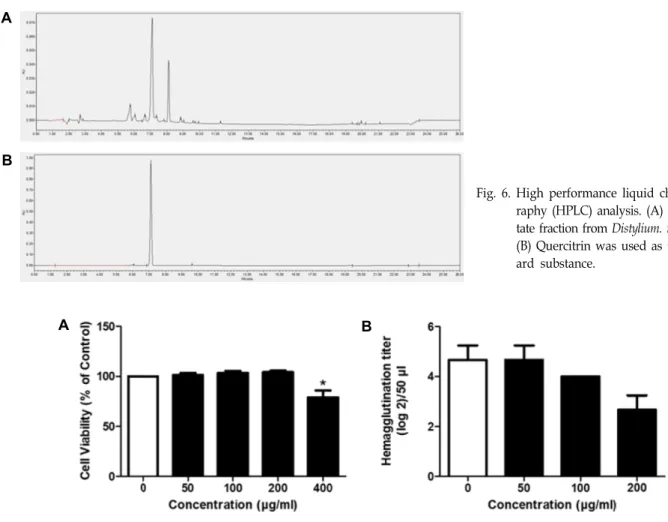
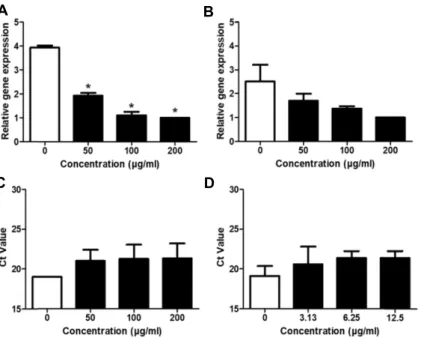
관련 문서
Inhibition effects of flavonoids on hOAT1 and hOAT3: The inhibitory effects of five flavonoids such as morin, naringin, naringenin, silybin and quercetin on the uptake of [
Inhibitory effects of classified methanol extracts of Smilax china L on the COX-2 and iNOS expression of human colorectal cancer cell lines... Inhibitory
Based on the experimental results, the execution time in the matching process for 36 fingerprint minutiae, 200 chaff minutiae and 34 authentication fingerprint
The purpose of this study was to investigate the effects of sound-finding strategies based on phonological awareness on word recognition, oral reading
In the present study we investigated the proliferation effect of human oral cancer KB cell treated with pulsatilla koreana extract.. We analyzed the effects of this
출처 : IAEA 발표 자료(Comprehensive inspection exercise at bulk handling facilities, “U-235 Enrichment measurements by gamma-ray spectroscopy”) 13. Uranium
The study on the clinical efficacy of korean red ginseng extract on postmenopausal syndrome..
The main effect according to the results of the repeated measures ANOVA on physical fitness based on test time showed a significant difference in grip