International Journal of Oral Biology, Vol. 33, No. 3, September 30 2008, p. 113~116 Copyright ⓒ 2008, The Korean Academy of Oral Biology
113
International Journal of Oral Biology
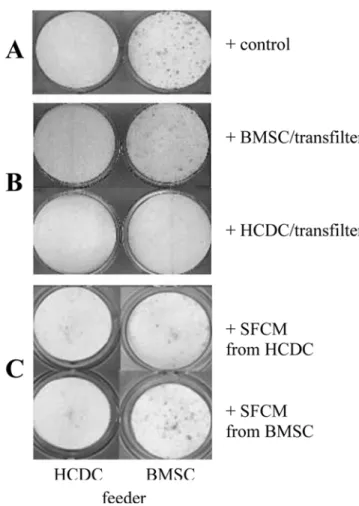
Inhibition of osteoclast formation by putative human cementoblasts
Mi Ri Kim
1, Won Kyung Yang
1, Wojciech Grzesik
2, and Hyun Jung Ko*
11
Department of Dentistry, Asan Medical Center, University of Ulsan.
2