346 서 론
Minichromosome maintenance proteins (MCM protein)는 효 모에서 인간에 이르기까지 모든 진핵세포에 존재하고 있는 복제단백질로써 MCM2에서 MCM7까지 6종류로 구
성되어 있으며,1) DNA 복제개시와 신장반응에 필수적인 역할을 담당할 뿐 아니라 한 세포주기에서는 단 한 번의 복제만 일어나도록 하는 DNA 복제 Licensing factor로 잘 알려져 있다.1∼3) 세포의 증식은 우선 세포주기의 M기와 G1기에 걸쳐서 6종의 MCM이 heterohexamer 복합체의 형 태로 ORC, CDC6, Cdt1과 함께 chromatin의 복제기점에
산호말 추출물의 복제단백질 MCM의 발현 억제와 항암활성
동의대학교 자연과학대학 생명응용학과, 1분자생물학과, 2생활과학대학 식품영양학과
배신영․이혜림․김광현․한창희1․김영만2․김병우․권현주
Reduction of MCM Protein Expression and Anticancer Activity of the Ethanolic Extracts of Corallina pilulifera
Sin-Yung Bae, Hae-Lim Lee, Kwang-Hyeon Kim, Chang-Hee Han1, Young-Man Kim2, Byung-Woo Kim and Hyun-Ju Kwon
Departments of Life Science and Biotechnology, 1Molecular Biology, College of Natural Science,
2Department of Food and Nutrition, College of Human Ecology, Dongeui University, Busan 614-714, Korea
The MCM protein family has six highly conserved subunits, MCM2 to 7, and is essential for initiation and elongation of DNA replication in all eukaryotes. Deregulation of the MCM function and expression resulted in cells undergoing tumorigenesis. Therefore, these proteins are potential diagnostic markers for cancer and promising targets for anticancer drug development. On the other hand, recently marine organisms are regarded as attractive sources of novel anticancer compounds due to their tremendous biodiversity. In this study, we screened several seaweed for down-regulatory compound of MCM and selected Corallina pilulifera as candidate. Ethanolic extract of Corallina pilulifera (EECP) reduced MCM expression in a dose-dependent manner(0-200μg/ml). EECP showed cytotoxic and antiproliferative activity against six human cancer cells tested such as HeLa, HT29, T24, HepG2, A549 and Jurkat E6-1.
A 30% of tumor mass volume reduction was observed in vivo experiment using mouse bearing Sarcoma 180 cells after treatment of EECP for 14 days, and antitumor efficacy was 72.1%. Oligonucleotide microarray analysis identified 69 up-regulated genes and 79 down-regulated genes in HeLa cells after EECP treatment. The deregulated genes were involved in various biological process including cell cycle regulation and cell proliferation, signal transduction, apoptosis, and predominantly down-regulated genes were mainly associated with cell cycle. (Cancer Prev Res 11, 346-353, 2006)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: MCM protein, Corallina pilulifera, Cytotoxicity, Microarray, Antitumor efficacy
책임저자:권현주, ꂕ 614-714, 부산시 부산진구 엄광로 995 동의대학교 자연과학대학 생명응용학과
Tel: 051-890-1519, Fax: 051-890-1532 E-mail: [email protected]
접수일:2006년 9월 4일, 게재승인일:2006년 9월 29일
Correspondence to:Hyun-Ju Kwon
Department of Life Science and Biotechnology, College of Natural Science, Dongeui University, 995, Eomgwang-no, Busanjin-gu, Busan 614-714, Korea
Tel: +82-51-890-1519, Fax: +82-51-890-1532 E-mail: [email protected]
결합함으로써 복제 전 복합체인 pre-RC를 형성하여 DNA 합성을 하기 위한 준비를 하는데 이를 replication licencing이라 한다.4,5) S기에 진입하면 MCM은 pre-RC의 다른 구성분자들과 함께 인산화효소인 DDK와 S-CDK에 의해 인산화 되어 구조적 변형을 일으키고 여기에 Cdc45 가 결합됨으로써 helicase 기능이 활성화되어 복제기점에 서 DNA의 이중나선을 풀게 되며,6) 이는 PCNA, DNA polymerase 등의 다른 복제단백질들의 결합을 유도하여 DNA의 복제를 시작한다.7,8) 복제 개시 후에도 MCM은 복제분기의 진행에 따라 DNA의 이중나선을 풀어가면서 복제 신장반응에도 도움을 준다.9) 한편 복제가 개시되어 MCM 복합체가 복제기점을 떠나고 나면 남아있는 pre-RC 구성요소들은 S-CDK에 의해 이미 인산화되어 있 으므로 복제기점에 새로운 pre-RC를 형성할 수 없다. 따 라서 이러한 기작에 의해 한 세포주기에서는 단 한번만 복제를 허용하는, 즉 이중 복제가 방지된다.10) 복제가 끝 난 후에는 Cdk2 인산화 작용에 의해 CDC6가 복합체에서 떨어져 나오며 MCM 단백질도 분리되어 복제가 완료된 다. MCM의 이러한 복제 조절기능에 이상이 생기면 세포 는 암화한다.11)
암세포는 분화된 체세포에 비해 고도로 증식하고 있 는 세포이기 때문에 세포 증식 표지인자는 암 진단에 매 우 중요하다.12) 최근의 연구 결과에 의하면 MCM 단백질 은 자궁경부암,13) 신경아세포종,14) 직장암,15) 구강암,16) 유방암17) 전립선암,18) 폐암19) 등 많은 암세포에서 정상세 포보다 과발현 되는 것으로 보고되어 있다. 따라서 MCM 은 기존의 암 진단에 사용되는 Ki67나 PCNA와 같은 증 식 표지인자보다도 훨씬 민감하고 광범위한 암에 적용 할 수 있다는 점과 암의 조기진단이나 악성화를 판단하 는데 중요한 수단으로 사용할 수 있다는 장점 때문에 새 로운 암 진단 표식인자로 부각되고 있으며, 실제로 이를 임상으로 실용화하려는 연구들이 활발히 진행되고 있 다. 게다가 MCM은 세포의 증식에 있어 중요 요소이기 때문에 무한 증식하고 있는 암세포를 타켓으로 한 항암 제 개발의 매우 효과적인 표적으로도 사용할 수 있다.
항암제 개발 전략에 있어서 중요한 기준은 암세포 특이 적이고, 증식 억제력을 가지고 있으며 이러한 활성을 갖 는 물질 탐색에 적절한 측정 방법을 제공할 수 있어야 하는데, MCM은 이러한 기준을 충족시키는 유력한 표적 분자로 인식되고 있다. MCM을 이용한 항암제 개발 방법 은 MCM의 helicase 활성을 저해하는 화합물, MCM 단백 질들의 결합체 형성을 저지하는 화합물, 또는 DDK에 의 한 MCM 인산화를 저지하여 활성형으로의 전환을 억제 하는 화합물 등 MCM의 기능을 저해하는 물질의 개발이
나 MCM의 발현량을 저하시키는 물질의 탐색을 들 수 있다.20)
해양생물은 종의 다양성과 육지 생물과는 다른 생화 학적 다양성 등으로 새로운 생리활성물질의 보고로 인 식되고 있다. 최근 수십 년간 많은 연구자들에 의해 해양 생물로부터 3,000여 종 이상의 신물질이 보고되었고, 이 들 중 몇 가지는 임상응용에 쓰이고 있다.21) 해양생물 중 해조류는 한국을 포함한 아시아에서 전통적으로 식품으 로 이용될 뿐만 아니라 회충구제, 통풍 등의 질병에 민간 요법으로 널리 이용되어 왔다.22) 해조류로부터 유용물질 의 추출에 관한 연구는 여러 연구자들에 의해 수행되어 왔으며 최근 해조류로부터 항암제를 개발하고자 해조류 추출물을 이용하여 인간의 암세포를 대상으로 실험해 암세포의 성장억제나 생체 내 종양억제효과를 가지는 물질들을 분리하여 보고하고 있다.23,24)
따라서 본 연구는 MCM 발현 억제 물질을 분리하여 항암제로의 개발 가능성을 타진하기 위하여 여러 종류 의 해조류를 대상으로 탐색한 결과 작은 구슬 산호말 (Corallina pilulifera)에서 MCM 발현 억제 효과를 확인할 수 있었다. 산호말은 우리나라 연안의 조간대에 널리 분포 되어 있는 홍조류로 적조에 대한 살균활성은 보고된 바 있으나25) 항암활성에 대해 보고된 바는 없다. 이에 본 연 구는 산호말 추출물의 각종 암 세포주에 대한 세포 독성 효과와 세포 증식억제, MCM 단백질의 발현저하 및 발암 을 유도한 마우스에 대한 in vivo 항암활성을 확인하였다.
재료 및 방법 1. 재료의 채취 및 처리
부산 근교 연안의 조간대 바위에서 채취한 작은 구슬 산호말(Corallina pilulifera)을 건조 후 에탄올을 가하여 실 온에서 24시간 냉침추출 하였다. 추가적으로 70oC의 온 도에서 3회 반복 추출하고, 이를 합하여 여과한 다음 감 압농축기로 농축 분말화하고 향후 실험에 사용하였다.
2. 세포주와 배양배지
본 실험에 사용한 세포주는 ATCC (American Type Cul- ture Collection, Rockville)에서 구입하여 사용하였다. 자궁 경부암 세포주인 HeLa, 간암세포주인 HepG2는 DMEM (Dulbecco’s modified Eagle’s medium)에 10% (v/v)의 FBS (Fetal bovine serum)와 0.1% gentamycine을 첨가한 배지를 사용하여 배양하였다. 인간 대장암 세포인 HT-29, 폐암 세포주인 A549, 방광암 세포주인 T-24, 혈액암 세포주인 Jurkat E6-1은 RPMI-1640에 10% (v/v)의 FBS와 0.1% gen-
tamycine을 첨가한 배지를 사용하였으며 37oC, 5% CO2 조 건하에서 배양하였다.
3. 세포독성 및 세포 증식억제 활성의 측정
각각의 종양세포에 대한 산호말 에탄올 추출물(EECP) 의 세포독성을 측정하기 위하여 MTT [3-(4,5-dimethyl- thiazol-2-yl)-2,5-diphenyl tetrazolium bromide] 실험법을 행 하였으며26) 초기 세포수 1×104 cell/ml를 microtiter plate에 분주하여 배양하였다. EECP를 각기 다른 농도(0∼200μg/
ml)로 처리하고, 대조군은 0.1% DMSO로 처리하였다.
37oC에서 24시간 처리한 뒤에 MTT 용액을 첨가하여 37oC 에서 4시간 더 배양한 후 효소면역측정(ELISA, Enzyme linked immunosolvent assay) 판독기를 이용하여 540 nm에 서 측정하였다. 세포증식 억제 활성의 측정은 Trypan blue exclusion assay로 측정하였다.27) 세포를 35 mm dish에 분주하여 각기 다른 농도(0∼250μg/ml) EECP를 처리하 여 24시간 배양 후 세포를 수집하여 trypsin 처리와 PBS 용액으로 세척 후 Trypan blue 용액을 가하여 염색하고 살아있는 세포를 hemacytometer로 측정하였다. 대조군은 0.1% DMSO로 처리하였다.
4. Oligonucleotide microarray analysis
EECP를 처리한 HeLa 세포와 처리하지 않은 대조군 사 이의 유전자 발현 변화 차이를 (주)지노첵에서 제작된 Platinum Biochip Human cancer 3.0 K oligo chip을 사용하여 확인하였다. 총 RNA 표본은 reverse transcription 과정을 통하여 특정한 primer가 부착된 cDNA를 합성하였으며 Platinum oligo chip에 1차 hybridization을 실시하였고, 2차 hybridization 과정 시 labeling 하였다. 본 실험에서 사용된 지노첵 Platinum Biochip Human cancer 3.0K oligo chip은 Qiagen Operon에서 제공하는 Array_Ready Oligo set의 Hu- man cancer oligo set의 알려진 유전자(2,959개), 기능이 알 려지지 않은 EST 유전자(81개)와 housekeeping gene 및 대 조군유전자로 Arabidopsis DNA를 점적한 총 3,096개의 spot이 포함되어 있으며 각 oligonucleotide는 (주)지노첵에 서 제공하는 spotting 용액에 용해시켜 pixsys 5500 arrayer (Cartesian Technologies, CA)를 사용하여 24 Stealth Micro spotting pins로 CMT-GAPS II silane slide glass (Corning, NY) 에 점적하였다. 점적한 슬라이드는 1X SSC 용액으로 1분 간 rehydration하고 UV crosslinker (Stratagene, CA)로 DNA 를 슬라이드에 link시킨 후 succinic anhydride/sodium borate 용액으로 적신 다음 95oC water bath에 2분간 방치하였다.
그 후 슬라이드를 95% 에탄올에 옮겨 1분간 방치한 후 3,000 rpm에서 20초간 원심분리하여 건조시켰다. DNA
chip 분석을 위한 총 RNA의 추출은 EECP를 48시간 처리 한 HeLa 세포와 처리하지 않은 대조 HeLa 세포를 TRI REAGENT (MRC Co. OH)를 사용하여 제조사의 권장법에 따라 추출하였다. 형광 label cDNA probe는 40μg의 총 RNA를 SuperScript II reverse transcriptase를 사용하여 제작 하였다. 역전사 반응액은 400 U SuperScript RNase H-re- verse transcriptase, 0.5 mM dATP, dTTP 와 dGTP, 0.2 mM dCTP와 0.1 mM Cy3 또는 Cy5가 표지된 dCTP를 포함하 였다. 역전사 반응 후 표본 RNA는 5μl의 stop solution (0.5 M NaOH/50 mM EDTA)을 가하여 65oC에서 10분간 반응하여 역전사반응을 정지하였다. 표지된 cDNA 혼합 액은 에탄올 침전법으로 농축하였다. 농축된 Cy3와 Cy5 로 표지된 cDNAs 20μl hybridization solution에 재용해 시 켰다. 두 cDNA를 혼합하여 95oC에서 2분간 변성시킨 후 45oC water chamber에서 20분간 반응시킨 후 cDNA 혼합 액을 점적한 슬라이드를 62oC에서 12시간 hybridization chamber에서 혼성화시켰다. 혼성화된 슬라이드는 2X SSC와 0.1% SDS 용액으로 2분간 세척 후 상온에서 1X SSC 용액으로 3분, 0.2X SSC 용액으로 세척하였다. 그 다 음 슬라이드를 3000rpm에서 20초 원심분리하여 건조하 였다. 혼성화 슬라이드는 Axon Instruments GenePix 4000B 스캐너로 스캔하여 이미지를 enePix Pro 5.1 (Axon, CA)과 GeneSpring 6.1 (Sillicongenetics, CA), R 패키지 프로그램을 사용하여 분석하였다. DNA chip 결과의 분석 과정은 스 캐닝을 통하여 얻어진 spot의 초기 강도(signal intensity) 수치 데이터를 사용하여 전체 표준화(global normalization) 와 강도 의존형 표준화(intensity dependent normalization)를 실시하였다. 또한 chip의 각 block에 대한 차이를 최소화 하기 위하여 block-wise 표준화를 실시하였다.
5. Western blot analysis에 의한 단백질 발현 분석
HeLa 세포를 CSK 완충액(10mM Pipes, pH 6.8, 100 mM NaCl, 1 mM MgCl2, 1 mM EGTA, 1 mM dithiothreitol, 1 mM phenylmethanesulfonyl fluoride)에 0.1% Triton X-100, 1 mM ATP 와 단백질 분해효소 저해제(Pharminogen A)가 첨가된 용액을 사용하여 현탁한 후 초음파 분해기로 파 쇄 하였다. 파쇄된 세포는 15,000×g로 30분간 원심 분리 하였으며, 상층액의 단백질 농도는 BCA assay kit를 사용 하여 측정하였다. SDS-PAGE 시에는 BCA 방법으로 정량 한 동량의 단백질(20μg)을 사용하였고, 10%의 분리 gel 을 사용하였다. 단백질 전기영동 후 gel 내의 단백질을 PVDF membrane에 옮긴 후 blocking buffer를 사용하여 실 온에서 blocking과정을 거쳤다. 1차 항체는 MCM2 (Santa- Cruz, sc-9839), MCM3 (SantaCruz, sc-9850), MCM4 (BD Phar-
mingen, 559544), MCM6 (SantaCruz, sc-9843), MCM7 (Santa- Cruz, sc-9966), actin (SantaCruz, sc-1616)을 사용하여 37oC에 서 1시간 반응하였다. 1차 항체를 제거한 후 peroxidase가 결합된 이차 항체를 사용하여 37oC에서 반응시키고 면 역반응 단백질은 화학발광시스템(chemiluminescence system;
SuperSignal West Femto Maximum sensitivity Substrate, Pierce Co.)으로 검출하였다. 정량은 FluorchemTM 5500 (Alpha Innotech)을 사용하였다.
6. In vivo 항암 활성 측정
산호말 추출물의 항암 활성 측정을 위해 동물 모델로 는 체중 20∼24 g, 6∼8주령의 C3H/HeNTacSam-MTV 웅 성 마우스를 샘타코(Samtako Bio, Inc., Osan, Korea)에서 주문하여 사용하였다. 동물실험과 모든 과정에 관한 것 은 규격화된 지침(D.L. No. 116, G.U., Suppl. 40, Feb. 18, 1992; Circolare No. 8, G.U., July 1994)과 국제법 (EEC Council Directive 86/609, OJ L 358. 1, Dec 12, 1987; Guide for the Care and Use of Laboratory Animals, United States National Research Council, 1996)에 따라 행하였다. 1주일의 적응기간을 가진 후, 계대 배양한 Sarcoma-180 세포를 1
×106 세포수가 되도록 PBS로 희석하여 피하에 주사하 여 암을 유발시켰으며 그룹당 5마리씩 4개 그룹으로 구 성하여 실험을 수행하였다. 암세포 주사 후에 10∼14일 에 걸쳐 복강 팽만 및 털의 팽윤 등 암의 징후를 보이는 마우스를 선별하여 음성대조군은 10% tween 80을 100μl 투여하였다. 양성대조군은 항암제인 doxorubicin 1 mg/kg, 실험군은 EECP 10 mg/kg을 각각 경구 투여하였다. Doxo- rubicin은 Sigma (Sigma Chemical Co.)에서 구입하였으며 0.9%의 생리식염수에 녹여서 사용하였으며 EECP는 10%
tween 80용액에 현탁하여 사용하였다. 약물 투여 후 15일 째에 마우스를 희생시키고 해부하여 실험군과 대조군의 복강 내에 발생한 종양을 적출하여 무게와 크기를 측정 하였다. 종양의 크기(V)는 다음 식 V=(width×depth×
length)/2에 대입하여 계산하였다. 항암효율(TVI%)은 백 분율로 나타내었으며 다음 식 항암 효율=[1-(EECP 처 리군의 종양 부피 평균값/대조군의 종양 부피 평균값)]
×100에 따라 계산하여 나타내었다.
결과 및 고찰
1. 산호말 추출물 EECP의 MCM 발현 저하 효과
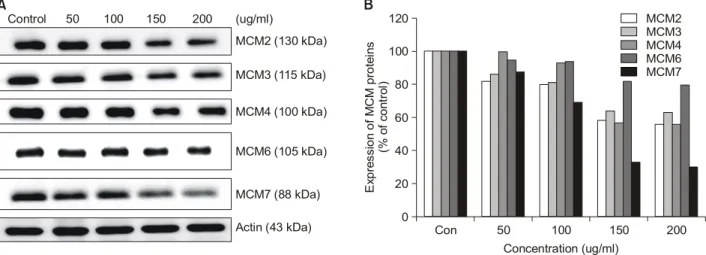
MCM 단백질은 진핵세포의 복제 개시와 genome의 중 복복제 방지 기능을 가지고 있기 때문에 진핵생물의 ge- nome의 안정한 복제에 중요한 역할을 담당하고 있다.1∼3) 또한 HeLa 세포나 사람 자궁 경부 암 조직세포 등에 있 어 MCM의 발현이 정상세포와 비교해 과발현 되고 있다 는 사실이 보고되어,13) MCM이 새로운 항암제 개발의 표 적 분자로 주목받고 있다. 따라서 해조류에서 MCM 단백 질의 발현을 저하시키는 물질을 찾기 위하여 탐색한 결 과 산호말 에탄올 추출물(EECP)에서 이와 같은 활성을 확인할 수 있었다(Fig. 1). EECP를 0∼200μg/ml 처리한 HeLa 세포에서 MCM2, 3, 4, 6, 7 단백질의 발현이 농도 의존적으로 감소하는 것을 확인하였다. MCM 단백질들 간의 발현의 차이는 MCM7이 가장 큰 발현 변화를 보여 200μg/ml의 EECP를 처리했을 때 대조군에 비해 69.8%
의 발현 감소를 보였으며, 변화가 가장 적은 것은 MCM6 이었다. MCM 유전자는 몇 개의 E2F 결합 부위를 pro- moter 영역에 가지고 있으며 이들 유전자의 전사는 E2F
Fig. 1. Effects of EECP on MCM expression in HeLa cells. (A) Western blot analysis (B) relative expression profile.
AControl 50 100 150 200
MCM2 (130 kDa)
MCM3 (115 kDa)
MCM4 (100 kDa)
MCM6 (105 kDa)
MCM7 (88 kDa) (ug/ml)
Actin (43 kDa) Con 50 100 150 200
Expression of MCM proteins (% of control)
Concentration (ug/ml) 0
20 40 60 80 100
B 120
MCM3 MCM2
MCM6 MCM4 MCM7
전사인자에 의해서 조절되는 것으로 보고되어 있다.28∼30) HeLa 세포는 papilloma E7 oncogene의 생산물이 hyperphos- phorylated Rb에 결합하여 Rb/E2F결합체를 불안정화시키 며 이에 따라 E2F에 의해 발현조절 되는 것으로 보고되 어 있다.31) 따라서 EECP의 MCM 발현저하 효과는 E2F에 의한 전사 레벨에서의 발현량 감소로 추정할 수 있으며 향후 이에 대한 자세한 연구가 필요하다.
2. 인간 암 세포별 세포독성 및 증식억제 효과
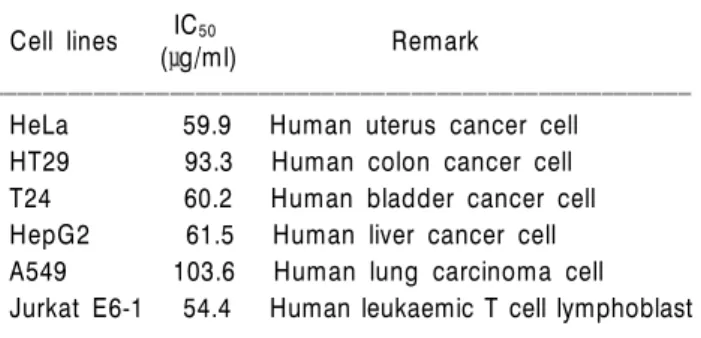
MCM 발현 감소 물질인 EECP의 암 세포에 대한 세포독 성을 알아보기 위해 6종의 인간 암세포에 추출물을 처리 하여 그 효과를 검토해 보았다. 사용한 세포는 자궁경부 암 세포(HeLa), 대장암세포(HT29), 방광암세포(T24), 간암 세포(HepG2), 폐암세포(A549), 혈액암세포(Jurkat E6-1)를 사용하였으며 암 세포별 성장 억제 정도를 MTT법으로 측정하여 그 IC50을 Table 1에 나타내었다. IC50은 Jurkat E6-1, HeLa, T24, HepG2 세포에서 각각 54.4, 59.9. 60.2, 61.5μg/ml로 비슷한 세포독성을 나타내었으며 HT29와
A549 세포에는 93.3, 103.6μg/ml로 다른 세포에 비해 낮 은 세포 독성을 나타내었다. 각 세포에 대한 증식억제 효 과를 검토하기 위하여 EECP를 농도별(5, 10, 15, 30, 60, 120 and 250μg/ml)로 24시간 처리하여 Trypan blue assay로 생존율을 측정하였다. 대조군은 0.1% DMSO를 사용하였 고, 생존율은 대조군 대비 생존율 백분율로 계산하였다.
모든 세포에서 농도 의존적으로 증식억제 효과가 관찰되 었으며, 특히 HeLa 세포와 HepG2 세포에 대한 증식억제 효과가 뛰어나 30μg/ml의 EECP를 24시간 처리하였을 때 HeLa 세포와 HepG2 세포는 70% 정도의 생존율을 나타내 었으며 60μg/ml의 농도 이후부터는 생존율이 급격하게 줄어들어 120μg/ml에서는 약 20%의 생존율을 나타내었 다(Fig. 2).
3. In vivo 항암활성
EECP의 in vivo 항암 활성을 측정하기 위하여 Sarcoma 180 세포를 마우스의 피하에 주사하여 종양을 형성시켰 다. EECP는 10 mg/kg의 양으로, doxorubicin은 1 mg/kg의
Table 1. Cytotoxic activity of EECP on human cancer cell lines ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
IC50
Cell lines (μg/ml) Remark
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ HeLa 59.9 Human uterus cancer cell
HT29 93.3 Human colon cancer cell T24 60.2 Human bladder cancer cell HepG2 61.5 Human liver cancer cell A549 103.6 Human lung carcinoma cell Jurkat E6-1 54.4 Human leukaemic T cell lymphoblast ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Fig. 2. Dose-dependent growth inhibitory effects of EECP on cancer cell lines.
Fig. 3. Inhibition of tumor growth in C3H/HeNTacSam-MTVC mouse bearing Sarcoma 180 cancer cells. EECP was admin- istered orally every day on day 0 to day 14 at doses of 10 mg/
kg. The figure shows the relative tumor sizes. (A) is tumor in control mice (B) and (C) are tumors in mice of doxorubicin and EECP administration, respectively.
A B C
Table 2. Antitumoric effect of EECP in Sarcoma 180 bearing tumor mouse
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Control Doxorubicin EECP
(n=5) (n=4) (n=4)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Body weight (g) 31.2±4.8 29.1±0.6 27.6±0.32 Tumor volume (mm3) 445.5±88.4 173.5±81.4a121.8±13.9a Tumor mass (mg) 385±56 158±67a 154±23a Antitumor efficacy (%) 0 61.1 72.1 ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
aCompared with control group, p<0.05.
양으로 2주간 경구 투여하였다. 2주 경과 후 실험군과 대 조군에서 종양을 적출하여 각각의 종양 크기를 측정해 보았다. 그 결과 EECP 처리군과 doxorubicin 처리군의 종 양의 크기와 무게는 대조군에 비해 현저히 작은 것을 확 인할 수 있었다(Fig. 3). 종양의 크기와 무게는 EECP 및 doxorubicin을 처리하였을 때 변화가 뚜렷하였으며(Table 2) 항암효율은 EECP가 72.1%, doxorubicin이 61.1%로 나타나 양성대조군인 상용 항암제인 doxorubicin보다 항암 효과 가 약간 나은 것으로 나타났다. 이와 같은 결과로 보아 EECP는 MCM 복제 단백질의 발현을 억제하여 암세포의 증식을 억제하는 새로운 기작의 항암제 개발의 후보 물 질로서 이용될 수 있는 가능성이 매우 높은 물질로 기대 된다.
4. Microarray
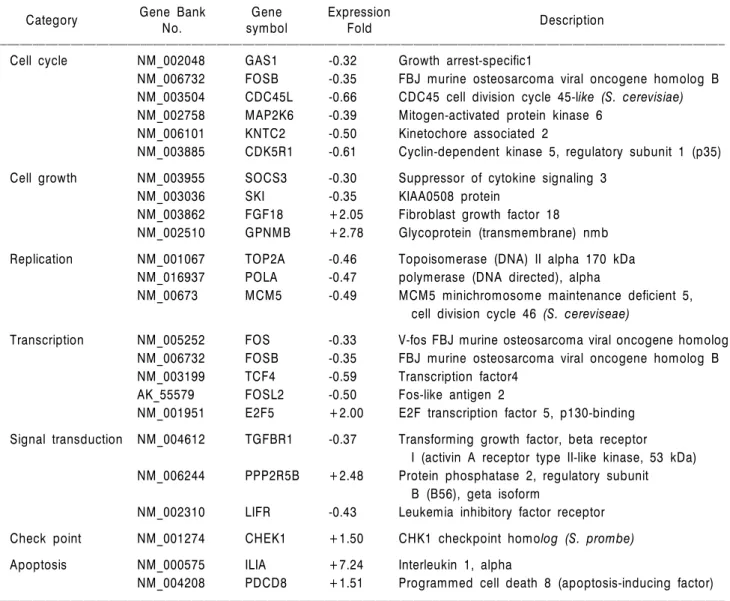
EECP는 진핵세포의 중요한 복제개시인자인 MCM 단 백질의 발현을 저하시킴과 동시에 암세포에 대한 세포 독성과 증식 억제 효과 및 in vivo 항암 활성을 나타내었 다. EECP의 MCM 단백질 발현 저하 활성 이외의 다른 항암 관련 유전자의 발현 변화를 검토하기 위하여 (주)지 노첵에서 제작된 Platinum Biochip Human cancer 3.0K oligo chip을 사용하여 실험하였다. 발현량의 차이를 나타 내는 유의성은 EECP 처리군과 대조군의 2배 이상의 발 현량의 차이를 보이는 유전자로 분류하였다. Microarray 분석 결과 유전자 발현변이는 IL1A gene을 포함해서 총 69개의 유전자가 2배 이상 과발현 하였으며, 반면 2배
Table 3. List of up to down-regulated genes by EECP in HeLa cell
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Gene Bank Gene Expression
Category No. symbol Fold Description
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Cell cycle NM_002048 GAS1 -0.32 Growth arrest-specific1
NM_006732 FOSB -0.35 FBJ murine osteosarcoma viral oncogene homolog B NM_003504 CDC45L -0.66 CDC45 cell division cycle 45-like (S. cerevisiae) NM_002758 MAP2K6 -0.39 Mitogen-activated protein kinase 6
NM_006101 KNTC2 -0.50 Kinetochore associated 2
NM_003885 CDK5R1 -0.61 Cyclin-dependent kinase 5, regulatory subunit 1 (p35) Cell growth NM_003955 SOCS3 -0.30 Suppressor of cytokine signaling 3
NM_003036 SKI -0.35 KIAA0508 protein
NM_003862 FGF18 +2.05 Fibroblast growth factor 18
NM_002510 GPNMB +2.78 Glycoprotein (transmembrane) nmb Replication NM_001067 TOP2A -0.46 Topoisomerase (DNA) II alpha 170 kDa
NM_016937 POLA -0.47 polymerase (DNA directed), alpha
NM_00673 MCM5 -0.49 MCM5 minichromosome maintenance deficient 5, cell division cycle 46 (S. cereviseae)
Transcription NM_005252 FOS -0.33 V-fos FBJ murine osteosarcoma viral oncogene homolog NM_006732 FOSB -0.35 FBJ murine osteosarcoma viral oncogene homolog B
NM_003199 TCF4 -0.59 Transcription factor4
AK_55579 FOSL2 -0.50 Fos-like antigen 2
NM_001951 E2F5 +2.00 E2F transcription factor 5, p130-binding Signal transduction NM_004612 TGFBR1 -0.37 Transforming growth factor, beta receptor
I (activin A receptor type II-like kinase, 53 kDa) NM_006244 PPP2R5B +2.48 Protein phosphatase 2, regulatory subunit
B (B56), geta isoform
NM_002310 LIFR -0.43 Leukemia inhibitory factor receptor Check point NM_001274 CHEK1 +1.50 CHK1 checkpoint homolog (S. prombe) Apoptosis NM_000575 ILIA +7.24 Interleukin 1, alpha
NM_004208 PDCD8 +1.51 Programmed cell death 8 (apoptosis-inducing factor) ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
이상 저발현 한 유전자는 AKAP12 유전자를 포함한 79개 였다(data was not shown). 발현 변이 유전자는 세포주기, 세포증식, DNA 복제와 전사, 신호전달, apoptosis에 관련 한 유전자들로 그 중 EECP에 의해 발현이 증감되는 중요 유전자들을 정리하여 Table 3에 나타내었다. 암에 관련 한 유전자 중 복제에 관계하는 유전자 중에는 topoi- somerase 유전자인 TOP2A가 2배 이상(-0.46) 감소하였음 을 확인하였다. 염색체의 DNA는 아주 조밀하게 꼬여있 으며 이것을 superhelix라고 하는데, topoisomerase가 DNA 복제 시 꼬여있는 DNA를 풀어주는 작용을 담당한다.
Topoisomerase는 생물체에서 DNA 복제 시 반드시 필요한 효소 중 하나이기 때문에 항암제 개발의 주요 표적 단백 질로 인식되어 최근 이 효소에 대해 많은 연구가 이루어 지고 있으며32) EECP가 이 효소의 발현을 억제한다는 사 실은 추출물 내에 발현억제 물질이 있을 가능성을 내포 하고 있어서 매우 흥미로운 결과로 사료되며 현재 EECP 내에서 이 물질을 순수 분리하려는 연구를 진행 중에 있 다. 그 외에도 최근 새로운 항암제 개발 표적 분자로 주 목받고 있는 CDC45 (-0.66), CDK5 (-0.61)의 감소도 확 인되었으며, 본 연구의 중요 표적 분자인 MCM5 (-0.49) 의 감소도 확인하였다. 전체적으로는 세포주기 조절 유 전자들인 GAS1, FOSB CDC45, MAP2K, KNTC2, CDK5 등 의 유전자 발현 저하가 현저하여 산호말 추출물의 항암 활성은 세포주기조절에 의한 것으로 생각되며, 이에 대 한 정확한 기작을 규명하기 위해 현재 이들 분자들에 대 한 발현량의 변화를 Western blot과 Real Time PCR로 확인 중에 있다.
결 론
Minichromosome maintenance protein (MCM protein)은 MCM2-7까지 6종의 유사 subunit로 구성된 복제단백질로 진핵세포의 복제단계에서 복제개시와 신장에 매우 중요 한 역할을 한다. MCM의 기능 이상이나 발현조절 실패로 세포는 암화한다. 따라서 MCM은 암 진단이나 항암제 개 발의 중요 표적으로 인식되고 있다. 한편 최근에는 해양 생물이 그 종의 다양성으로 인해 새로운 항암제 개발의 중요 자원으로 각광받고 있다. 본 연구에서는 여러 가지 해조류를 탐색한 결과 작은 구슬 산호말 에탄올 추출물 (EECP)이 농도 의존적(0∼200μg/ml)으로 MCM 발현 억 제 활성을 가지는 것을 확인하였다. EECP는 자궁경부암 세포(HeLa), 대장암세포(HT29), 방광암세포(T24), 간암세 포(HepG2), 폐암세포(A549), 혈액암세포(Jurkat E6-1) 등 실험한 6종의 인간 암세포에 세포독성을 나타내었으며,
세포의 증식 억제 효과도 확인되었다. Sarcoma 180 세포 로 암화된 mouse를 이용한 in vivo 실험에서 EECP를 10 mg/kg씩 14일간 투여하였을 때 종양 크기가 30% 이하로 감소하는 것을 확인하였으며 항암 효율은 72.1%였다. 종 양 관련 유전자의 발현에 미치는 영향을 조사하기 위하 여 자궁경부암 세포인 Hela 세포를 이용한 Microarray 분 석 결과 과발현 유전자 69개, 저발현 유전자 79개로 주로 세포주기관련 유전자의 발현저하를 확인할 수 있었다.
감사의 글
본 연구는 해양수산부 마린바이오21사업의 해양바이 오프로세스연구단 연구비지원에 의해 수행되었습니다.
참 고 문 헌
1) Tye BK. MCM proteins in DNA replication. Annu Rev Biochem 68, 649-686, 1999.
2) Lei M, Tye BK. Initiating DNA synthesis: from recruiting to activating the MCM complex. J Cell Sci 114, 1447-1454, 2001.
3) Diffley JF, Labib K. The chromosome replication cycle. J Cell Sci 115, 869-872 2002.
4) Bell SP, Stillman B. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature 357, 128-134, 1992.
5) Diffley JF, Cocker JH, Dowell SJ, Rowley A. Two steps in the assembly of complexes at yeast replication origins in vivo.
Cell 78: 303-316, 1994.
6) Masuda T, Mimura S, Takisawa H. CDK- and Cdc45- dependent priming of the MCM complex on chromatin during S-phase in Xenopus egg extracts: possible activation of MCM helicase by association with Cdc45. Genes Cells 8, 145-161, 2003.
7) Aparicio OM, Stout AM, Bell SP. Differential assembly of Cdc45p and DNA polymerases at early and late origins of DNA replication. Proc Natl Acad Sci USA 96, 9130-9135, 1999.
8) Walter J, Newport J. Initiation of eukaryotic DNA repli- cation: origin unwinding and sequential chromatin association of Cdc45, RPA, and DNA polymerase α. Mol Cell 5, 617- 627, 2000.
9) Labib K, Tercero JA, Diffffley JF. Uninterrupted MCM2-7 function required for DNA replication fork progression. Science 288, 1643-1647, 2000.
10) Diffley JF, Cocker JH, Dowell SJ, Rowley A. Two steps in the assembly of complexes at yeast replication origins in vivo.
Cell 78, 303-316, 1994.
11) Gonzalez MA, Tachibana KE, Laskey RA, Coleman N.
Control of DNA replication and its potential clinical ex-
ploitation. Nat Rev Cancer 5, 135-141, 2005.
12) Davis RL, Onda K, Shubuya M, Lamborn K, Hoshino T.
Proliferation markers in gliomas: a comparison of BUDR, Ki-67, and MIB-1. J Neurooncol 24, 9-12, 1995.
13) Ishimi Y, Okayasu I, Kato C, Kwon HJ, Kimura H, Yamada K, Song SY. Enhanced expression of MCM proteins in cancer cells derived from uterine cervix. Eur J Biochem 270, 1089- 1101, 2003.
14) Shohet JM, Hicks MJ, Plon SE, Burlingame SM, Stuart S, Chen SY. Minichromosome maintenance protein MCM7 is a direct target of the MYCN transcription factor in neur- oblastoma. Cancer Res 62, 1123-1128, 2002.
15) Yoshida K, Inoue I. Conditional expression of MCM7 increases tumor growth without altering DNA replication activity. Fed Eur Biochem Soc Lett 553, 213-217, 2003.
16) Kodani M, Osaki K, Araki SK, Goto E, Ryoke K, Ito H.
Minichromosome maintenance 2 expression is correlated with mode of invasion and prognosis in oral squamous cell car- cinomas. J Oral Pathol Med 32, 468-474, 2003.
17) Yu K, Lee CH, Tan PH, Hong GS, Wee SB, Wong CY, Tan P. A molecular signature of the Nottingham prognostic index in breast cancer. Cancer Res 64, 2962-2968, 2004.
18) Meng, MV, Grossfeld GD, Williams GH, Dilworth S, Stoeber K, Mulley TW, Weinberg V, Carroll PR, Tisty TD. Mini- chromosome maintenance protein 2 expression in prostate:
characterization and association with outcome after therapy for cancer. Clin cancer Res 7, 2712-2718, 2001.
19) Ramnath N, Hernandez FJ, Tan DF, Huberman JA, Nata- rajan N, Beck AF, Hyland A, Todorov IT, Brooks JS, Bepler G. MCM2 is an independent predicter of survival in patients with non-small-cell-lung cancer. J Clin Oncol 19, 4259-4266, 2001.
20) Lei M. The complex: its role in DNA replication and impli- cations for cancer therapy. Current Cancer Drug Targets 5, 365- 380, 2005.
21) Cragg GM, Newman DJ, Weiss RB. Oral reefs, and thermal vents: the worldwide exploration of nature for novel antitumor agents. Semin Oncol 24, 156-163, 1997.
22) Michanek G. Seaweed resources for pharmaceutical uses, In:
eds, by Hoppe HA, Levring T, Tanaka Y, Welter de Gruyter Marine algae in Pharmaceutical Science, Berlin, New York, pp 203-234, 1979.
23) Furusawa E, Furusawa S. Anticancer activity of a natural product, viva-natural, extracted from Undaria pinnantifida on intraperitoneally implanted Lewis lung carcinoma. Oncology 42, 364-369, 1985.
24) Deslande E, Pondaven P, Auperin T, Roussakis C, Guezennec J, Stiger V, Payri C. Preliminary study of the in vitro anti- proliferative effect of a hydroethanolic extract from the sub- tropical seaweed Turbinaria ornata (Turner J. Argadh) on a human non-small-cell bronchopulmonary carcinoma cell line (NSCLC-N6). J Appl Phycol 12, 257-262, 2000.
25) Jeong JH, Jin HJ, Sohn CH, Suh KH, Hong YK. Algicidal activity of the seaweed Corallina pilulifera against red tide microalgae. J Appl Phycol 12, 37-43, 2000.
26) Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immu Methods 65, 55-63, 1983.
27) Jones KH, Senf JA. An improved method to determine cell viability by simultaneous staining with fluorescein diacetate- propidium iodide. J Histochem Cytochem 33, 77-79, 1985.
28) Tsuruga H, Yabuta N, Hosoya S, Tamura K, Endo Y, Nojima H. HsMCM6: a new member of the human MCM/P1 family encodes a protein homologous to fission yeast Mis5. Genes Cells 2, 381-399, 1997.
29) Suzuki S, Adachi A, Hiraiwa A, Ohashi M, Ishibashi M, Kiyono T. Cloning and characterization of human MCM7 promoter. Gene 216, 85-91, 1998.
30) Ohtani K, Iwanagam R, Nakamura M, Ikeda M, Yabita N, Tsuruga H, Nojima H, Cell growth-regulated expression of mammalian MCM5 and MCM6 genes mediated by the transcription factor E2F. Oncogene 18: 2299-2309, 1999.
31) Goodwin EC, DiMaio D. Repression of human papillomavirus oncogenes in HeLa cervical carcinoma cells causes the orderly reactivation of dormant tumor suppressor pathways. Proc Natl Acad Sci USA 97, 12513-12518, 2000.
32) Dutta R. Inouye GHKL, an emergent ATPase/kinase super- family. Trends Biochem Sci 25, 24-28, 2000.