DOI 10.17480/psk.2018.62.2.83
Extracts of Polygonum multiflorum Inhibit RANKL-mediated Osteoclast Differentiation
Kwang-Jin Kim*, Yongjin Lee*, Kyung-Yun Kang**, Yun-Ho Hwang*, Sung-Tae Yee*, Sang-Jip Nam***, and Young-Jin Son*
, #*
Department of Pharmacy, Sunchon National University, Jeonnam, Suncheon 57922, Korea
**
Suncheon Research Center for Natural Medicines, Jeonnam, Suncheon 57922, Korea
***
Department of Chemistry and Nano Science, Global Top 5 Program, Ewha Womans University, Seoul 03760, Korea (Received March 7, 2018; Revised March 30, 2018; Accepted April 13, 2018)
Abstract — Osteoporosis is a disease caused by decreased bone density. The number of patients with osteoporosis is increasing, and the cost of medical treatment is rising because of the increase in fracture patients. Treatment of osteoporosis is important to prevention of bone loss. One treatment for osteoporosis is inhibition of osteoclast formation. Osteoclasts, which are multinucleated cells that absorb bone, are greatly increased and over-activated in bone disorders, including oste- oporosis and rheumatoid arthritis. In this study, we investigated whether extracts of P. multiflorum (PME) influence RANKL-induced osteoclast differentiation. The PME significantly inhibited RANKL-induced osteoclast differentiation by inhibiting transcriptional and translational expression of NFATc1, an essential element of RANKL-mediated osteoclast for- mation. Furthermore, it inhibited the mRNA expression of TRAP, DC-STAMP, and cathepsin K, which are all related to osteoclast differentiation and function. Thus, our results suggest that PME has the potential for use as a functional food and therapeutic substance for the treatment of osteoporosis.
Keywords bone, osteoporosis, osteoclast, RANKL, NFATc1, Polygonum multiflorum
Introduction
Bones are dynamic tissues composed of various types of cells that undergo regenerative and repair processes known as bone remodeling. Osteoclasts and osteoblasts are the main cell types involved in bone modification.
1)It is important to bal- ance the activity of osteoclasts and osteoblasts in bone homeo- stasis. The increased activity or number of osteoclasts in bone homeostasis causes bone diseases such as osteoporosis, Paget's disease, rheumatoid arthritis and periodontal disease.
2)Osteoporosis, which is the most common bone disease world- wide, is associated with decreased bone mass and increased risk of fracture. Osteoporosis patients are more likely to experience fractures, with osteopenia, worsening bone microstructure, and increased bone vulnerability being among the pathological fea-
tures of the disease. The main cause of osteoporosis is related to increased bone resorption because of increased osteoclast number.
2)Osteoclasts are large multinucleated cells formed by the fusion of several mononuclear precursors and the main cells responsible for absorption by the skeleton. Osteoclasts remove old bones and maintain mineral homeostasis.
3,4)Osteoclast dif- ferentiation is regulated by two cytokines, macrophage colony stimulating factor (M-CSF) and receptor activator of nuclear fac- tor- κB (NF-κB) ligand (RANKL).
5)M-CSF produced by immune cells/osteoblasts induces RANK expression and osteoclast sur- vival signals in osteoclast precursor cells.
6)RANKL, which is secreted by osteoblasts/activated T cells, binds to receptor RANK in osteoclasts, after which it induces activation of JNK, ERK, p38, NF-kB, and NFATc1.
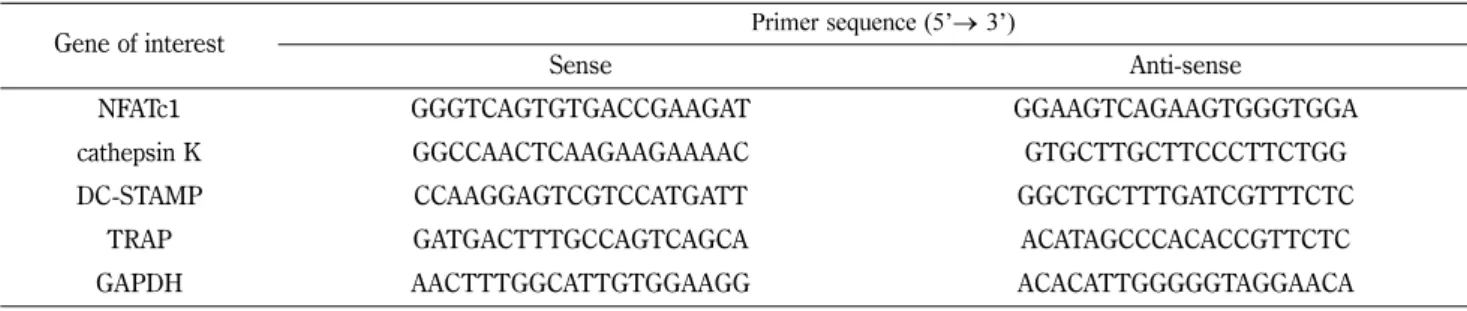
4,5,7-9)NFATc1, which is the key transcription factor for osteoclastogenesis, regulates the expres- sion of osteoclast differentiation and activation factors, such as tartrate-resistant acid phosphatase (TRAP), cathepsin K and dendritic cell-specific transmembrane protein (DC-STAMP).
10-14)Polygonum multiflorum (PM) is one of the most popular Tra- ditional Chinese Medicines and a component of many medi-
#