Med 41, 1413-1424.
Greene, D.H.S., Selivonchick, D.P., 1990. Effects of dietary vegetable, animal and marine lipids on muscle lipid and hematology of rainbow trout (Oncorhynchus mykiss). Aquaculture 89, 165-182.
Guillou, A., Soucy, P., Khalil, M., Adambounou, L., 1995.
Effects of dietary vegetable and marine lipid on growth, muscle fatty acid composition and organoleptic quality of flesh of brook charr (Salvelinus fontinalis). Aquaculture 136, 351-362.
Halliwell, B., Chirico, S., 1993. Lipid peroxidation: its mechanisms, measurement, and significance. Am.
J. Clin. Nutr 57, 715S-725S.
Hardy, R.W., Scott, T.M., Harrell, L.W., 1987. Replacement of herring oil with menhaden oil, soybean oil, or tallow in the diets of Atlantic salmon raised in marine net-pens. Aquaculture 65, 267-277.
Hennig, B., Wang, Y., Boissonneault, G.A., Alvarado, A., Glauert, H. P., 1990. Effects of fatty acids enrichment on the induction of peroxisomal enzymes in cultured porcine endothelial cells.
Biochem. Arch 6, 141-146.
Izquierdo, M.S., Arakawa, T., Takeuchi, T., Haroun, R., Watanabe, T., 1992. Effect of n-3 HUFA levels in Artemia on growth of larval Japanese flounder (Paralichthys olivaceus). Aquaculture 105, 73-82.
Kadenbach, B., Arnold, S., Lee, I., Hüttemann, M., 2004.
The possible role of cytochrome c oxidase in stress-induced apoptosis and degenerative diseases, Biochim. Biophys. Acta 1655, 400-408.
Kamada, S., Kusano, H., Fujita, H., Ohtsu, M., Koya, R.C., Kuzumaki, N., Tsujimoto, Y., 1998. A cloning method for caspase substrates that uses the yeast two-hybrid system: cloning of the antiapoptotic gene gelsolin. Proc Natl Acad Sci USA 95, 8532-8537 Kaneko, T., Honda, S., Nakano, S., Matsuo, M., 1987. Lethal
effects of a linoleic acid hydroperoxide and its autoxidation products, unsaturated aliphatic aldehydes, on human diploid fibroblasts, Chem.-Biol.
Interact 63, 127-137.
Kaneko, T., Kaji, K., Matsuo, M., 1988. Cytotoxicities of a linoleic acid hydroperoxide and its aliphatic aldehydes toward cultured human umbilical vein endothelial cells, Chem.-Biol. Interact 67, 295-304.
Kelley, D.S., Taylor, P.C., Nelson, G.J., Mackey, B.E., 1998.
Arachidonic acid supplementation enhances synthesis of eicosanoids without suppressing immune functions in young healthy men. Lipids 33, 125-30.
Kenneth, J.L., Thomas, D.S., 2001. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-∆∆CT Method. Methods 25, 402-408 Kothakota, S., Azuma, T., Reinhard, C., Klippel, A., Tang,
J., Chu, K., McGarry, T. J., Kirschner, M. W., Koths, K., Kwiatkowski, D. J., Williams, L. T., 1997. Caspase-3-generated fragment of gelsolin:
effector of morphological change in apoptosis.
Science 278, 294-298.
Maria, F. C., Celine, P., Rui, C., 2005. Comparative toxicity of oleic acid and linoleic acid on Raji cells. Nutrition 21, 395-405.
Matsuo, M., Kaneko, T., 2000. The chemistry of reactive oxygen species and related free radicals, in: Z.
Radak (Ed.), Free Radicals in Exercise and Aging, Human Kinetics. Champaign IL, 1-33.
Montero, D., Robaina, L., Caballero, M.J., Ginés, R., Izquierdo, M.S., 2005. Growth, feed utilization and flesh quality of European sea bass (Dicentrarchus labrax) fed diets containing vegetable oils: a time course study on the effect of re-feeding period with a 100% fish oil diet. Aquaculture 248, 121-134.
Mourente, G., Dick, J.R., Bell, J.G., Tocher, D.R., 2005. Effect of partial substitution of dietary fish oil by vegetable oils on desaturation and -oxidation of [1-14C]18:3n-3 (LNA) and [1-14C]20:5n-3 (EPA) in hepatocytes and enterocytes of European sea bass (Dicentrarchus labrax L.). Aquaculture 248, 173-186.
Owen, J.M., Adron, J.W., Middleton, C., Cowey, C.B., 1975.
Elongation and desaturation of dietary fatty acids
in turbot Scophthalmus maximus L. and rainbow trout Salmo gairdnerii Rich. Lipids 10, 528-531.
Pike, I.H., 1999. Health benefits from feeding fish oil and fish meal: The role of long chain omega-3 polyunsaturated fatty acids in animal feeding.
International Fishmeal and Oil Manufacturers Association, Herts.
Pepe, S., 2005. Effect of dietary polyunsaturated fatty acids on age-related changes in cardiac mitochondrial membranes. Exp. Gerontol 40, 751-758 Rattan, S.I., (2006). “Theories of biological aging: genes, proteins,
and free radicals”. Free Radic. Res 40, 1230-1238.
Reinitz, G.L., Yu, T.C., 1981. Effects of dietary lipids on growth and fatty acid composition of rainbow trout (Salmo gairdneri). Aquaculture 35, 19-27.
Robaina, L., Izquierdo, M.S., Moyano, F.J., Socorro, J., Vergara, J.M., Montero., 1998. Increase of the dietary n-3/n-6 fatty accid ratio and addition of phosphorus improves liver histological alterations induced by feeding diets containing soybean meal to gilthead seabream, Sparus aurata. Aquaculture 161, 281-293.
Rodríguez, C., Pérez, J.A., Henderson, J., 2002. The esterification of n-3 and n-6 polyunsaturated fatty acids by hepatocytes and liver microsomes of turbot (Scophthalmus maximus). Comp. Biochem.
Physiol. B 132, 559-570.
Salte, R., Thomassen, M.S., Wold, K., 1988. Do high levels of dietary polyunsaturated fatty acids (EPA/DHA) prevent diseases associated with membrane degeneration in farmed Atlantic salmon at low water temperatures?. Bull. Eur. Assoc. Fish Pathol 8, 63-65.
Saraswathi, V., Wu, G., Toborek, M., Hennig, B. 2004.
Linoleic acid-induced endothelial activation: role
of calcium and peroxynitrite signaling. J. Lipid Res 45, 794-804.
Sargent, J., Bell, G., McEvoy, L., Tocher, D., Estevez, A., 1999. Recent developments in the essential fatty acid nutrition of fish. Aquaculture 177, 191-199.
Seierstad, S.L., Haugland, Ø., Larsen, S., Waagbø, R., Evensen, Ø., 2009. Pro-inflammatory cytokine expression and respiratory burst activity following replacement of fish oil with rapeseed oil in the feed for Atlantic salmon (Salmo salar L).
Aquaculture 289, 212-218.
Tamura, H., and Shibamoto, T., 1991. Gas chromatographic analysis of malonaldehyde and 4hydroxy-2-(E)- nonenal produced from arachidonic acid and linoleic acid in a lipid peroxidation model system.
Lipids 26, 170-173.
Thomassen, M.S., Røsjø, C., 1989. Different fats in feed for salmon: influence on sensory parameters, growth rate, and fatty acids in muscle and heart.
Aquaculture 79, 129-135.
Thompson, K.D., Tatner, M.F., Henderson, R.J., 1996. Effects of dietary (n-3) and (n-6) polyunsaturated fatty acid ratio on the immune response of Atlantic salmon, Salmo salar L. Aquacult. Nutr 2, 21-31.
Toborek, M., Barger, S.W., Mattson, M.P., Barve, S., McClain, C.J., Hennig, B., 1996. Linoleic acid and TNF-alpha cross-amplify oxidative injury and dysfunction of endothelial cells. J. Lipid Res 37, 123-135.
Toborek, M., Lee, Y.W., Garrido, R., Kaiser, S., Hennig, B., 2002. Unsaturated fatty acids selectively induce an inflammatory environment in human endothelial cells. Am. J. Clin. Nutr 75, 119-125.
Tocher, D.R., Bell, J.G., MacGlaughlin, P., McGhee, F.,
Dick, J.R., 2001. Hepatocyte fatty acid desaturation and polyunsaturated fatty acid composition of liver in salmonids: effects of dietary vegetable oil. Comp.
Biochem. Physiol. B 130, 257-270.
Tocher, D.R., Bell, J.G., Dick, J.R., Crampton, V.O., 2003.
Effects of dietary vegetable oil on Atlantic salmon hepatocyte fatty acid desaturation and liver fatty acid compositions. Lipids 38, 723-732.
Torstensen, B.E., Lie, Ø., Frøyland, L., 2000. Lipid metabolism and tissue composition in Atlantic salmon (Salmo salar L.)-effects of capelin-, palm-and oleic acid enriched sunflower oil as dietary lipid sources. Lipids 35, 653-664.
Torstensen, B.E., Froyland, L., Lie, O., 2004. Replacing dietary fish oil with increasing levels of rapeseed oil and olive oil-effects on Atlantic salmon (Salmo salar L.) tissue and lipoprotein lipid composition and lipogenic enzyme activities. Aquac. Nutr 10, 175-192.
Valko, M., Leibfritz, D., Moncol, J., Cronin, M.T., Mazur, M., Telser, J., 2007. “Free radicals and antioxidants in normal physiological functions and human disease”. Int. J. Biochem. Cell Biol 39, 44-84.
Waagbø, R., Sandnes, K., Joergensen, J., Engstad, R., Glette, J., Lie, Ø., 1993. Health aspects of dietary lipid sources and vitamin E in Atlantic salmon (Salmo salar) 2. Spleen and erythrocyte phospholipid fatty acid composition, nonspecific immunity and disease resistance. Fiskeridirektoratets Skrifter.
Serie Ernaering 6, 63-74.
Manuscript Received : October 18, 2012 Revised : November 23, 2012 Accepted : November 26, 2012