The Effects of the Expression of GATA Binding Protein 6 on Heart and Brain Development
Jungwon Seo1,2*
1College of Pharmacy, Wonkwang University, Iksan 570-749, Korea
2Institute of Pharmaceutical Research and Development, College of Pharmacy, Wonkwang University, Iksan 570-749, Korea Received October 10, 2015 /Revised November 10, 2015 /Accepted November 10, 2015
GATA binding protein 6 (GATA6) is a transcription factor that is expressed in the early blastocyst stage and controls the expression of important genes in the differentiation and development of the heart, pancreas, and intestine. This study confirmed the role of GATA6 in cell differentiation and or- gan development using mouse embryonic stem cells and zebrafish, respectively. First, the mouse em- bryonic stem cells were differentiated into pacemaker cardiomyocytes. An RT-PCR analysis revealed that the expression of the GATA6 gene was greatly increased from day 4 of differentiation. The ex- pression of GATA6 was upregulated prior to increased expression of NK2 homeobox 5 (Nkx2.5) and myocyte enhancer factor 2C (MEF2C), which are critical transcription factors involved in regulating heart formation. To examine the role of GATA6 in development, GATA6 morpholino was micro- injected into zebrafish embryos. Knockdown of GATA6 expression significantly decreased the heart size and heart rate in the zebrafish compared to a control. In addition, the brains were degenerated in the GATA6 morpholino-injected zebrafish. Acridine orange staining showed that knockdown of GATA6 expression increased apoptotic cells in the brain. Interestingly, knockdown of GATA6 ex- pression decreased apoptotic cells in the early bud stage. This study points to the importance of the GATA6 gene in heart and brain development.
Key words : Brain, embryonic stem cells, GATA6, heart, zebrafish
*Corresponding author
*Tel : +82-63-850-6819, Fax : +82-63-850-6819
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2015 Vol. 25. No. 11. 1230~1234 DOI : http://dx.doi.org/10.5352/JLS.2015.25.11.1230
서 론
GATA6는 6개의 포유류 GATA 전사 인자 family의 하나로 (G/A)GATA(A/T) DNA motif와 결합하는 2개의 잘 보존된 zinc-finger DNA binding domain을 가지고 있다[12]. GATA1, GATA2, GATA3는 신경계와 조혈세포의 발달에 관여하는 것 으로 잘 알려져 있고, GATA4, GATA5, GATA6는 심장, 장, 난소, 혈관 등의 여러 장기의 발달에 관여하는 것으로 알려져 있다[18]. GATA6는 초기 배반포(blastocyst) 단계에서 발현되 며, GATA6 -/- 쥐는 배아기(E5.5~E6.5)에서 내배엽 분화의 결 함에 의해 태내 사망하게 된다[16]. GATA6는 발생 초기 원시 내배엽 생성과 분화에서 중요한 역할을 하는 것 뿐만 아니라 [1], 심장, 간, 췌장, 폐, 장 등에서 발현되며 조직 특이적 유전자 발현을 조절하는 역할을 한다[14]. 인간의 GATA6유전자의 돌 연변이는 췌장 무형성이나 선청성 심장 기형과 관련된 질환을 유발하는 것이 밝혀져 있다[3]. 심장 발달에 있어서 GATA4와
GATA6는 ventricular myosin heavy chain (vMHC)와 같은 심장 특이적 유전자 발현을 제어한다[17]. 뇌와 관련하여서는 인간과 쥐의 성체 뇌에서 GATA6의 발현이 확인되었다[7].
GATA6 knock-out 쥐는 초기 발달 결함에 의해 발달이 중단되 기 때문에, 이후 장기 발달에서의 GATA6의 역할을 확인하는 데 어려움이 있다. 따라서, 본 연구에서는 zebrafish의 mor- pholino를 이용하여 GATA6를 knock-down 시켜 GATA6가 심장과 뇌 발달에 미치는 영향을 조사하였다.
쥐 배아줄기세포(embryonic stem cells, ES cells)는 쥐 배아 의 발달초기단계(E3.5)의 내세포괴(inner cell mass, ICM)에서 유래한다[4]. 배아줄기세포는 자기복제능과 분화능을 가진 세 포로서, 대부분의 장기 조직들로 분화하는 특성을 가지고 있 다. 따라서, 배아줄기세포를 단일한 성숙 세포로 분화시킬 수 있다면, 세포 이식을 통해 질병 치료에 기여할 수 있을 것이다.
또한 줄기세포의 조직 특이적 세포로의 분화 과정은 인체 발 생 과정에서의 분화 시의 분자 조절 기전을 반영한다. 따라서, 줄기세포 분화에 영향을 미치는 분자는 개체 발생 과정에도 영향을 미칠 가능성이 높다.
Zebrafish는 발생 과정에서 척추 동물 배아 발생의 특징인 내배엽, 중배엽, 외배엽을 형성하고 배아 축을 만드는 원장형 성(gastrulation) 단계를 거친다[9]. Zebrafish의 수정란은 자궁 외에서 투명한 상태로 발달하므로, 직접 관찰이 가능하다. 또 한, 48시간 이내에 대부분의 장기가 형성되므로 빠르게 그 결
과를 확인할 수 있다. Zebrafish는 조직 이식이나 유전자 조작 도 간편하여, morpholino를 1-4세포기의 배아에 주입하는 것 만으로 유전자의 knockdown이 가능하다. 따라서, 발달 과정 과 그에 관여하는 인자를 연구하기에 적합한 연구 동물이다.
뿐만 아니라, 형광 reporter 유전자를 활용한 질병모델로의 개 발도 활발하게 진행되고 있으며, 약물 개발을 위한 스크리닝 에도 사용되고 있다[13].
본 실험에서는 쥐 배아줄기세포와 zebrafish를 이용하여 GATA6의 심장과 뇌 발달에 미치는 영향을 연구하였다. 배아 줄기세포의 심근세포 분화 시 GATA6의 발현이 급격하게 증 가함을 확인하였고, zebrafish에서 GATA6의 발현 감소는 심 장 발달의 저하와 더불어 신경세포의 apoptosis를 유도함을 확인하였다.
재료 및 방법
배아줄기세포 배양
Feeder cell 비의존적인 E14K 배아줄기세포는 젤라틴으로 코팅된 dish에서 15% FBS, 0.1% 2-mercaptoethanol, 1,000 U/ml LIF를 포함한 DMEM 배지(Gibco)로 배양되었다[19, 21]. 세포 배양 배지에서 LIF를 제거하고 25 μl의 배지에 3X103 개의 세포를 넣은 후 hanging drop을 만들어 심근세포로의 분화를 유도하였다. 이 hanging drop을 Petri dish의 뚜껑에 놓고 배양하여, 세포들이 서로 붙어서 배상체(embryoid bod- ies, EBs)를 형성하도록 하였다. 2일 후, 이 배상체를 모아 Petri dish에서 4일 동안 부유 상태로 배양하였다. 그 후, 젤라틴 코 팅된 조직 배양 dish에 옮겨 부착 배양하였다. 부착된 세포는 자발적인 heartbeat을 나타내었다.
Reverse-transcription polymerase chain reaction (RT-PCR)
배아줄기세포, 배상체, E12.5의 쥐 태아에서 분리한 심장을 Trizol reagent (Invitrogen)에 lysis한 후, SuperScript II–re- verse transcriptase와 oligo-dT primer (Invitrogen)를 사용하 여 first-strand cDNA를 합성하였다. PCR 반응에 사용된 pri- mer들은 다음과 같다. GATA6, 5’-CGG TCA TTA CCT GTG CAA TG-3’와 5’-GAG GAA GTA GGA GTC ATA GG-
3’; MHC, 5’-CCG AAA GTC AGA GAA GGA AC-3’와 5’-AGG TAT AGA TCA TCC AGG CC-3’; myosin light chain 2v (MLC2v), 5’-GCC AAG AAG CGG ATA GAA GG
-3’와5’-TTG TGT GGT CAG CAT CTC CC-3’; elongation factor 1 α (EF1α), 5’-TCA CAC AGC CCA CAT AGC AT-3’와 5’-CAC CAC TGA TTA AGA CTG GG-3’
실험 동물
AB wild-type (WT) strain zebrafish (Danio rerio)는 ‘‘The
Zebrafish Book’’ [20]에서 기술된 방법으로 사육하였다. 성숙 zebrafish는 14:10 시간의 낮과 밤의 주기로 28.5℃의 수온을 유지, 폐쇄 순환 여과 시스템을 갖춘 수조에서 사육하였다[10].
Zebrafish의 알의 채취는 채취 전날 특수망을 설치한 알 채취 용 수조에 암컷과 수컷 성어를 1:2의 비율로 넣고 다음날 광주 기 시작 1~2시간 경과 후 알을 채취하였다. 배(embryo)의 단계 는 수정 후 시간(hours post-fertilization, hpf)으로 표시하였다.
동물 실험은 원광대학교 동물실험윤리위원회의 승인을 받아 실시하였다(WKU13-48).
Microinjection
GATA6 antisense morpholino nucleotide는 Open Biosys- tems에서 구입하였다[7]. Morpholino는 exon-intron junction 에 결합하도록 디자인되었다. 0.5 pmol의 GATA6 morpholi- no를 zebrafish배아의 1~4 세포 stage의 난황에 주입하였다.
control에는 injection buffer를 주입하였다. 12, 28, 48 hpf 단계 의 배아를 methyl cellulose에 놓고 현미경으로 관찰하였다.
Acridine orange 염색
살아있는 zebrafish 배아를 5 μg/ml acridine orange를 함 유한 배지에 30분간 빛을 차단한 상태로 둔 후, 3차례 정도 씻어준다. Methyl cellulose에 zebrafish를 올려 고정시키고 형 광 현미경으로 FITC 필터 하에서 관찰하였다. Acridine or- ange는 세포자멸사(apoptosis)한 세포의 DNA에 결합하여 형 광을 나타내는 물질로, 세포자멸사를 살아있는 상태에서 확인 할 수 있다.
결과 및 고찰
배아줄기세포의 심근세포 분화
쥐의 배아줄기세포(E14K)의 배지에서 LIF를 제거하고, 3X103 개의 세포가 들어있는 25 μl 의 배지 방울을 Petri dish의 뚜껑에 놓고 2일간 배양하여, 세포들이 서로 붙어서 배상체를 형성하게 한 후, 부유상태에서 4일간 배양하면, 이 배상체들은 자발적으로 beating 하는 pacemaker 심근세포로 분화하게 된 다. 이렇게 분화된 세포들이 심근세포로 분화되었는지 여부를 확인하기 위하여, 심근세포 특이적인 마커인 Nkx2.5, MEF2C, MHC와 MLC2v의 발현을 RT-PCR을 이용하여 확인하였다 (Fig. 1). 그 결과, 심근세포 분화에 중요한 전사인자인 Nkx2.5 와 MEF2C의 발현이 각각 4일째와 5일째에 증가함을 확인하 였다. 또한, MHC와 MLC2v의 발현이 6일째인 분화 후반기에 증가함을 확인하였다. 이때 GATA6는 배아줄기세포 때에 약 간의 발현을 보인 후, 배상체를 형성하는 hanging drop 시기 에는 발현이 감소하게 되고 4일째부터 그 발현이 급격하게 증가하는 양상을 나타내었다. GATA6의 발현은 다른 심장 발 생에 중요한 전사인자인 Nkx2.5와 MEF2C의 발현보다 더욱
Fig. 1. GATA6 expression is increased during the differentiation of mouse ES cells into pacemaker cardiomyocytes. The mouse ES cells were suspended in a hanging drop for the formation of EBs. After 2 days (on D3), the EBs were placed into a bacterial Petri dish. After 4 days (on D7), the EBs were plated on a gelatin-coated tissue culture dish and expressed a spontaneous heartbeat. ES cells, EBs, or mouse heart from E12.5 embryo were analyzed by RT-PCR to detect mRNA expression of GATA6, Nkx2.5, MEF2C, MHC, MLC2v and EF1α.
A B
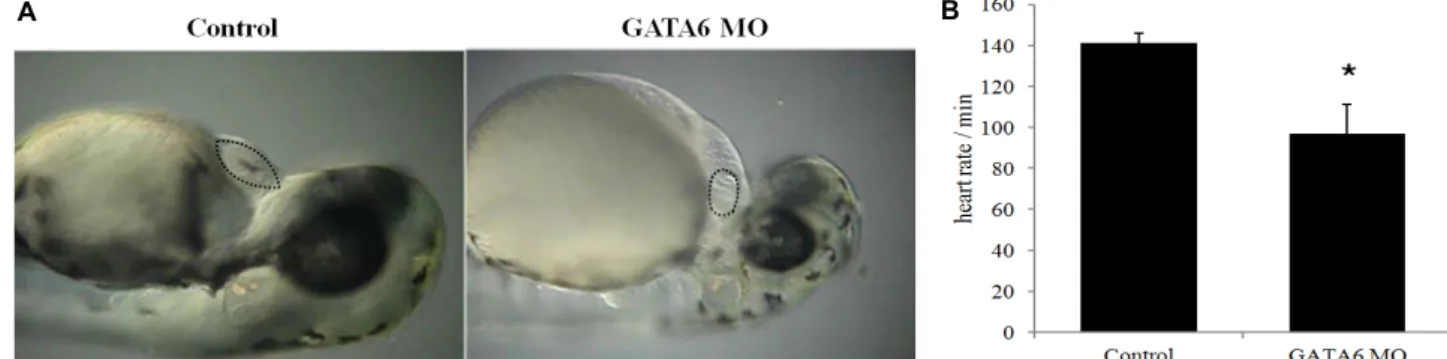
Fig. 2. Knockdown of GATA6 gene decreases heart size and heart rate in the zebrafish. GATA6 morphants (MO) were injected with GATA6 morpholino at one to four-cell stage (n=35) and control zebrafish were injected with injection buffer (n=32).
(A) The representative images of control or GATA6 MO were taken at the stage of 48 hpf. (B) Heart rate was measured by the number of ventricular contraction per minute in control or GATA6 MO zebrafish at the stage of 48 hpf. Data are represented as mean ± S.D. (n=3). Statistical significance of differences between test groups were determined using student’s t test and the difference was considered significant at a *p<0.05.
크고 빠른 변화를 나타내었다. 이 결과는 GATA6가 쥐 배아줄 기세포의 분화 과정에서 초기에 그 운명을 조절할 가능성을 제시한다. 최근의 연구 결과에 의하면, 배양 중인 배아에 LIF 를 처리하면 JAK/STAT 신호전달계를 통해 GATA6 양성의 primitive endoderm의 비율을 증가시킨다고 한다[15]. 따라 서, 배아줄기세포 상태에서의 GATA6의 발현은 LIF에 의한 영향일 것으로 사료된다. 이후 LIF의 제거와 배상체의 형성은 GATA6의 발현을 감소시키고 4일째에는 GATA6발현의 급격 한 증가가 나타난다. 기존의 연구 결과에서도 배아줄기세포의 pacemaker 심근세포 분화 시에 GATA4와 GATA6의 발현이 증가함이 보고되어 있으며[6], GATA4/GATA6 double heter-
ozygous mutant embryo는 MHC와 MEF2C의 발현을 감소시 킨다는 보고가 있다[22]. 이번 연구 결과에서도 GATA6의 발 현 증가가 먼저 나타나고 이후 5일과 6일째에 MEF2C와 MHC 의 발현이 급격하게 증가함을 확인하였다.
GATA6 유전자의 knock-down 시의 심장 발달 변화 배아줄기세포가 pacemaker 심근세포로 분화하는 과정에 서 GATA6 발현 증가의 역할을 in-vivo에서 확인하기 위하여, GATA6 morpholino를 제작하여 zebrafish 배아의 1~4 세포 stage의 난황에 injection하였다(Fig. 2). Injection buffer를 주 입한 control의 경우 35마리 중 97%(34 마리)에서 정상 크기의 심장 발달을 보인 반면, GATA6 morpholino 를 주입한 배아 (n= 32)에서는 90%(29마리)에서 심장의 크기가 배아 발달기 48 hpf의 정상 심장 크기의 50% 이하로 감소하였다(Fig. 2A).
특히, 분당 심박동수는 control의 경우 평균 141회였으나, GATA6 morpholino를 주입한 배아의 경우 평균 97회로 약 31% 감소하였다(Fig. 2B). 이러한 심박동수의 감소는 GATA6 발현 감소에 의한 pacemaker 심근세포의 분화의 결함과 관련 될 것으로 추측된다. 최근 연구에 의하면, 심근세포 특이적 GATA6의 발현 결손은 HCN4와 Id2의 발현 감소와 방실전도 의 이상 및 방실결절 형성의 결함을 나타내어 심장전도계의 발달에 중요한 역할을 한다고 보고되었다[11].
GATA6 knock-down 시의 뇌 발달 변화
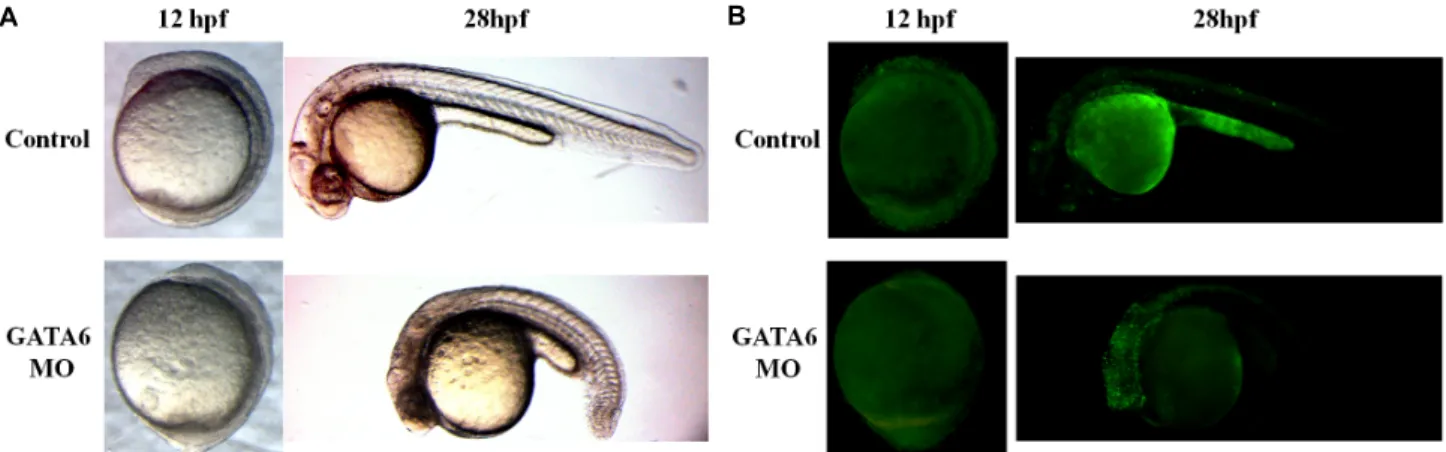
놀랍게도 GATA6 morpholino를 주입하여 GATA6의 발현 을 감소시킨 배아는 28 hpf 시기에 뇌 발달의 결함을 나타내었 다(Fig. 3A). Control zebrafish의 경우 38마리 모두에서 정상 적인 뇌의 발달을 보인 반면, GATA6 morpholino를 주입한 zebrafish (n=39)는 82%(32마리)에서 뇌 구조의 형성에 실패하 였고, 뇌를 구성하는 세포들이 어두운 색을 띄었다. 또한 15%(6마리)에서는 16~18 hpf에서의 성장 arrest가 발생하였
A B
Fig. 3. Knockdown of GATA6 gene increases apoptosis in the brain of zebrafish. GATA6 morphants (MO) were injected with GATA6 morpholino at one to four-cell stage (n=39) and control zebrafish were injected with injection buffer (n=38). (A) The representa- tive images of control or GATA6 MO were taken at the stage of 12 or 28 hpf. (B) The live zebrafish were stained with acridine orange for detecting apoptotic cells and the representative fluorescent images of control or GATA6 MO were taken at the stage of 12 or 28 hpf.
다. 이러한 뇌세포의 변화가 세포자멸사에 의한 것인지를 확 인하기 위하여, acridine orange를 이용하여 살아있는 상태의 zebrafish 배아에서의 세포자멸사를 비교하였다(Fig. 3B).
Acridine orange는 세포자멸사한 세포의 DNA에 결합하여 형 광을 나타내는 물질이다. 28 hpf에서 GATA6 morpholino를 주입한 zebrafish의 뇌에서는 control zebrafish의 뇌에 비하여 세포자멸사가 현저하게 증가함을 확인하였다. 그러나, GATA6 morpholino를 주입한 zebrafish는 12 hpf의 초기 발생 단계 (bud 단계)에서의 세포자멸사가 정상적으로 발생하지 않았다.
기존의 연구에 의하면, V12Ras 혹은 p53-/-의 성상세포종에 서 GATA6의 발현 감소는 암발생을 증가시키는 것으로부터 GATA6가 암억제인자로 작용하는 것으로 보고되었다[8]. 그 러나, 정상 성상세포에서는 세포 사멸을 증가시키는 역할을 하지 않았다. 한편, 대장암 세포에서 GATA6의 발현은 암세포 의 생존을 증진하는 효과를 나타내었고[2], 복강 대식세포에서 도 GATA6가 발현되지 않으면 생존율이 감소하였다[5]. 따라 서, GATA6가 세포의 생존과 사멸에 미치는 영향은 분화된 세포 특이적 성격을 나타내는 것으로 보인다. 본 연구에서도 zebrafish의 발생 초기 bud 단계(12 hpf)의 분화되지 않은 세포 들에서는 정상적인 세포자멸사의 감소가 나타난 반면, 뇌 형 성과 더불어 뇌세포 특이적으로 세포자멸사의 급격한 증가가 확인되었다. 이러한 뇌세포의 세포자멸사가 초기 단계의 정상 세포자멸사의 실패에 의한 결과인지, 아니면 GATA6 발현 감 소에 따른 뇌세포의 생존 실패에 의한 현상인지에 대해서는 더 연구가 필요할 것으로 생각된다. 이전 연구에 따르면, 신경 세포와 성상세포 모두에서 GATA6의 발현이 확인되었다[8].
따라서, 세포자멸사한 뇌세포가 성상세포인지 신경세포인지 에 대해서도 더 연구가 필요하다.
본 연구에서는 쥐 배아줄기세포와 zebrafish를 이용하여 GATA6의 세포 분화와 개체 발달에서의 역할을 확인하였다.
배아줄기세포의 pacemaker 심근세포 분화 초기에 GATA6의 발현이 급격하게 증가하고 zebrafish에서 GATA6의 발현 감 소가 심장 크기와 심박동수의 감소를 유발하는 것으로부터 심장의 발달과 pacemaker 심근세포의 분화에 GATA6의 발현 이 중요한 역할을 함을 알 수 있었다. 뿐만 아니라, GATA6의 발현 감소는 zebrafish 뇌에서 세포자멸사를 증가시킴을 확인 하였다. 본 연구 결과는 심장과 뇌 발달에서 GATA6 발현의 중요성을 시사한다. 특히, 본 연구를 통하여 pacemaker 심근 세포 분화에서의 GATA6의 중요성과 뇌 세포의 생존에서의 GATA6의 역할을 새롭게 제시하였다.
감사의 글
이 논문은 2013학년도 원광대학교의 교비지원에 의해 수행됨.
References
1. Aronson, B. E., Stapleton, K. A. and Krasinski, S. D. 2014.
Role of GATA factors in development, differentiation, and homeostasis of the small intestinal epithelium. Am. J. Physiol.
Gastrointest. Liver Physiol. 306, G474-490.
2. Cai, W. S., Shen, F., Li, J. L., Feng, Z., Wang, Y. C., Xiao, H. Q. and Xu, B. 2014. Activated protease receptor-2 induces GATA6 expression to promote survival in irradiated colon cancer cells. Arch. Biochem. Biophys. 555-556, 28-32.
3. Chao, C. S., McKnight, K. D., Cox, K. L., Chang, A. L., Kim, S. K. and Feldman, B. J. 2015. Novel GATA6 mutations in patients with pancreatic agenesis and congenital heart malformations. PLoS One 10, e0118449.
4. Evans, M. J. and Kaufman, M. H. 1981. Establishment in culture of pluripotential cells from mouse embryos. Nature 292, 154-156.
5. Gautier, E. L., Ivanov, S., Williams, J. W., Huang, S. C.,
초록:심장과 뇌 발달에서 GATA6 유전자 발현 감소가 미치는 영향
서정원1,2*
(1원광대학교 약학대학, 2원광대학교 약품연구소)
GATA binding protein 6 (GATA6)는 초기 배반포 단계에서 발현이 시작되어, 심장, 췌장, 장 등의 분화와 발달 에 중요한 유전자 발현을 조절하는 전사인자이다. 본 연구에서는 GATA6의 세포 분화와 개체 발달 과정에서의 역할을 마우스 배아줄기세포와 zebrafish를 이용하여 확인하였다. 먼저, 마우스 배아줄기세포를 박동하는 pace- maker 심근세포로 분화 유도하였다. RT-PCR을 실시하여 심근세포 분화 과정에서 GATA6 유전자 발현 변화를 확인한 결과, Gata6의 발현이 분화 4일째부터 증가함을 확인하였다. GATA6 유전자의 발현 증가는 심장 발달에 필수적인 전사인자인 NK2 homeobox 5 (Nkx2.5)나 myocyte enhancer factor 2C (MEF2C)의 발현 증가에 앞서 나타났다. GATA6 유전자가 발달 과정에 미치는 영향을 확인하기 위하여 GATA6의 morpholino를 zebrafish 배아 에 microinjection하여 발생 변화를 관찰하였다. GATA6의 발현을 knockdown시킨 zebrafish의 심장은 크기가 감 소하였고, 심박동률 또한 감소하였다. 한편, 뇌에서는 전체적인 뇌 퇴행이 관찰되었는데, acridine orange로 염색 한 결과, 뇌 전체에서의 세포사멸의 증가를 나타내었다. 흥미롭게도, GATA6의 발현 감소는 초기 bud 단계에서는 오히려 세포사멸을 감소시켰다. 본 연구는 심장과 뇌 발달에서의 GATA6 유전자의 중요성을 시사한다.
Marcelin, G., Fairfax, K., Wang, P. L., Francis, J. S., Leone, P., Wilson, D. B., Artyomov, M. N., Pearce, E. J. and Randolph, G. J. 2014. Gata6 regulates aspartoacylase ex- pression in resident peritoneal macrophages and controls their survival. J. Exp. Med. 211, 1525-1531.
6. Hashem, S. I. and Claycomb, W. C. 2013. Genetic isolation of stem cell-derived pacemaker-nodal cardiac myocytes.
Mol. Cell. Biochem. 383, 161-171.
7. Kamnasaran, D. and Guha, A. 2005. Expression of GATA6 in the human and mouse central nervous system. Brain Res.
Dev. Brain Res. 160, 90-95.
8. Kamnasaran, D., Qian, B., Hawkins, C., Stanford, W. L. and Guha, A. 2007. GATA6 is an astrocytoma tumor suppressor gene identified by gene trapping of mouse glioma model.
Proc. Natl. Acad. Sci. USA 104, 8053-8058.
9. Keller, R. 2002. Shaping the vertebrate body plan by polar- ized embryonic cell movements. Science 298, 1950-1954.
10. Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B.
and Schilling, T. F. 1995. Stages of embryonic development of the zebrafish. Dev. Dyn. 203, 253-310.
11. Liu, F., Lu, M. M., Patel, N. N., Schillinger, K. J., Wang, T. and Patel, V. V. 2015. GATA-binding factor 6 contributes to atrioventricular node development and function. Circ.
Cardiovasc. Genet. 8, 284-293.
12. Lowry, J. A. and Atchley, W. R. 2000. Molecular evolution of the GATA family of transcription factors: conservation within the DNA-binding domain. J. Mol. Evol. 50, 103-115.
13. MacRae, C. A. and Peterson, R. T. 2015. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 14, 721-731.
14. Molkentin, J. D. 2000. The zinc finger-containing tran- scription factors GATA-4, -5, and -6. Ubiquitously expressed regulators of tissue-specific gene expression. J. Biol. Chem.
275, 38949-38952.
15. Morgani, S. M. and Brickman, J. M. 2015. LIF supports prim itive endoderm expansion during pre-implantation devel-
opment. Development doi: 10.1242/dev.125021.
16. Morrisey, E. E., Tang, Z., Sigrist, K., Lu, M. M., Jiang, F., Ip, H. S. and Parmacek, M. S. 1998. GATA6 regulates HNF4 and is required for differentiation of visceral endoderm in the mouse embryo. Genes Dev. 12, 3579-3590.
17. Park, J. S., Kim, H. S., Kim, J. D., Seo, J., Chung, K. S., Lee, H. S., Huh, T. L., Jo, I. and Kim, Y. O. 2009. Isolation of a ventricle-specific promoter for the zebrafish ventricular myosin heavy chain (vmhc) gene and its regulation by GATA factors during embryonic heart development. Dev.
Dyn. 238, 1574-1581.
18. Patient, R. K. and McGhee, J. D. 2002. The GATA family (vertebrates and invertebrates). Curr. Opin. Genet. Dev. 12, 416-422.
19. Shimizu, N., Watanabe, H., Kubota, J., Wu, J., Saito, R., Yokoi, T., Era, T., Iwatsubo, T., Watanabe, T., Nishina, S., Azuma, N., Katada, T. and Nishina, H. 2009. Pax6-5a pro- motes neuronal differentiation of murine embryonic stem cells. Biol. Pharm. Bull. 32, 999-1003.
20. Westerfield, M. 1994. The Zebrafish Book : A Guide for the Laboratory Use of Zebrafish. Institute of Neuroscience.
Eugene, OR: University of Oregon.
21. Wu, J., Kubota, J., Hirayama, J., Nagai, Y., Nishina, S., Yokoi, T., Asaoka, Y., Seo, J., Shimizu, N., Kajiho, H., Watanabe, T., Azuma, N., Katada, T. and Nishina, H. 2010. p38 Mitogen-activated protein kinase controls a switch between cardiomyocyte and neuronal commitment of murine embry- onic stem cells by activating myocyte enhancer factor 2C-de- pendent bone morphogenetic protein 2 transcription. Stem Cells Dev. 19, 1723-1734.
22. Xin, M., Davis, C. A., Molkentin, J. D., Lien, C. L., Duncan, S. A., Richardson, J. A. and Olson, E. N. 2006. A threshold of GATA4 and GATA6 expression is required for car- diovascular development. Proc. Natl. Acad. Sci. USA 103, 11189-11194.