Shewanella oneidensis PKA 1008의 알긴산 분해 조효소 생산 최적 조건과 조효소의 특성
선우찬1, 김꽃봉우리2, 김동현1, 정슬아1, 김현지1, 정다현1, 정희예1, 강보경1, 박시우1, 임성미3, 홍용기4, 안동현1*
1부경대학교식품공학과/식품연구소
2부경대학교수산과학연구소
3동명대학교식품공학과
4부경대학교생물공학과
Received : December 27, 2012 / Revised : May 7, 2013 / Accepted : May 9, 2013
현재해조류는새로운기능성소재로많은연구가이루어 지고있는데, 이는육상과는다른환경에서식하는해조류 의특성상육상생물과는다른생리활성물질을함유할것 으로기대되고있기때문이며해조류의풍부한다당류가새
로운생리활성물질의보고라고할수있기때문이다[10]. 이
러한해조류의특이적생육환경으로기인한기능성으로는 항종양[15], 항혈액응고[23] 및면역력증강[2] 등이알려져 있다. 해조류의기능성성분은녹조류(green algae), 홍조류 (red algae) 및갈조류(brown algae)의골격, 구성및저장다 당류에서 많이 찾을 수 있으며, 대표적인 것은 다시마 (Laminaria japonica), 감태(Ecklonia cava), 미역(Undaria pinnatifada) 및모자반(Sargassum fulvellum)과같은갈조
류에포함되어있는알긴산이다. 알긴산은갈조류세포벽구 성다당류로서 2종류의 uronic acid인 D-mannuronic acid와 L-guluronic acid가다양한비율로 1,4 glycoside 결합을하 고있는 polyuronide이다[26]. 알긴산은분자량, 결합순서, 결 합구조에따라다양한겔형성능, 점도증진능, 수분흡수능, 결착능및필름형성능등의물리적특징을가지며, 이러한 특징은식품산업에서안정제, 보습제및증점제로이용되고
있다[8]. 또한알긴산은난소화성특성을부각시켜식이섬유
로서변비치유[3], 비만억제등의성인병예방식품으로기
대되고있으며[4], 중금속배출및콜레스테롤배출등의효
능이보고되고있다[9].
이와같은다양한기능성이밝혀지면서알긴산의이용을 확대하기위해많은연구가이루어지고있지만, 상온에서용 해시간이길고농도가높아짐에따라고점도특성을보여 산업적이용이제한적이다. 이를해결하기위해알긴산의저 분자화방법이제시되고있으며, 산업적이용이가능한알긴 Optimization of Conditions for the Production and Properties of Alginate-degrading Crude Enzyme from Shewanella oneidensis PKA 1008. Sunwoo, Chan1, Koth-Bong-Woo-Ri Kim2, Dong-Hyun Kim1, Seul-A Jung1, Hyun-Jee Kim1, Da- Hyun Jeong1, Hee-Ye Jung1, Bo-Kyeong Kang1, Si-Woo Bark1, Sung-Mee Lim3, Yong-Ki Hong4, and Dong-Hyun Ahn1*.
1Department of Food Science & Technology/Institute, Pukyong National University, Busan 608-737, Korea, 2Institute of Fisher- ies Sciences, Pukyong National University, Busan 619-911, Korea, 3Department of Food Science & Technology Tongmyong University, Busan 608-735, Korea, 4Department of Biotechnology, Pukyong National University, Busan 608-737, Korea An alginate-degrading bacterium, identified as Shewanella oneidensis PKA 1008 by 16S ribosomal RNA sequence analysis, was isolated from the green alga Ulva pertusa. Optimal conditions for the alginate-degrading ability of its crude enzyme were then determined. The optimal culture conditions for the growth of S. oneidensis PKA 1008 were pH 9, 2% NaCl, 30oC, and 24 hours incubation time. The crude enzyme produced by S. oneidensis PKA 1008 showed the highest alginate-degrading activity at pH 9, 30oC and produced 1.001 g of reducing sugar per liter in 3.5% (w/v) sodium alginate for 1 hour.
Keywords: Alginate-degrading crude enzyme, Shewanella oneidensis PKA 1008
*Corresponding author
Tel: +82-51-629-5831, Fax: +82-51-629-5824 E-mail: [email protected]
© 2013, The Korean Society for Microbiology and Biotechnology
산올리고당생산을위해알긴산분해효소에관한연구가 많이이루어지고있다.
현재해양환경, 해양동물및미생물유래의알긴산분해 효소의탐색과생화학및분자생물학적연구가활발히이루 어지고있다[18]. 알긴산분해효소는성게, 전복, 소라등해
조류 섭식 동물[30] 및 해수와 해저 토양 유래 미생물인
Azotobacter vinelandii [7], Vibrio sp. [27], Pseudomonas sp. [17], Klebsiella aerogenes [13], Flabobacterium sp.
[1], Alginovibrio aquatilis [24] 등에서생산하는것으로알 려져있지만알긴산분해효소를고효율로생산하는미생물 에관한보고는아직미비한실정이다.
따라서본연구에서는알긴산올리고당을효소적으로제조 하기위한기초연구로서, 분해중인해조류및그주변해수로 부터알긴산분해활성이우수한미생물을탐색하고, 미생물 이생성한알긴산분해조효소의분해특성을확인하고자한다.
재료 및 방법
재료
본실험에서 alginic acid sodium salt from brown algae (Aldrich Chemical Co., USA)를 사용하였으며, marine broth (MB, Difco, USA) 및 marine agar (MA, Difco, USA) 를사용하였다.
알긴산 분해 미생물의 분리 및 동정
부산의송정연안에서분해중인해조류 Ulva pertusa를
채집한 후멸균된 2% NaCl phosphate buffer (pH 7.4)을 이용하여희석하였다. 희석액은 MA 배지에분주·도말한 후 30oC에서 24시간배양하였으며, 생성된 colony 중에서형 태적으로다른 colony를선택하였다. 선택한 colony는알긴 산분해능을확인하기위해 4% 알긴산과 106 cfu/ml 농도의 균배양액을 1:1 혼합하여 30oC에서 24시간반응후점도및 환원당 함량을 측정하였다. 이 중 알긴산 분해능이 높은 colony는 (주)마크로젠에 의뢰하여 16S rRNA sequence
analysis로 분석하였고 이후 16S rDNA 염기서열은
National Center for Biotechnology Information (NCBI;
http://www.ncbi.nlm.nih.gov/)의 blast server를 사용하여 염기서열을비교분석하여균들을동정하였다.
분리·동정된 알긴산 분해 미생물의 생육 특성 확인 분리·동정된알긴산분해미생물의최적생육조건(pH, NaCl, 배양온도및배양시간)을확인하기위해 MB 배지 를기본배지로사용하였고, 균은 106 cfu/ml 농도로접종하
여각실험조건에서배양한후 600 nm에서흡광도를측정
하였다. 먼저, pH 확인을 위해 MB 배지를 0.1 N 및 1 N
HCl과 0.1 N 및 1 N NaOH로 pH 2-10으로조정하였으며, 균을접종후 30oC에서 24시간배양하였다. 최적 NaCl 농도 를확인하기 위해 최적 pH인 pH 9로 조정한 MB 배지의 NaCl 농도를 2-8%로조정한 후균을접종하고 30oC에서 24시간배양하였다. 최적 pH 및 NaCl 농도인 pH 9 및 2%
NaCl로조정한 MB 배지에균을 접종후배양온도를 10- 50oC로조정하고 24시간배양하여최적온도조건을확인하 였고, pH 9, 2% NaCl 및 30oC 조건에서균을 36시간까지 배양하며 3시간간격으로균체량을측정하여최적배양시 간조건을확인하였다.
알긴산 분해 조효소액 제조
분리·동정한알긴산분해균을최적생육조건(pH 9, 2%
NaCl, 30oC 및배양 24시간)으로대량배양하였으며, 배양 액을 원심분리기(SUPRA 22K, Hanil Science Co., Korea) 로 12,000×g, 30 min, 4oC 조건으로원심분리하여상층액 과침전물을분리하였고, 상층액을조효소로하였다. 알긴산 분해 조효소의 특성 확인
분리·동정한알긴산분해균이생성한조효소의최적알긴 산분해 pH, 온도, 알긴산농도및반응시간조건을확인하 기위해점도및환원당을측정하였다. 점도측정은점도계 (LVLTDV-II, Brookfield Co., USA)를이용하여 Richard 등 [21]의방법을참고하여 25oC, 52 cP 및 0.5 rpm 조건에서 실험을수행하였다. 환원당측정은 Somogyi-Nelson법[19]으 로 520 nm에서표준당(glucose)으로작성한검량곡선으로 환원당함량을정량하였다. 효소 1 unit는 1분에 1 µmole의 환원당을생산하는효소양으로정의하였고, 각실험조건에
서최대활성을보인것을 100%로간주하고상대활성으로
나타내었다. 최적 pH 조건을확인하기위해 0.1 N 및 1 N HCl과 0.1 N 및 1 N NaOH를이용하여 pH 2-10까지조정 하였으며, 알긴산과조효소를 1:1 혼합후 30oC에서 24시간 반응시켰다. pH 9 조건에서온도를 10-50oC로달리하여알 긴산과조효소 1:1 혼합물을 24시간반응시킨후최적온도 조건을확인하였으며, 최적알긴산농도를확인하기위해 1- 9% 농도의알긴산을조효소와 1:1 혼합한후 pH 9 및 30oC 조건에서 24시간반응시켰다. 또한반응시간에따른조효 소의알긴산분해능을확인하기위해 pH 9, 30oC 조건에서 7% 알긴산과조효소를 1:1 혼합한후 60시간까지반응시키 며시간별로분해정도를확인하였다.
결과 및 고찰
알긴산 분해균의 분리 및 동정
부산송정연안에서채취한분해중인해조류 Ulva pertusa
로부터형태적으로다른다수의 colony를분리하였으며, 그
중알긴산분해능이우수한 colony는환원당생성량이 418
µg/ml이고, 99% 점도감소율을보였다(Fig. 1). 또한이균주 의 16S rRNA 염기서열을 분석하였고(Fig. 2) NCBI blast search를통해확인한결과, Shewanella oneidensis strain (NCBI accession #; AB447987)와 99% 유사성을 보여 Shewanella oneidensis strain으로동정되었으며, Shewanella oneidensis PKA 1008로명명하였다.
알긴산 분해균의 생육 특성 확인
S. oneidensis PKA 1008의최적 pH 조건을확인한결과 (Fig. 3), pH 6, 7, 8, 9 및 10에서흡광도값은 0.279, 0.309, Fig. 1. The alginate degrading ability of isolated marine bac-
teria measured by viscomerty and reducing sugar assay.
Fig. 2. 16S rRNA nucleotide particle sequence of Shewanella oneidensis PKA 1008.
Fig. 3. Effect of pH on growth of Shewanella oneidensis PKA 1008.
Fig. 4. Effect of NaCl concentrtaion on growth of Shewanella oneidensis PKA 1008.
0.355, 0.442 및 0.298으로측정되었으며 pH 9에서흡광도 값이가장높게측정되어 S. oneidensis PKA 1008은 pH 9 에서생육활성이가장좋은것으로확인되었다.
해양에서분리한 S. oneidensis PKA 1008의 NaCl 농도와 생육관계를알아본결과(Fig. 4), 2%와 3% 농도에서흡광도 값이각각 0.449, 0.385로나타났고, 4% 이상에서는흡광도 값이급격히줄어드는것으로측정되었다. 해양유래미생물
의경우 NaCl 농도가 3% 내외의범위에서최적생육조건
인것으로알려져있으며[14], 해양에서분리한알긴산분해
균인 Bacillus licheniformis [29]와 Methylobacterium sp.
HJM27 [11]은 2%와 2.5%가최적 NaCl 농도인것으로알려 져있다. 또한 Joo 등[5]의연구에서해양에서분리한 4종의 균주는 2.5-3.0% NaCl에서알긴산분해활성이높고토양에
서분리한 1종의균주는 0.85% NaCl 이하에서분해활성이 높다고보고된바있다. 이러한결과를미루어볼때, 해양유
래의알긴산분해균의생육및알긴산분해활성이 NaCl 농
도 2% 이상에서높은것으로사료되어지며, 본연구에서분 리한 알긴산 분해균 S. oneidensis PKA 1008 역시 2%
NaCl에서 가장 높은 생육 활성을 보였다. S. oneidensis PKA 1008의최적생육온도를확인한결과(Fig. 5), 10, 20, 25, 30 및 35oC에서흡광도값이각각 0.153, 0.203, 0.313, 0.497 및 0.294로 30oC에서균주의활성이최대인것을확 인하였다. 이러한결과는해수유래미생물이 20-30oC 부근 에서생육활성이높은것과유사한것으로확인되었다.
S. oneidensis PKA 1008의시간별생육정도를확인한결 과(Fig. 6), 배양 24시간까지는균수가증가하는대수증식기 로확인되었으며, 배양 24시간부터 36시간까지는균수가완 만하게증가하는정지기로나타나본실험에서는 24시간을 S. oneidensis PKA 1008의최적생육시간조건으로하였다. 이상으로 S. oneidensis PKA 1008의최적 생육 조건은 pH 9, 2% NaCl, 30oC 및배양 24시간인것으로확인되었으 며, 이러한해양유래의알긴산분해균인 Vibrio crassostreae PKA 1002 [25]가 pH 9, 2% NaCl, 30oC 그리고 Bacillus licheniformis AL-577 [29]가 2% NaCl, pH 7.5 및 30oC인 결과와유사한것을확인할수있었으며, S. oneidensis PKA 1008도알칼리(pH 9) 및중온(30oC) 영역에서대량배양하 여균체외효소를획득할수있을것으로사료되어진다.
알긴산 분해 조효소의 특성 확인
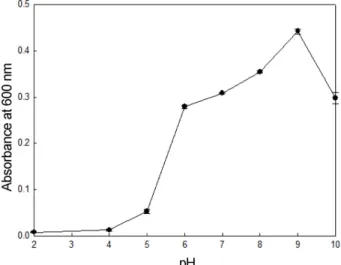
S. oneidensis PKA 1008이생성한알긴산분해조효소의 최적 pH를확인한결과(Fig. 7), pH 8, 9 및 10의알칼리영
역에서알긴산분해능활성이높았으며특히, pH 9에서분
해 활성이 가장 높은 것으로 확인되었다. 이러한 결과는 Pseudomonas sp. SM0524 [16]과 Agarivorans sp. [12]이 각각약알칼리영역인 pH 8.5 및강알칼리영역인 pH 10 부 근에서알긴산분해효소의활성이최대인결과와유사한것 으로나타났다.
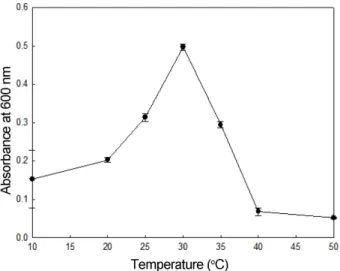
조효소의알긴산분해활성과온도와의상관관계를확인 한결과(Fig. 8) 10oC에서는분해활성이상당히낮았으나, 분해활성이 20oC부터증가하여 30oC에서최대로증가하였 으며, 그이후에는활성이감소하는것으로나타났다. 점도 측정결과에서도, 30oC에서 110 cP로가장낮은값을나타 내어조효소의알긴산분해활성이최대로보인온도와일 치하였다. 이러한결과는 Azotobacter sp. [7], Enterobacter cloacae M-1 [20] 및 Alteromonas sp. strain H-4 [22]가생 성한효소의최적활성온도가 30oC인결과와일치하였다. 또한본연구에서분리한균주의최적생육온도가 30oC인 것으로미루어보아미생물의서식환경이분비하는효소의 Fig. 5. Effect of temperature on growth of Shewanella
oneidensis PKA 1008.
Fig. 6. Effect of incubation time on growth of Shewanella oneidensis PKA 1008.
최적활성온도와상관관계가있는것으로사료되어진다. 알긴산농도를 0.5-4.5% (working concentration)로달리 하여조효소액과 1:1 혼합후 30oC에서 24시간반응시킨후 효소활성을측정하였다. 그결과(Fig. 9), 3.5%인알긴산농 도까지조효소의알긴산분해활성은증가하였고, 그이상의 농도에서는활성이다소감소하여최적기질농도가 3.5%임 을확인하였다. 이러한결과는일정알긴산농도이상에서 균체성장의급격한저하와함께알긴산분해활성이저하
된다고보고한 Yonemoto 등[31] 및 Uo 등[29]의연구와유
사한것으로배지의 alginate 농도가높아지면점성이커져
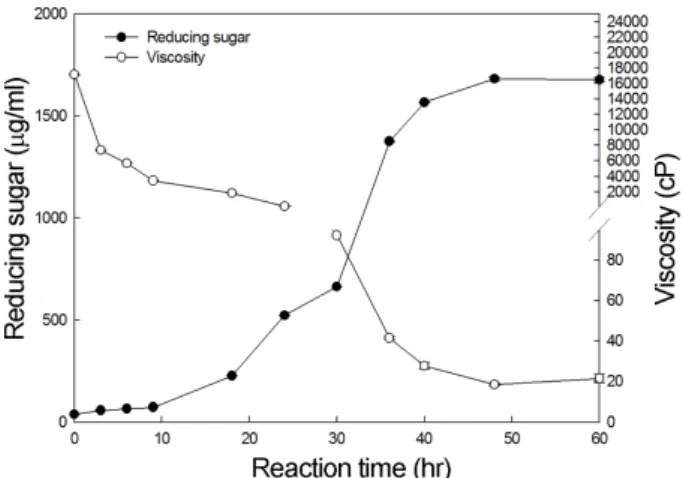
교반이제대로되지않아배지의조효소작용이잘이루어 지지않기때문인것으로사료되어진다. 또한 pH 9, 30oC 및 3.5% 알긴산조건에서 60시간까지반응한결과(Fig. 10), 환 원당생성량이반응 0시간부터반응 40시간까지점차적으로 증가하였으며, 반응 48시간에는 1681 µg/ml로가장많았으 며, 점도의경우반응 0시간에 17146 cP에서반응 48시간에 는 18.43 cP로가장낮은값을나타내었다.
따라서 S. oneidensis PKA 1008이생성한알긴산분해조 Fig. 7. Effect of pH on alginate degrading activity of crude
enzyme measured by viscometry and reducing sugar assay.
The mixture of alginate and crude enzyme were incubated for 24 h and viscosity was measured at 25oC, 52 cP and 0.5 rpm. The rel- ative activity was shown as percentages of the enzyme activity.
Fig. 8. Effect of temperature on alginate degrading activity of crude enzyme measured by viscometry and reducing sugar assay.
The mixture of alginate and crude enzyme were incubated for 24 h and viscosity was measured at 25oC, 52 cP and 0.5 rpm. The rel- ative activity was shown as percentages of the enzyme activity.
Fig. 9. Effect of alginate concentration on alginate degrading activity of crude enzyme measured by reducing sugar assay.
The mixture of alginate and crude enzyme were incubated at 30oC for 24 h. The relative activity was shown as percentages of the enzyme activity. Means with different letters, displayed in figure, are significantly different (p < 0.05).
Fig. 10. Effect of reaction time on alginate degrading activity of crude enzyme measured by reducing sugar assay.
The mixture of alginate and crude enzyme were incubated at 30oC for 72 h. The viscosity was measured at 25oC, 40 cP and 0.5 rpm.
효소는 pH 9, 배양온도 30oC, 3.5% 알긴산, 배양시간 48시 간조건에서알긴산을가장효과적으로분해하는것으로나 타났다.
이상의 결과로 미루어 보았을 때, 해양에서 분리한 S.
oneidensis PKA 1008 유래 알긴산분해 조효소의특성은 Thiang 등[28]이대부분의해양세균유래의알긴산분해효 소의최적 pH 및온도조건이 pH 7.5-8.5 및 25-50oC라고보
고한바있는데[28], 이러한결과와유사한것으로나타났
다. 또한 Methylobacterium sp. HJM27 [11], Vibrio sp.
AL-145 [6] 및 Bacillus licheniformis AL-577 [29]가각각 환원당생성능이반응 1시간에 1.217 g/l, 반응 50분에 0.355
및 0.899 g/l로산업적이용가능성이있다고보고하였으며
본연구에서분리·동정한알긴산분해균 S. oneidensis PKA 1008의환원당생성능은반응 1시간에 1.001 g/l로확인되었 다. 따라서추후 S. oneidensis PKA 1008가생산하는알긴 산분해효소를정제하여알긴산저분자화에이용가능할것 으로사료되어진다.
요 약
부산송정연안에서분해중인해조류로부터알긴산분해 미생물을분리·동정하고미생물의생육조건및미생물이 생성한 조효소의 알긴산 분해 특성을 확인하였다. Ulva
pertusa로부터 분리한 알긴산 분해균을 동정한 결과,
Shewanella oneidensis strain로확인되었으며, S. oneidensis PKA 1008 명명하였다. S. oneidensis PKA 1008의최적생 육조건을확인한결과, pH 9, 2% NaCl, 30oC 및배양 24시 간인것으로확인되었다. 또한 S. oneidensis PKA 1008 유 래알긴산분해조효소는 pH 9, 30oC에서분해활성이최대 이며, 3.5% 알긴산(working concentration)에서 1시간반응 시 1.001 g/l의환원당을생성하는것으로확인되었다.
Acknowledgments
This work was supported by a Research Grant of Pukyong National University (2013Year:20130572).
References
1. An QD, Zhang GL, Wu HT, Zhang ZC, Zheng GS, Luan L, et al. 2009. Alginate-deriving oligosaccharide production by algi- nase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. J. Appl.
Microbial. 106: 161-170.
2. Cho KJ, Lee YS, Ryu BH. 1990. Antitumor effect and immu- nology activity of seaweeds toward sarcoma-180. Bull.
Korean Fish. Soc. 23: 245-352.
3. Choi JH, Kim DW. 1997. Effect of alginic acid-added func- tional drink (HAETOMINI) brown algae (Undaria pinnatifida) on obesity and biological activity of SD rats. Korean J. Life Sci.
7: 361-370.
4. Choi JH, Rhim CH, Kim JY, Yang JS, Choi JS, Byun DS.
1986. Basic studies on the development of diet for the treat- ment of obesity: the inhibitory effect of alginic acid as a dietary fiber on obesity. J. Korean Fish. Soc. 19: 303-311.
5. Joo DS, Cho SY, Lee EH. 1993. Isolation of alginate-degrad- ing bacteria and production of alginate-degrading activity by the bacteria. Korean J. Appl. Microbiol. Biotechnol. 21: 207- 213.
6. Joo DS, Lee JS, Park JJ, Cho SY, Ahn CB, Lee EH. 1995.
Purification and charaterization of the intracellular alginase from Vibrio sp. AL-145. Korean J. Appl. Microbiol. Biotechnol.
23: 432-438.
7. Kennedy L, Mcdowell K, Sutherland IW. 1992. Alginases from Azotobacter species. J. Gen. Microbiol. 138: 2465-2471.
8. Kim HS, Lee CG, Lee EY. 2011. Alginate lyase: structure, property, and application. Biotech. Biopro. Eng. 16: 843-851.
9. Kim IH, Nam TJ. 2005. Effect of polymannuronate on choles- terol contents of liver tissue and feces in rats. J. Korean Soc.
Food Sci. Nutr. 34: 1553-1558.
10. Kim JH, Kim YH, Kim SK, Kim BW, Nam SW. 2011. Properties and industrial applications of seaweed polysaccharides- degrading enzymes from the marine microorganisms. Korean J. Microbiol. Biotechnol. 39: 189-199.
11. Kim OJ, Lee DG, Lee SM, Lee SJ, Do HJ, Park HJ, et al.
2010. Isolation and characteristics of alginate-degrading Methylobacterium sp. HJM27. Korean J. Microbiol. Biotech- nol. 38: 144-150.
12. Kobayashi T, Uchimura K, Miyazaki M, Nogi Y, Horikoshi K.
2009. A new high-alkaline alginate lyase from a deep-sea bacterium Agarivorans sp.. Extrmophiles 13: 121-129.
13. Lange B, Wingender J, Winkler UK. 1989. Isolation and char- acterization of an alginate lyase from Klebsiella aerogenes.
Arch. Microbiol. 152: 302-308.
14. Lee JH, Bae MJ, Kim YC, Nam SW. 2009. Identification and characterization of alginate lyase producing Pseudomonas sp. N7151-6. Korean J. Microbiol. Biotechnol. 4: 350-354.
15. Lee YS, Kim DS, Ryu BH, Lee SH. 1992. Antitumor and immunomodulating effects of seaweeds toward sarcoma-180 cell. J. Korean Soc. Food Nutr. 21: 544-550.
16. Li JW, Dong S, Song J, Li CB, Shen XL, Xie BB, et al. 2011.
Purification and characterization of bifunctional alginate lyase from Pseudoalteromonas sp. SM0524. Marine Drugs. 9: 109- 123.
17. Liyan L, Xialu J, Huashi G, Peng W, Hong G. 2011. Three algi- nate lyases from marine bacterium Pseudomonas fluore- scens. HZJ216: purification and characterization. Appl. Biochem.
Biotech. 164: 305-317.
18. Miyake O, Hashimoto W, Murata K. 2003. An exotype alginate lyase in Sphingomonas sp. Al: overexpression in Escherichia
coli, purification and charaterzation of alginate lyase. Protien Expres. Prurif. 29: 33-41.
19. Nelson A. 1944. A photometric adaptation of the somogyi method for the determination of glucose. J. Biol. Chem. 153:
375-380.
20. Nibu Y, Satoh T, Nishi Y, Takeuchi T, Murata K, Kusakabe I.
1995. Purification and characterization of extraccellular algi- nate lyase from Enterobacter cloacae M-1. Biosci. Biotechnol.
Biochem. 59: 623-637.
21. Richard, AS, Levein RE. 1976. Viscometric assay of bacterial alginase. Appl. Eviro. Microbiol. 31: 896-899.
22. Sawabe T, Ohtsuka M, Ezura Y. 1997. Novel alginate lyases from marine bacterium Alteromonas sp. strain H-4. Carbo- hydr. Res. 304: 69-76.
23. Scot M, Colin GM, David J, Mills L, Brian JB. 1987. Estimation of meiodenthic nematoe diversity by non specialists. Marine Pollution Bulletin 38: 646-649.
24. Stevens RA, Levin RE. 1977. Purification and charateristics of an alginase from Alginovibrio aquatilis. Appl. Environ. Microb.
3: 1156-1161.
25. Sunwoo C, Kim KBWR, Kim DH, Jung SA, Kim HJ, Jeong DH, et al. 2012. Optimization of conditions for the production of alginate-degrading crude enzyme from Vibrio crassostreae
PKA 1002. Korean J. Microbial. Biotechnol. 40: 243-249.
26. Takamistu C, Takashi B, Nozomi H, Mikihiko K. 2008. Purifica- tion and Characterization of alginate lyase from Pseudoaltero- monas sp. Strain No. 1786. J. Appl. Glycosci. 55: 81-88.
27. Takeshita S, Sato N, Igarashi M, Muramatsu T. 1993. A highly denaturant durable alginate lyase from a marine bacterium:
purification and proterties. Biosci. Biotech. Bioch. 57: 1125- 1128.
28. Thiang YW, Preston LA, Schiller NL. 2000. Alginate lyase:
review of major sources and enzyme characteristics structure- function analysis, biological roles and applications. Annu. Rev.
Microbial. 54: 289-340.
29. Uo MH, Joo DS, Joo SY. 2006. Screening and cultivation characteristics of alginate degrading bacteria. J. Korean Soc.
Food Sci. Nutr. 35: 109-114.
30. Yi KW, Shin IS. 2006. Isolation of marine bacterium decom- posing sea tangle (Laminaria japonica) to single cell detritus.
Korean J. Food Sci. Technol. 38: 237-240.
31. Yonemoto Y, Mutata K, Kimura A, Yamaguchi H, Okayama K.
1991. Bacterial alginate lyase: characterization of alginate lyase-producing bacteria and purification of the enzyme. J.
Ferm. Bioeng. 72: 152-157.