454
Copyright © 2016 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
수인성
∙
식품매개바이러스성식중독의대표적인원인체인노 로바이러스는Caliciviridae
과에속하며 외피(envelope)
가없 는단일가닥RNA
바이러스(
약7.6 kb)
로3
개의open reading frame (ORF)
로구성되어있고,
변이가쉬운RNA
바이러스특 성상노로바이러스는5
개의genogroup (GⅠ-GⅤ)
으로분류되 고있으며,
이들중GⅠ
및GⅡ genogroup
의노로바이러스가식중독을일으키는주요원인체로알려져있다
.
특히 노로바이러스
GⅡ genogroup
에는22
종의 유전자형(genotype)
이있는것으로알려져있으며,
그중노로바이러스GⅡ 4
에의한집단감염증환자가2004
년부터2005
년까지여 러나라에서많이발생하였고,
국내에서도노로바이러스GⅡ 4
에의한식중독사고가발생한사례도있다(Bull et al., 2006;
Jung et al., 2015).
노로바이러스감염증은감염량이 매우적어
10
개의노로바해수 중의 수온, 염분 및 pH에 따른 노로바이러스 및 Male-Specific Coliphage 농도변화
김풍호·박용수·박큰바위*·권지영·유홍식
1·이희정
2·김지회
3·이태식
국립수산과학원 식품위생가공과, 1국립수산과학원 서해수산연구소, 2립수산과학원 남해수산연구소, 3국립수산과학원 연구기획과
Change in Concentrations of Human Norovirus and
Male-Specific Coliphage under Various Temperatures, Salinities, and pH Levels in Seawater
Poong Ho Kim, Yong Soo Park, Kunbawui Park*, Ji Young Kwon, Hong Sik Yu
1
, Hee Jung Lee2
, Ji Hoe Kim3
and Tae Seek LeeFood Safety and Processing Research Division, National Institute of Fisheries Science, Busan 46083, Korea
1
West Sea Fisheries Research Institute, National Institute of Fisheries Science, Incheon 22383, Korea
2
South Sea Fisheries Research Institute, National Institute of Fisheries Science, Yeosu 59780, Korea
3
Research and Development Planning Department, National Institute of Fisheries Science, Busan 46083, Korea
Pre- or post-harvest processing is required to mitigate the risk of norovirus infection mediated by shellfish or seafood.
We investigated the environmental resistance of human norovirus (HuNoV) under various conditions of temperature, salinity, and pH in seawater. Male-specific coliphage (MSC) was as the reference virus for all tests. At 4℃, HuNoV GII4 spiked into seawater was continually detected by RT-PCR for 35 days, regardless of salinity or pH level. It main- tained nearly stable concentrations, meaning HuNoV can sustain a viral population in seawater long enough to be accumulated by shellfish and other filter feeders during winter. MSC was also stable at 4℃ although viral infectivity dropped sharply after 28 days. The effects of salinity and pH on MSC were indistinct. At 25℃ the detectable period of HuNoV GII4 by RT-PCR in seawater decreased to about one-third or half of the period at 4℃. High salinity (32 psu) and alkaline pH (8.5) were also unfavorable for sustaining HuNoV abundance at 25℃ in seawater. The resis- tance patterns of MSC to high temperature, high salinity, and alkaline pH were more dramatic and viral infectivity decreased over time, almost in direct proportion to experimental days. MSC was undetectable after 12 days under all salinities and pH levels at 25℃.
Key words: Male specific coliphage, Norovirus, pH, Salinity, Seawater
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2016.0454 Korean J Fish Aquat Sci 49(4) 454-459, August 2016 Received 30 June 2016; Revised 7 July 2016; Accepted 14 July 2016
*Corresponding author: Tel: +82. 51. 720. 2632 Fax: +82. 51. 720. 2619
E-mail address: [email protected]
노로바이러스와 male-specific coliphage 농도변화
455
이러스입자만으로도감염증이유발될수있으며
,
인체감염 시많은수의노로바이러스가분변으로배출되는특징이있다(Koopmans et al., 2002; Blackburn et al., 2004).
노로바이러스식중독은감염증환자의분변에의해식품이오 염되고
,
오염된식품을섭취할경우환자가발생하는순환고리(fecal-oral route)
를형성하고있으며,
특히패류중에서도주 로날것으로섭취가많은굴에의한노로바이러스식중독사 고는호주,
뉴질랜드,
일본등 여러나라에서 보고되고있다(Webby et al., 2007; Simmons et al., 2007; Kroneman et al., 2008; Scallan et al., 2011; Alfano-Sobsey et al., 2012; Iritani et al., 2014).
우리나라에서도노로바이러스에의한식중독사고가사회적 인문제가되고있으며
,
감염증환자의분변에오염된해역에서 생산된패류가노로바이러스의매개체역할을할가능성이커 지고있다(Lee et al., 2010; Moon et al., 2011; Oh et al., 2012;
Shin et al., 2013; Shin et al., 2014).
Male specific coliphage (MSC)
는처리되지않은폐수또는 하수처리장에서처리된후배출되는방출수중에높은농도로 검출되며크기(head diameter)
는25 nm
이고,
단일가닥RNA
구조로염소소독및환경스트레스에대한내성정도가노로바 이러스등다른장관계바이러스의생화학적특성이유사하여 인간분변유래장관계바이러스의오염지표로써주목받고있 다(Burkhardt et al., 1992).
한편
,
노로바이러스라는새로운도전에직면한위생당국들은 오염원에인접한연안에서생산되는패류의안전성확보는전 통적인해역관리만으로는불가능하며수확후처리나가공을 병행하여야한다는점에의견들이있어정화처리(depuration),
고압처리(high hydrostatic pressure)
등의노로바이러스제어 연구가 활발히이루어지고 있으나,
대규모의 패류 가공공장 에적용이가능할정도의연구성과는아직까지없는실정이다(Ueki et al., 2007; Arcangeli et al., 2012).
또한
,
노로바이러스는숙주세포에의한인공배양법이개발되 지않아murine calicivirus (MNV)
나feline calicivirus (FCV)
등과같은대체바이러스(Norovirus surrogates)
로환경중에서 의생존능연구,
고압처리등과같은불활성화연구가수행되어 지고있으나,
인간유래노로바이러스를대상으로해수등수중 환경속에서의노로바이러스생존능등과같은일반특성변화 등에관한연구가매우미미한실정이다(Allwood et al., 2003;
Chen et al., 2005; Seo et al., 2012).
따라서본연구에서는노로바이러스감염증예방을위한새로 운패류유통및패류수확후가공기술개발에기초자료를제 공하고자인간분변에서회수한노로바이러스
GⅡ 4
가패류양 식과저장에사용되는해수의환경조건(
수온,
염분및pH)
에 서의노로바이러스GⅡ 4
및MSC
의농도변화를조사하였다.
재료 및 방법
시험재료 및 조건
해수중의수온
,
염분및pH
조건에따라시간경과에따른노 로바이러스GⅡ 4
및MSC
의농도변화를조사하기위해자연 해수(32 psu, pH 8.5)
에멸균된증류수를첨가하면서염분농도 를10
및20 psu
되게조정하였고, pH
는자연해수에1 N HCl
를 첨가하면서pH 5.5
및7.0
되게조정한후각각의해수량은1 L
로맞추어시험에사용하였다.
염분및
pH
를조정한각각의해수1 L
에인간분변에서회수 한노로바이러스GⅡ 4 (10
3-10
4copies/mL)
및MSC (10
3-10
4PFU/mL)
를첨가한후4℃
및25℃
에서정치하면서일정시간 마다해수15 mL
를채취하여시간경과에따라해수의염분(10, 20, 32 psu)
및pH (pH 5.5, 7.0, 8.5)
조건에서의노로바이러스GⅡ 4
및MSC
농도변화를조사하기위한시료로사용하였다. 해수 중의 MSC 검출
해수중에함유되어있는
MSC
는DeBartolomeis
와Cabelli (1991)
의한천중첩법(agar overlay method)
을사용하였다.
실 험에사용한MSC
숙주세포는Escherichia coli HS (pFamp) R (ATCC 700891)
사용하였으며,
멸균한2.5 mL soft agar
에E.
coli HS (pFamp)R
배양액을0.2 mL
접종하고, 2.5 mL
시료를 첨가한후bottom agar
에중첩시키고, 35℃
에서24
시간배양 후형성된plaque
수를측정하고, MSC
에대한결과표기는100 mL
당PFU (plaque-forming unit)
으로나타내었다.
노로바이러스 RNA 추출
해수중의노로바이러스
RNA
추출은Viral RNA mini kits (QIAGEN)
을사용하였다.
노로바이러스분석시료액500 μL
에6M Guanidine isothiocyanate (GITC)
용액500 μL
첨가하 고,
실온에서5
분간반응시킨후50% ethanol 700 μL
넣고잘흔 들어준다.
이반응용액들을column
에700 μL
넣고8,000 rpm
에서1
분간원심분리하며,
이과정을2
회반복하여노로바이러 스를회수한다.
노로바이러스
RNA
순도를높게추출하기위해서RW1 buf- fer 700 μL
를column
넣고실온에서15
분간방치한후8,000 rpm
에서1
분간원심분리하고,
다시column
에RPE buffer 500 μ L
넣고실온에서15
분간방치한후8,000 rpm
에서1
분간원 심분리하여잔사를제거하고, ethanol
로column
을세척한후70℃ TE buffer 60 μL
를column
에첨가하여원심분리한RNA
를노로바이러스정량분석를위한시료로사용하였다. Realtime RT-PCR에 의한 노로바이러스 정량분석
추출된
RNA
로부터해수중의수온,
염분및pH
조건에따른 노로바이러스농도변화를조사하기위하여다음과같이수행하 였다. Realtime RT-PCR
반응을위하여OneStep RT-PCR kit
(QIAgen, USA)
및RNase inhibitor (Ambion, USA)
시약을 사용하였다.
폴리오바이러스의RNA
를Internal Control RNA (IC, US FDA
제공)
로첨가하여반응이적절히이루어지는지 확인하였으며음성대조군으로RNase-free water
를사용하여 실험의신뢰성을확보하였다.
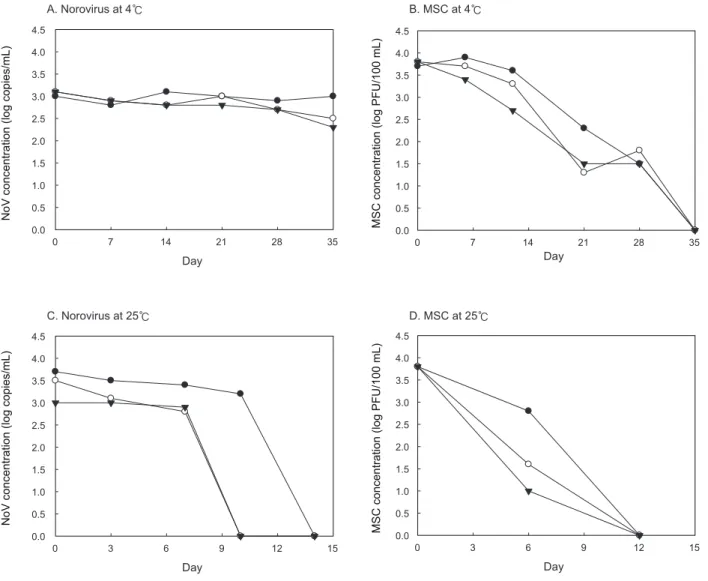
노로바이러스 유전자 검출을 위하여
Table 1
의primer
와probe
를이용하여25× Enzyme mix 0.5 μL, 5× buffer 5 μL, 10× dNTPs 1 μL, RNase inhibitor (5 units/μL) 0.25 μL, 10 μM primer (Forward
및Reverse) 1 μL, 10 μM IC primer (For- ward
및Reverse) 0.5 μL, 10 μM probe (
노로바이러스및IC) 0.5 μL, IC RNA 1 μL,
추출한RNA 5 μL
로반응액을조성한 후,
멸균증류수를첨가하여최종적으로25 μL
의반응액을조 성하였다.
유전자증폭을위해서는
Thermal cycler dice TP800 (Takara, Japan)
를이용하여50℃
에서50
분간reverse transcription
을수 행하고, 95℃
에서15
분간DNA
를변성하였다.
이후95℃
에서10
초, 53℃
에서25
초, 62℃
에서70
초로45 cycles
를반복하였 다.
양성대조군으로노로바이러스RNA (Takara, Japan)
를사 용하고,
음성대조군으로멸균증류수를사용하였다.
결과 및 고찰
수온 및 염분에 따른 노로바이러스와 MSC의 농도변화
패류양식과저장에사용되는자연해수의환경조건에서의노 로바이러스GⅡ 4
와MSC
농도가시간경과에따라변화되는 특성을파악하기위하여자연해수에서노로바이러스는주로동 절기에검출이되고,
하절기에는거의검출이되지않는점을감 안하여시험해수의수온을4℃
와25℃
로고정하였다.
실험에사용한자연해수
(32 psu)
와염분농도(10
및20 psu)
를 조정한조건에서시간경과에따라첨가한노로바이러스GⅡ 4
및MSC
농도변화를Realtime RT-PCR
및plaque assay
를통 해각각정량적으로조사하였으며,
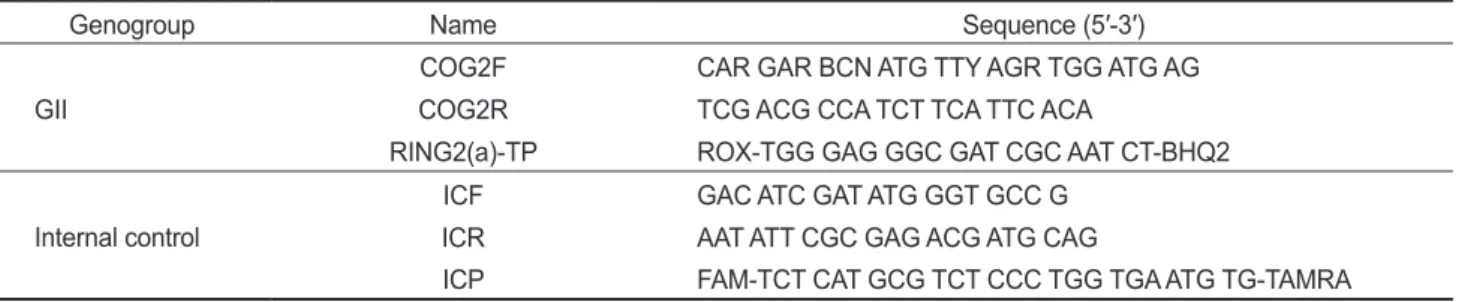
그결과를Fig. 1
에나타내 었다.
4℃
시험구의경우,
염분10, 20
및32 psu
에첨가한노로바이러스
GⅡ 4
는염분농도차이에따라노로바이러스농도의뚜렷한감소차이는없었으며
,
시험기간동안노로바이러스GⅡ
4
가일정농도이상으로지속적으로검출되는것으로볼때4℃
해수에서는염분농도에상관없이노로바이러스는안정적으로 유지되고있는것으로사료된다
.
한편
, 25℃
시험구의경우염분농도조건이10
과20 psu
인경 우에는10
일후노로바이러스GⅡ 4
농도가각각0.7
및0.8 log
감소하여15
일후에는검출한계이하에도달하였고, 32 psu
의 해수중에서는10
일만에노로바이러스GⅡ 4
가검출한계이하 로확인되었다(Fig. 1A
및1C).
한편
, MSC
는4℃
시험구에서염분농도차이에구분없이28
일까지그농도가약1 log
정도감소하였으며35
일후에는검출한계이하로
MSC
농도가급격히낮아졌으며, 25℃
시험구의경우
MSC
는전염분농도조건에서그농도가일수에비례 적으로농도가감소하여12
일이후에는검출한계이하에도달 하였다(Fig. 1B
및1D).
이상의결과
,
노로바이러스와MSC
는해수중의염분농도보 다는수온이높을수록농도가빠르게감소하였으며, 25℃
조건 에서는염분농도가높을수록노로바이러스및MSC
의사멸율 이다소증가하는것으로확인되었다. Lee et al. (2008)
는MNV
의경우실온에서염분농도가높을수록감염력의상실속도가 증가하는것으로보고하였으며, Allwood et al. (2003)
은FCV
는4℃
보다25℃
에서FCV
생존율이감소하였다고보고한연 구결과와유사한경향을나타내었다.
수온 및 pH에 따른 노로바이러스와 MSC의 농도변화
실험에사용한자연해수
(pH 8.5)
와중성및약산성조건에서의노로바이러스
GⅡ 4
및MSC
의농도변화를알아보기위하 여시험구간을pH 5.5, 7.0
및8.5
로설정하고,
해수온도를4℃
와
25℃
로고정하면서시간경과에따른노로바이러스및MSC
농도변화를조사하였으며,
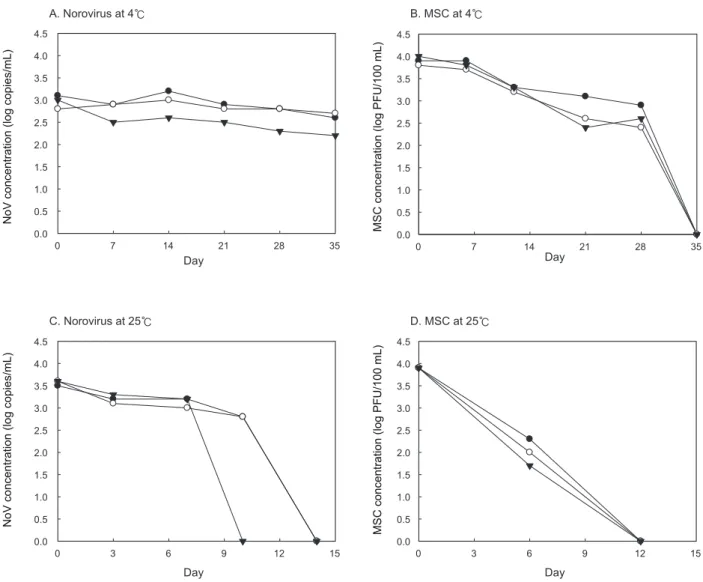
그결과를Fig. 2
에나타내었다.
4℃
시험구의경우pH 5.5
에서는시험기간동안노로바이러스
GⅡ 4
의농도변화는거의없었으며, pH 7.0
및8.5
에서도각 각0.6
및0.8 log
정도로약간감소하여pH
차이에따른노로바이러스
GⅡ 4
농도변화차이는미비하였으나, 25℃
시험구에서
pH 7.0
및8.5
의경우10
일후에는노로바이러스GⅡ 4
가검 출한계이하로도달하였고, pH 5.5
에서는14
일만에노로바이 러스GⅡ 4
가검출되지않았다(Fig. 2A
및2C).
한편
, MSC
는4℃
경우pH 5.5, 7.0
및8.5
시험구에서21
일Table 1. Primers and probes used to detect norovirus
Genogroup Name Sequence (5′-3′)
GII
COG2F CAR GAR BCN ATG TTY AGR TGG ATG AG
COG2R TCG ACG CCA TCT TCA TTC ACA
RING2(a)-TP ROX-TGG GAG GGC GAT CGC AAT CT-BHQ2 Internal control
ICF GAC ATC GAT ATG GGT GCC G
ICR AAT ATT CGC GAG ACG ATG CAG
ICP FAM-TCT CAT GCG TCT CCC TGG TGA ATG TG-TAMRA
노로바이러스와 male-specific coliphage 농도변화
457
후에는각각
1.4, 2.5
및2.3 log
감소하였지만35
일후에는모든 시험구간에서MSC
가검출되지않았으며, 25℃
경우에는pH 5.5, 7.0
및8.5
시험구에서6
일후에는각각1.0, 2.2
및2.8 log
감소하였고, 12
일후에는모든시험구간에서MSC
가검출되지 않았다(Fig. 2B
및2D).
이상의결과
,
노로바이러스와MSC
는해수중의pH
가약산 성(pH 5.5)
보다는약알카리(pH 8.5)
에서농도가빠르게감소 하였고,
수온이높고, pH
가알카리쪽일수록해수중의노로바 이러스와MSC
의농도감소율이더증가하는것으로나타났다.
Cannon et al. (2006)
에의하면MNV
와FCV
는낮은pH
조건(pH 2-4)
에대한내성이높은pH
조건(pH 5-9)
에서보다높았 으며, Seo et al. (2012)
도pH
조건에따른MNV
내성정도를조 사한결과,
중성조건(pH 7.0)
보다는강한산성조건(pH 2)
에서 상대적으로내성율이높았다고보고하였다.
이상의연구결과를종합해보면
,
패류생산해역으로유입된노 로바이러스는해수의수온이높고,
염분의농도가높을수록생존기간이짧아지고
, pH 7.0
이상의영역에서빠르게사멸되는것으로나타났으며
, MSC
는노로바이러스와유사한동태를보 이나고수온조건에대한민감도는노로바이러스보다다소높 은것으로확인되었으며동결과는새로운패류유통,
수확후 패류정화및가공기술개발에기초자료로써활용될수있을것 으로생각된다.
한편
, Shin et al. (2014)
은패류양식장에서의노로바이러스는 수온이낮은겨울철과봄철에주로검출되고있다고보고한연 구결과와본연구의조사결과로볼때겨울철및봄철에패류 생산해역으로유입된노로바이러스는낮은수온으로인해장기 간사멸되지않고,
해수중에존재하여패류에축적될가능성이 높아지고,
패류가노로바이러스식중독을유발시킬수있는원A. Norovirus at 4℃ B. MSC at 4℃
Day
0 7 14 21 28 35
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
Day
0 7 14 21 28 35
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
C. Norovirus at 25℃ D. MSC at 25℃
Day
0 3 6 9 12 15
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
Day
0 3 6 9 12 15
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
A. Norovirus at 4℃ B. MSC at 4℃
Day
0 7 14 21 28 35
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
0 7 14
Day
21 28 350.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
C. Norovirus at 25℃ D. MSC at 25℃
Day
0 3 6 9 12 15
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
Day
0 3 6 9 12 15
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
NoV concentration (log copies/mL) MSC concentration (log PFU/100 mL)
NoV concentration (log copies/mL) MSC concentration (log PFU/100 mL)
NoV concentration (log copies/mL) MSC concentration (log PFU/100 mL)
NoV concentration (log copies/mL) MSC concentration (log PFU/100 mL)
Fig. 1. Concentration analysis of Norovirus GⅡ 4 and male-specific coliphage (MSC) by the different temperature and salinity in seawater.
10 psu (●), 20 psu (○) and 32 psu (▼).
김풍호
ㆍ
박용수ㆍ
박큰바위ㆍ
권지영ㆍ
유홍식ㆍ
이희정ㆍ
김지회ㆍ
이태식458
인식품될수있으므로안전한패류를지속적으로생산하기위 해서는패류생산시기이전부터관리당국의오염원관리조치가 필요할것으로사료된다
.
사 사
이 논문은
2016
년도 국립수산과학원 수산과학연구사업(R2016059)
의지원으로수행된연구이며연구비지원에감사드립니다
.
References
Alfano-Sobsey E, Sweat D, Hall A, Breedlove F, Rodriguez R, Greene S, Pierce A, Sobsey M, Davies M and Led- ford SL. 2012. Norovirus outbreak associated with under-
cooked oysters and secondary household transmission.
Epidemiol Infect 140, 276-282. http://dx.doi.org/10.1017/
S0950268811000665.
Allwood PB, Malik YS, Hedberg CW and Goyal SM. 2003.
Survival of F-specific RNA coliphage, feline calicivirus and
Escherichia coli in water: a comparative study. Appl Envi-
ron Micrbiol 69, 5707-5710.Arcangeli G, Terregino C, De Benedictis P, Zecchin B, Man- frin A, Rossetti E, Magnabosco C, Mancin M and Brutti A.
2012. Effect of high hydrostatic pressure on murine noro- virus in Manila clams. Lett Appl Microbiol 54, 325-329.
http://dx.doi.org/10.1111/j.1472-765X.2012.03211.x.
Blackburn, BG, Craun GF, Yoder JS, Hill V, Calderon RL, Chen N, Lee SH, Levy DA and Beach MJ. 2004. Surveillance for waterborne-disease outbreaks associated with drinking water-United States. 2001-2002. MMWR Surveill Summ
A. Norovirus at 4℃ B. MSC at 4℃
Day
0 7 14 21 28 35
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
Day
0 7 14 21 28 35
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
C. Norovirus at 25℃ D. MSC at 25℃
Day
0 3 6 9 12 15
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
Day
0 3 6 9 12 15
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
Day
0 7 14 21 28 35
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5
0 7 14
Day
21 28 350.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5
C. Norovirus at 25℃ D. MSC at 25℃
Day
0 3 6 9 12 15
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
Day
0 3 6 9 12 15
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
NoV concentration (log copies/mL) MSC concentration (log PFU/100 mL)
NoV concentration (log copies/mL) MSC concentration (log PFU/100 mL)
NoV concentration (log copies/mL) MSC concentration (log PFU/100 mL)
NoV concentration (log copies/mL) MSC concentration (log PFU/100 mL)
Fig. 2. Concentration analysis of Norovirus GⅡ 4 and male-specific coliphage (MSC) by the different temperature and pH in seawater. pH 5.5 (●), pH 7.0 (○) and pH 8.5 (▼).
노로바이러스와 male-specific coliphage 농도변화
459
53, 23-45.
Burkhardt W 3rd, Watkins WD and Rippey SR. 1992. Survival and replication of male-specific bacteriophages in mollus- can shellfish. Appl Environ Microbiol 58, 1371-1373.
Bull RA, Tu ET, Mclver CJ, Rawlinson WD and White PA.
2006. Emergence of a new norovirus genotype Ⅱ. 4 vari- ant associated with global outbreaks of gastroenteritis. J Clin Microbiol 44, 327-333.
Cannon JL, Papafragkou E, Park GW, Osborne J, Jaykus LA and Vinje J. 2006. Surrogate for the study of norovirus sta- bility and inactivation in the environment: A comparison of murine norovirus and feline calicivirus. J Food Protect 69, 2761-2765.
Chen H, Hoover DG and Kingsley DH. 2005. Temperature and treatment time influence high hydrostatic pressure inactiva- tion of feline calicivirus, a norovirus surrogate. J Food Pro- tect 68, 2389-2394.
DeBartolomeis J and Cabelli VJ. 1991. Evaluation of an Esch-
erichia coli of F male-specific bacteriophages. Appl Environ
Microbiol 57, 1301-1305.Iritani N, Kaida A, Abe N, Kubo H, Sekiguchi JI, Yamamoto SP, Goto K, Tanaka T and Noda M. 2014. Detection and genetic characterization of human enteric viruses in oyster- associated gastroenteritis outbreaks between 2001 and 2012 in Osaka City, Japan. J Med Virol 86, 2019-2025. http://
dx.doi.org/10.1002/jmv.23883.
Jung S, Hwang BM, Jeong HJ, Chung GT, Yoo CK, Kang YH and Lee DY. 2015. Occurrence of Norovirus GⅡ 4 Sydney variant-related outbreaks in Korea. Osong Public Health Res Perspect 6, 322-326.
Koopmans M, Bonsdor CH, Vinje J, Medici D and Monroe S.
2002. Foodborne viruses. FEMS Microbiol 26, 187-205.
Kroneman A, Verhoef L, Harris J, Vennema H, Duzier E, Van Duynhoven Y, Gray J, Iturriza M, Böttiger B, Falken- horst G, Johnsen C, von Bonsdorff CH,Maunula L, Kuusi M, Pothier P, Gallay A, Schreier E, Höhne M, Koch J, Szücs G, Reuter G, Krisztalovics K, Lynch M, McKeown P, Foley B, Coughlan S, Ruggeri FM, Di Bartolo I, Vainio K, Isak- baeva E, Poljsak-Prijatelj M, Grom AH, Mijovski JZ, Bosch A, Buesa J, Fauquier AS, Hernandéz-Pezzi G, Hedlund KO and Koopmans M. 2008. Analysis of integrated virological and epidemiological reports of norovirus outbreaks collect- ed within the foodborne viruses in Europe Network from 1 July 2001 to 30 June 2006. J Clinical Microbiol 46, 2959- 2965. http://dx.doi.org/10.1128/JCM. 00499-08.
Lee TS, Oh EG, Yoo HD, Ha KS, Yu HS, Byun HS and Kim JH. 2010. Impact of rainfall events on the bacteriological water quality of the shellfish growing area in Korea. Korean J Fish Aquat Sci 43, 406-414. http://dx.doi.org/10.5657/
KFAS.2010.0406.
Lee JE, Zoh KD and Ko GP. 2008. Inactivation and UV disinfec- tion of murine norovirus with TiO2 under vatious environ-
mental conditions. Appl Environ Micrbiol 74, 2111-2117.
Moon A, Hwang IG and Choi WS. 2011. Prevalence of noro- viruses in oyster in Korea. Food Sci Biotechnol 20, 1151- 1154.
Oh EG, Yoo HD, Yu HS, Ha KS, Shin SB, Lee TS, Lee HJ, Kim JH and Son KT. 2012. Removal of fecal indicator bacteria from bivalves under natural and electrolyzed water. Ko- rean J Fish Aquat Sci 45, 11-16. http://dx.doi.org/10.5657/
KFAS.2012.0011.
Scallan E, Hoekstra RM, Angulo FJ and Tauxe RV. 2011. Food- borne illness acquired in the United States—Major patho- gens. Emerg Infect Dis 17, 7-15. http://dx.doi.org/10.3201/
eid1701.P11101.
Seo K, Lee JE, Lim MY and Ko G. 2012. Effect of temperature, pH and NaCl on the inactivation kinetics of murine norovi- rus. J Food Protect 69, 2761-2765.
Shin SB, Oh EG, Yu HS, Son KT, Lee HJ, Park JY and Kim JH 2013, Genetic diversity of noroviruses detected in Oyster in Jinhae bay, Korea. Food Sci Biotechnol 22, 1453-1460.
http://dx.doi.org/10.1007/s10068-013-0237-z.
Shin SB, Oh EG, Lee HJ, Kim YK, Lee TS and Kim JH. 2014.
Norovirus quantification in oysters Crassostrea gigas Col- lected from Tongyeoung, Korea. Korean J Fish Aquat Sci 47, 501-507. http://dx.doi.org/10.5657/KFAS.2014.0501.
Simmons G, Garbutt C, Hewitt J and Greening G. 2007. A New Zealand outbreak of norovirus gastroenteritis linked to the consumption of imported raw Korean oysters. The New Zealand Medical J 120:U2773.
Ueki Y, Shoji M, Suto A, Tanabe T, Okimura Y, Kikuchi Y, Saito N, Sano D and Omura T. 2007. Persistence of cali- civiruses in artificially contaminated oysters during depura- tion. Appl Environ Microbiol 73, 5698-5701. http://dx.doi.
org/10.1128/AEM.00290-07.
Webby RJ, Carville KS, Kirk MD, Greening G, Ratcliff RM, Crerar SK, Dempsey K, Sarna M, Stafford R, Patel M and Hall G. 2007. Internationally distributed frozen oyster meat causing multiple outbreaks of norovirus infection in Austra- lia. Clinical Infectious Disease 44, 1026-1031.