1. Introduction
Ginseng radix, the root of Panax ginseng C. A. Meyer (Araliaceae), is one of the most
famous Oriental medical herbs and has several therapeutic applications. It is well documented that Ginseng radix possesses a number of pharmacological effects including hypotensive, cardiotonic, sedative, aphrodisiac, anti-aging, and anti-oxidant actions
1-3).
It was reported that exposure to prenatal
▪교신저자 : 김이화, 충북 제천시 신월동 세명대학교 한의과대학 경락경 혈학 교실, Tel. 043-649-1348, Fax. 043-652-1348, E-mail : [email protected]
이 논문은 2004년도 한국학술진흥재단의 지원에 의하여 연구되었음 (KRF-2004-002-E00188).
⋅투고 : 2006/11/07 ⋅심사 : 2006/12/01 ⋅채택 : 2006/12/06
인삼약침이 소음스트레스로 인한 태아쥐의 NOS 신경세포 발현에 미치는 영향
이익재1⋅임강현2⋅서일복3⋅구성태4⋅최선미4⋅김이화1
세명대학교 1한의과대학 경락⋅경혈학교실, 2한의과대학 본초학교실,
3한의과대학 해부학교실, 4한국한의학연구원 의료연구부
Effect of Ginseng radix herb-acupuncture on noise stress-induced NOS expression in the offspring rats
Ek-Jae Lee
1, Kang-Hyun Lem
2, Il-Beok Seo
3, Sung-Tae Koo
4, Sun-Mi Choi
4, Ee-Hwa Kim
1Dept. of 1Meridian & Acupoint, 2Herborogy, 3Anatomy, College of Oriental Medicine, Semyung University
4Dept. of Medical Research, Korea Institute of Oriental Medicine
Abstract
목적 : 인삼은 전통적으로 학습과 기억능력 증진 및 각종 질병치료에 우수한 효과가 있는 것으로 알려져 있다. 본 연구에서는 이러한 인삼약침이 소음스트레스가 유발된 태아쥐의 해마 및 시상하부에서 nitric ox- ide synthase에 미치는 영향을 관찰하였다.
방법 : 인삼약침이 소음스트레스에 미치는 영향을 연구하기 위하여 태아기에 소음스트레스를 유발하고 생후 4주후 7일간 인삼약침을 투여한 후 NADPH-d 조직화학법을 시행하였다.
결과 : 소음스트레스는 태아쥐의 해마 및 시상하부에서 NOS 발현이 유의하게 증가하였다. 2. 인삼약침 은 해마 CA1에서만 소음스트레스에 의해서 증가된 NOS 발현을 유의하게 억제하였다. 3. 인삼약침은 시상 하부 PVN, DMH 및 LHA에서 소음스트레스에 의해서 증가된 NOS 발현을 유의하게 억제하였다.
결론 : 본 연구를 통하여 인삼약침이 소음스트레스가 유발된 태아쥐의 해마 및 시상하부에서 증가된 NOS 발현을 유의하게 억제시켜 태아의 스트레스 자극에 인삼약침치료가 유의한 효과가 있음을 확인하였 다.
Key words : Ginseng radix; prenatal noise stress; nitric oxide synthase; hippocampus; hypothalamus
T h e K orea n Jou rn al of M erid ia n & A cu p oin t
stress alters an individual's developmental trajectory through altered early brain development. Various physical and emotional stresses applied during pregnancy result in low birth-weight of the offspring, increased risk of premature delivery, and a higher in- cidence of neonatal abnormality
4). Delayed motor and cognition development was also observed in the offspring of stressed pregnant rats
5). In late gestation, the fetus can hear sound from the outside of the mother
6). Previous studies reported that exposure to noise during pregnancy adversely influenced the development of the fetus and neonate: in- creased antepartum fetal death and congenital anomaly in the central nervous system, im- paired social behavior in juvenile stage, and a long-term alteration in the immune function
7,8). In addition, stress is associated with activa- tion of the hypothalamic-pituitary-adrenal (HPA) axis. Various stress related inputs converge upon the neurons located in the par- aventricular nucleus (PVN). In fact, this nu- cleus has a pivotal role in the control of pitui- tary-adrenocortical activity in response to stress.
The hippocampal formation is a brain re- gion critically involved in learning and memo- ry formation. In humans, hippocampal damage impairs explicit memory, and in rodents, hip- pocampal damage impairs spatial and con- textual learning which require the formation of relational representations among multiple cues. In various studies, prenatal stress by re- straint
9), alcohol
10)and noise
11)is known to in-
fluence learning and memory capabilities of the offspring by altering neuronal activity in the hippocampus and related structures
12).
Nitric oxide (NO), endogenously gen- erated from L-arginine by NO synthase (NOS), is a free radical with signaling func- tions in the central nervous system (CNS). It has been known to play important roles im- plicated in numerous physiological and patho- logical processes in the brain
13). Nicotinamide adenine dinucleotide phosphate-diaphorase (NADPH-d) is a histochemical marker specific for NOS in the CNS. Neurons containing NADPH-d have been reported to be relatively resistant to various toxic insults and neuro- degenerative disorders.
In the present study, the influence of postnatal Ginseng radix herb-acupuncture on neuronal development, in particular with re- spect to NOS expression in the each regions of hippocampus and hypothalamus of offspring rats with prenatal noise stress during preg- nancy was investigated via NADPH-d histo- chemistry.
2. Materials & Methods
2.1. Animals and treatments
The experimental procedures were per-
formed in accordance with the guidelines of
the National Institute of Health (NIH) and the
Korean Academy of Medical Sciences. Male
Sprague-Dawley rats (250 ± 10 g, 12 weeks
old) and female Sprague-Dawley rats (180 ±
10g, 8 weeks old) were used in this study.
Female rats (n = 40) were allowed to mate with male rats (n = 40) for 24 h. One day lat- er, female rats were separated from the male rats and housed individually in a plastic home cage at the controlled temperature (20 ± 2℃) and the light-dark cycle of 12 h of light and 12 h of darkness (light on from 07:00 h to 19:00 h). Food and water were made available ad libitum.
After confirming of pregnancy on the 14th days after mating, female rats were ran- domly divided into six groups (experimental 1) : the control group, the 50 mg/kg Ginseng radix-treated group, the 100 mg/kg Ginseng radix-treated group, the noise-treated group, the noise- and 50 mg/kg Ginseng rad- ix-treated group, and the noise- and 100 mg/kg Ginseng radix-treated group and div- ided into five groups (experimental 2) : the control group, the noise-treated group, the noise- and 10 mg/kg Ginseng radix-treated group, and the noise-and 50 mg/kg Ginseng radix-treated group, and the noise- and 100 mg/kg Ginseng radix-treated group (n = 5 for each group). Starting on the 15th day of preg- nancy, rats of the prenatal noise-treated groups were applied with the 95 decibel su- personic machine sound for 1 h once a day until delivery
14).
After birth, the offspring in each group was left undisturbed together with the re- spective mother for 28 days, and then, off- spring rats were acupunctured at Chung-Wan (CV
12) acupoint once a day for 7 consecutive
days at the respective doses; they were sacri- ficed 6 weeks after birth. To obtain the aque- ous extract of Ginseng radix, 200 g of Ginseng radix was added to distilled water, and extraction was performed by heating at 80℃ concentrated with a rotary evaporator, and lyophilized. The resulting powder, weigh- ing 20 g (a collection rate of 10%), was di- luted with saline.
2.2. Tissue preparation
For the sacrificial process, animals were first weighed and overdosed with Zoletil 50(10 mg/kg, i.p.; Vibac, Carros, France). After a complete lack of response was observed, the rats were transcardially perfused with 50 mM phosphate-buffered saline (PBS) and then with 4% paraformaldehyde in 100 mM phos- phate buffer (PB) at pH 7.4. The brains were dissected, postfixed in the same fixative over- night, and transferred into a 30% sucrose sol- ution for cryoprotection. Serial coronal sec- tions of 40 ㎛ thickness were made using a freezing microtome (Leica, Nussloch, Germa- ny).
2.3. NADPH-d histochemistry
Sections were then stained for
NADPH-d activity according to a previously
described protocol
15). In brief, free-floating
sections were incubated at 37℃ for 1 h in 100
mM PB containing 0.3% Triton X-100, 0.1
mg/ml nitroblue tetrazolium, and 0.1 mg/ml β
-NADPH. The sections were then washed
three times with PBS and mounted onto gela- tin-coated slides. The slides were air dried overnight at room temperature, and coverslips were mounted using Permount®.
2.4. Data analysis
To score the number of NADPH-d-pos- itive cells in each area of the hippocampus, cell counting was performed using Image- Pro®Plus computer-assisted image analysis system (Media Cyberbetics Inc., Silver Spring, MD, USA) attached to a light microscope (Olympus, Tokyo, Japan). The staining in- tensities of the processed sections were as-
sessed in a quantitative fashion according to a microdensitometrical method based on opticval density using and image analyzer (Multiscan, Fullerton, CA).
2.5. Statistical analysis
Statistical analysis was performed by Student's t-test. The results were presented as the mean ± standard error mean (S.E.M.).
Differences were considered significant for P
< 0.05.
3. Results
3.1. Number of NADPH-d-positive cells in CA1 region of hippocampus
The number of NADPH-d-positive cells in the CA1 region was about 128.92 ± 12.59/mm
2in the control group, 123.04 ± 7.54/mm
2in the 50 mg/kg Ginseng rad- ix-treated group, and 114.71 ± 10.16/mm
2in the 100 mg/kg Ginseng radix-treated group.
This number was significantly increased to 231.86 ± 20.69/mm
2in the noise-treated group, but was dose-dependently decreased again to 206.37 ± 11.57/mm
2in the noise- and 50 mg/kg Ginseng radix-treated group, and 189.71 ± 20.10/mm
2in the noise- and 100 mg/kg Ginseng radix-treated group (Fig. 1).
3.2. Number of NADPH-d-positive cells in CA2 and CA3 regions of hippo- campus
The number of NADPH-d-positive cells in the CA2 and CA3 regions was about 30.40
Fig. 1. Mean number of NADPH-d-positive cellsin the CA1 region in each group.
* represents P < 0.05 compared to the control group.
# represents P < 0.05 compared to the noise-treated group.
A, the control group
B, the 50 mg/kg Ginseng radix-treated group
C, the 100 mg/kg Ginseng radix-treated group
D, the noise-treated group
E, the noise- and 50 mg/kg Ginseng rad- ix-treated group
F, the noise- and 100 mg/kg Ginseng rad- ix-treated group
0 50 100 150 200 250 300
A B C D E F
Number of NADPH-d-positive cells in the CA1 region (mm2) *
#
± 4.50/mm
2in the control group, 34.00 ± 3.11/mm
2in the 50 mg/kg Ginseng rad- ix-treated group, and 23.20 ± 2.30/mm
2in the 100mg/kg Ginseng radix-treated group. This number was significantly increased to 41.20 ± 4.33/mm
2in the noise-treated group, but was dose-dependently decreased again to 38.00 ± 3.13/mm
2in the noise- and 50 mg/kg Ginseng radix-treated group, and 38.20 ± 4.67/mm
2in the noise- and 100 mg/kg Ginseng rad- ix-treated group (Fig. 2).
3.3. Number of NADPH-d-positive cells in the dentate gyrus region of hippoca- mpus
The number of NADPH-d-positive cells in the dentate gyrus region was about 56.54 ± 5.89/mm
2in the control group, 54.19 ± 3.71/mm
2in the 50 mg/kg Ginseng rad- ix-treated group, and 50.79 ± 5.90/mm
2in the 100 mg/kg Ginseng radix-treated group. This number was significantly increased to 91.88 ± 9.23/mm
2in the noise-treated group, but was dose-dependently decreased again to 100.79 ± 7.73/mm
2in the noise- and 50 mg/kg Ginseng radix-treated group, and 93.19 ± 20.10/mm
2in
Fig. 2. Mean number of NADPH-d-positive cellsin the CA2 and CA3 regions in each group.
* represents P < 0.05 compared to the control group.
# represents P < 0.05 compared to the noise-treated group.
A, the control group
B, the 50 mg/kg Ginseng radix-treated group
C, the 100 mg/kg Ginseng radix-treated group
D, the noise-treated group
E, the noise- and 50 mg/kg Ginseng rad- ix-treated group
F, the noise- and 100 mg/kg Ginseng rad- ix-treated group
0 10 20 30 40 50
A B C D E F
Number of NADPH-d-positive cells in the CA2-CA3 regions (mm2) *
#
Fig. 3. Mean number of NADPH-d-positive cells in the dentate gyrus region in each group.
* represents P < 0.05 compared to the control group.
# represents P < 0.05 compared to the noise-treated group.
A, the control group
B, the 50 mg/kg Ginseng radix-treated group
C, the 100 mg/kg Ginseng radix-treated group
D, the noise-treated group
E, the noise- and 50 mg/kg Ginseng rad- ix-treated group
F, the noise- and 100 mg/kg Ginseng rad- ix-treated group
0 20 40 60 80 100 120
A B C D E F
Number of NADPH-d-positive cells in the dentate gyrus (mm2) *
#
the noise- and 100mg/kg Ginseng rad- ix-treated group (Fig. 3).
3.4. The density of NADPH-d-positive cells in lateral hypothalamic area (LHA) region of hypothalamus
The density of NADPH-d-positivity in the hypothalamic LHA region was 129.42 ± 1.90 in the control group. This was increased significantly to 153.76 ± 1.47, in the noise stress-treated group compared to the control group; but the densities dropped to 139.37 ± 2.56 in the noise- and 10 mg/kg Ginseng rad- ix-treated group, to 124.84 ± 1.86 in the noise- and 50 mg/kg Ginseng radix-treated
group, and to 121.92 ± 1.61 in the noise- and 100 mg/kg Ginseng radix-treated group (Fig.
4).
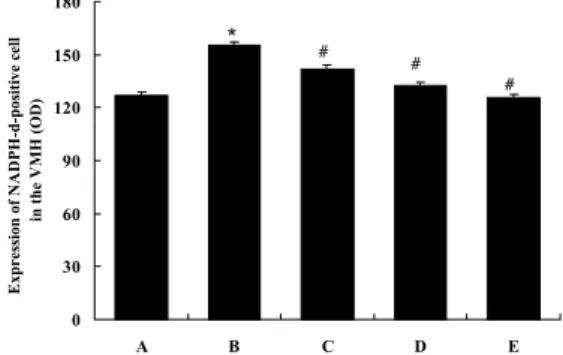
3.5. The density of NADPH-d-positive cells in ventromedial hypothalamic nucleus (VMH) region of hypotha- lamus
The density of NADPH-d-positivity in the hypothalamic VMH region was 127.11 ± 1.27 in the control group. This was increased significantly to 155.47 ± 1.31, in the noise stress-treated group compared to the control group; but the densities dropped to 141.92 ± 3.18 in the noise- and 10 mg/kg Ginseng rad-
Fig. 5. Mean densities of NADPH-d-positive cells in the ventromedial hypothalamic nucleus (VMH) region in each group.* represents P < 0.05 compared to the control group.
# represents P < 0.05 compared to the noise-treated group.
A, the control group B, the noise-treated group
C, the noise- and 10 mg/kg Ginseng rad- ix-treated group
D, the noise- and 50 mg/kg Ginseng rad- ix-treated group
E, the noise- and 100 mg/kg Ginseng rad- ix-treated group
0 30 60 90 120 150 180
A B C D E
Expression of NADPH-d-positive cell in the VMH (OD)
*
# #
#
Fig. 4. Mean densities of NADPH-d-positive cells in the lateral hypothalamic area (LHA) re- gion in each group.
* represents P < 0.05 compared to the control group.
# represents P < 0.05 compared to the noise-treated group.
A, the control group B, the noise-treated group
C, the noise- and 10 mg/kg Ginseng rad- ix-treated group
D, the noise- and 50 mg/kg Ginseng rad- ix-treated group
E, the noise- and 100 mg/kg Ginseng rad- ix-treated group
0 30 60 90 120 150 180
A B C D E
Expression of NADPH-d-positive cell in the LHA (OD)
*
#
# #
ix-treated group, to 132.42 ± 2.41 in the noise- and 50 mg/kg Ginseng radix-treated group, and to 125.71 ± 1.57 in the noise- and 100 mg/kg Ginseng radix-treated group (Fig.
5).
3.6. The density of NADPH-d-positive cells in periventricular nucleus (PVN) region of hypothalamus
The density of NADPH-d-positivity in the hypothalamic PVN region was 123.41 ± 3.05 in the control group. This was increased significantly, to 160.61 ± 0.60, in the noise stress-treated group compared to the control group; but the densities dropped to 141.65 ±
4.06 in the noise- and 10 mg/kg Ginseng rad- ix-treated group, to 123.87 ± 2.11 in the noise- and 50 mg/kg Ginseng radix-treated group, and to 118.84 ± 1.39 in the noise- and 100 mg/kg Ginseng radix-treated group (Fig.
6).
4. Discussion
Stressful experiences during the develop- ment period may exert a long-term effect on the hippocampal functions and may induce various psychosomatic problems such as me- ntal retardation and developmental disorders.
Various prenatal stresses have been reported to induce structural abnormality in the hippo- campal formation. It was reported that pre- natal stress reduced the density of the pyr- amidal neurons, decreased the total hippo- campal volume, and induced the synaptic loss in the hippocampus
4,9). In addition, Coe et al.
11)suggested that prenatal environment can alter behavior, dysregulate neuroendocrine systems, and affect the hippocampal structure of pri- mates in a persistent manner through sup- pression of neurogenesis.
NO is diffusible free radical that has rec- ognized as a biological messenger involved in several physiological and pathological func- tions
16). In the CNS, NO has been implicated in various neurophysiological functions includ- ing feeding, anxiety, immune response, and synaptic plasticity
17-20). Moreover, NO has also been implicated in the physiological processes of learning and memory formation and admin- istration of NOS inhibitors results in learning
Fig. 6. Mean densities of NADPH-d-positive cellsin the periventricular nucleus (PVN) region in each group.
* represents P < 0.05 compared to the control group.
# represents P < 0.05 compared to the noise-treated group.
A, the control group B, the noise-treated group
C, the noise- and 10 mg/kg Ginseng rad- ix-treated group
D, the noise- and 50 mg/kg Ginseng rad- ix-treated group
E, the noise- and 100 mg/kg Ginseng rad- ix-treated group
0 30 60 90 120 150 180
A B C D E
Expression of NADPH-d-positive cell in the PVN (OD)
*
#
# #