Streptomyces coelicolor A3(2) 로 부터 β-Glucosidase 유전자 클로닝 및 재조합 효소의 특성
김재영·김봉규1·이용섭*·강창수2·안중훈1·임융호1
호서대학교 한방화장품과학과 기초과학연구소, 1건국대학교 생명공학과 생명/분자정보학센터
2호서대학교 생명과학과
Cloning of β-Glucosidase Gene from Streptomyces coelicolor A3(2) and Characterization of the Recom- binant β-Glucosidase Expressed in Escherichia coli. Kim, Jaeyoung, Bongkyu Kim1, Changsoo Kang2, Yongsub Yi*, Joong-hoon Ahn1, and Yoongho Lim1. Department of herbal medicine, Basic Science Institute, Hoseo University, Asan 336-795, Korea, 1BMIC, Division of Bioscience and Biotechnology, Konkuk University, Seoul 143-701, Korea, 2Department of Biological Science, Hoseo University, Asan 336-795, Korea − The β-glu- cosidase gene from Streptomyces coelicolor A3(2) was cloned and expressed in Escherichia coli. The ORF consisted of 1377 nucleotides encoding 51 kDa in a predicted molecular weight. Effects of pH indicated that the β-glucosidase showed similar activity using α-pNPG(ρ-nitrophenyl-α-D-glucopyranoside), β-pNPG(ρ- nitrophenyl-β-D-glucopyranoside), and β-pNPF(ρ-nitrophenyl-β-D-fucopyranoside) at range of pH 3 to 10, and high activity using β-pNPGA (ρ-nitrophenyl-β-D-galactopyranoside) from pH 5 to 10, especially, 3.3 times higher activity at pH 9. Effects of temperature indicated that the β-glucosidase showed low activity using α-pNPG, β-pNPG, and β-pNPF from 20oC to 70oC, and increased activity using β-pNPGA from 30oC to 50oC, 1.8 times higher activity at 50oC than at 30oC. According to activity determination of other substrates, the enzyme was active on daidzin, genistin, and glycitin, inactive on esculin and apigenin-7-glucose. The EDTA and DTT as reducing agents inhibited β-glucosidase activity, but SDS and mercaptoethanol did not inhibit. Monovalent or divalent metal ions such as MnSO4, CaCl2, KCl, and MgSO4 did not inhibited β-glu- cosidase activity. CuSO4 and NaCl showed low inhibition, and ZnSO4 inhibited 3.3 times higher than control.
Key words: β-glucosidase, deglycosylation, Streptomyces coelicolor A3(2), substrate specificity
서 론
Glycosyl hydrolase family는 큰 유전자군으로 현재 114개 의 family로 분류되어 있으며 glycosyl hydrolase family 1 group에 β-glucosidase(EC 3.2.1.21)이 속해있다[4, 12].
지금까지 밝혀진 β-glucosidase (BGL)의 생리적인 역할은 식물의 경우 식물호르몬이나 열매의 향기형성을 활성화하는 데 관여하거나, 식물병원균의 저항성 기작에 관여하는 것으 로 연구되고 있으며[7, 11] 그 밖에 동물, 미생물 등 많은 생 물종에 존재하며, 기질로 aryl과 alkyl-β-D-glucosides를 사 용하는 등, 폭 넓은 기질특이성을 가지는 것으로 보고되고 있다. 이러한 BGL 유전자의 역할은 cellulose를 당화하는 과 정에 관여하는 것으로 보고되고 있으나, 이 효소의 폭 넓은 기질특이적 특성이 in vivo 상태에서 어떠한 역할을 하는지 에 대한 지식은 아직 미흡하다[23, 26]. 한편, 미생물의 당화 과정에 관여하는 효소로 β-glucosidase의 역할을 보고하고
있는데, cellulose 분해를 못하는 미생물에서도 이 효소가 보 고되고 있다[1, 8, 16]. 이러한 BGL는 아미노산이 450개인 type과 아미노산이 800개인 type로 각각 나누고 있으며, Type I에 속하는 BGL은 세균에서 주로 발견되고 있고, type II는 세균과 곰팡이에서 함께 발견되고 있다. 지금까지 Trichoderma, Clostridium, Bacillus, Pyrococcus, Bifidobac- terium 및 Aspergillus niger, Sporotichum cellulopolum, Bifidobacterium 등에서 BGL 효소가 보고되고 있다[5, 10, 14, 19, 24, 25, 29, 30].
BGL 효소는 폭 넓은 기질특이성을 통하여 식품, 화학, 세 척제 및 직물업 등 전분을 소당으로 분해하는데 이용되며, 특히 와인 생산공정에서 유용하게 사용되고 있다. 이러한 BGL 효소를 이용하여 glycoside 유도체로부터 glucose를 유 리하는 연구가 진행되고 있는데, 유리작용으로 생성된 물질 은 인체에 생리적 활성을 갖고 있어 고부가 가치가 있다[6, 17, 27].
본 연구에서 사용한 Streptomyces coelicolor A3(2)는 그 람양성 세균으로 streptomycin 생산균주이다. 현재 전체유전 자에 대한 유전정보가 보고되어 활용되고 있으며, 분석결과 glycosyl hydrolase family 1으로 추정되는 유전자가 5개로
*Corresponding author
Tel: 82-41-540-5979, Fax: 82-41-541-5979 E-mail: [email protected]
보고되고 있다[2, 4].
따라서, 본 연구는 S. coelicolor A3(2)의 유전정보를 바탕 으로 β-glucosidase 유전자를 분리하고, 그 특성을 조사하여 물질전환을 위한 응용분야에 활용하고자 한다.
재료 및 방법 사용균주, 배지 및 시약
본 연구에서 사용한 Streptomyces coelicolor A(3)2 균주 는 한국표준균주은행에서 분양받아 사용하였고, 배양을 위 하여 nutrient broth (Difco. USA)에서 7일간 28oC에서 배 양하였다. 활성측정에 사용된 기질은 ρ-nitrophenyl-α-D- glucopyranoside(α-pNPG), ρ-nitrophenyl-β-D-fucopyrano- side(β-pNPF), ρ-nitrophenyl-β-D-glucopyranoside(pNPG), ρ-nitrophenyl-β-D-galactopyranoside(β-pNPGA), Daidzin, Genistin, Glycitin, Esculin, Apigenin-7-glucose은 Sigma사
(USA)로부터 구입하여 사용하였고, 금속이온 및 환원제로
사용된 MnSO4, MgSO4, CuSO4, ZnSO4, CaSO4, NaSO4, KCl, EDTA, SDS, DTT, mercaptoethanol 또한 Sigma사로 부터 구입하여 사용하였다.
β-Glucosidase 유전자 분리 및 발현벡터 제작
S. coelicolor A(3)2 균주의 유전자 DNA를 추출한 뒤 sense primer : ATGAGTGAGTACCCCGGTTTCCCG, anti- sense primer : TCACCCGCGGTTGGCGGCGATCATC를 이용하여 PCR반응(95oC 1분, 58oC 1분, 72oC1분 30초 동 안 35회 증폭)을 하였다. 증폭된 PCR 산물을 pGEM-T Easy vector(Promega, USA)에 삽입하여 염기서열을 확인한 후, 발현 벡터인 pET15b(Invitrogen, USA)에 ligation하고, 재조합 단백질 발현용 host인 E. coli BL21(DE3) pLys (Novagen, Inc.)로 형질전환 시켰다.
β-Glucosidase의 발현 및 정제
발현을 위한 E. coli BL21(DE3) pLys 형질전환균을 10 mL LB 배지에 종균접종 후, 종균 배양액을 1 L LB배지 (Difco Lab., USA)에 접종하여 abs 600 nm에서 O.D. 값이 0.8 이상일 때 최종농도 0.1 mM가 되도록 IPTG를 첨가하여 16시간 동안 배양하였다. 배양액은 5000 rpm에서 30분간 원 심분리 한 뒤 균체를 20 mM Tris-HCl(pH 8.0) 10 mL에 현 탁하였다. 발현된 효소를 추출하기 위하여 균체현탁액을 초 음파 처리한 후 5000 rpm에서 10분간 원심분리하여 상등액 을 조추출액으로 사용하였다. 조추출액은 RC-membrane filter Minisart RC 25(Viva science, Germany)로 여과하여 사용하였다. Histrap FF column은 Tris-buffer(20 mM pH 8.0)로 평형을 맞춘 뒤 여과한 조추출액을 column에 넣고 Tris-buffer(20 mM pH 8.0)로 washing한 후, 500 mM im- idazole이 첨가된 Tris buffer(20 mM pH 8.0)를 이용하여
1 mL min-1의 속도로 선형농도구배를 가하여 용출하였다. 용 출된 재조합 β-glucosidase는 12% SDS-PAGE를 이용하여 확인하였으며[18], imidazole은 투석하여 제거하였다.
Imidazole이 제거된 β-glucosidase는 한외여과장치 (Centri- plus YM, MILLIPORE, USA)를 이용하여 농축하고 sodium acetate buffer(50 mM pH 6.0)로 투석한 다음 4oC에 보관 하여 활성측정에 사용하였다.
β-Glucosidase의 단백질 정량
농축된 단백질의 정량은 Bradford method를 이용하여 흡 광도 595 nm에서 측정하였으며[3], bovine serum albumin
(BSA)를 표준단백질로 사용하였다.
β-Glucosidase 활성 측정
β-Glucosidase의 활성은 ρ-nitrophenyl-β-D-glucopyrano- side(ρNPG)를 기질로 이용하여 측정하였다[20]. 반응액은 25 µL의 β-glucosidase와 25 µL의 ρNPG(10 mM)를 넣어준 뒤 sodium acetate buffer(50 mM pH 6.0)를 넣어 100 µL로 맞 추었으며 30oC에서 15분간 반응시킨 후 NaOH-glycine buffer(0.4 M pH 10.8) 100 µL를 첨가하여 반응을 정지시킨 다음 405 nm에서 흡광도를 측정하여 효소활성을 결정하였 으며, 표준물질은 ρ-nitrophenol을 사용하였다. β-Glucosidase 의 1 unit는 ρ-nitrophenyl-β-D-glucopyranoside에서 ρ-nitro- phenol(ρNP) 1 µmol을 1분 동안 생산 하는데 필요한 양으 로 정의하였다. 기질에 사용된 flavonoids는 HPLC(Varian, USA)를 이용하여 분석하였으며, Varian C18 역상 column을 이용하여 유속 1 mL/min으로, 이동상 A: 0.1% formic acid, B: acetonitrile로 245 nm에서 65분간 분석하였다.
β-Glucosidase 특성 분석
정제된 β-glucosidase를 β-glucosidase assay 법을 통해 측정하였으며 시간과 온도, pH를 단계적으로 조사하였다.
효소활성 최적반응시간은 10-30분까지 10분 간격으로 측 정하였으며, 최적반응온도는 buffer를 pH 5, 6으로 구분하여 20-70oC까지 10oC 간격으로 측정하였고, 최적활성 pH는 pH 3-10까지 pH 1 간격으로 측정하였다. 사용한 buffer는 pH 범위에 따라서 pH 3.0-5.0(50 mM, sodium acetate buffer), pH 6.0-8.0(50 mM, Tris-HCl buffer) 및, pH 9.0-10.0(50 mM, glycine-NaOH buffer)이며, 위의 모든 실험은 3회 반 복하여 실험하였다. 기질에 대한 효소의 최적반응온도와 pH 를 측정하였으며 기질로는 α-pNPG, β-pNPF, pNPG, β-
pNPGA를 사용하였다. 한편, 금속이온 등에 대한 효소활성
을 측정하였으며, 반응액에 10 mM이 되도록 MnSO4, MgSO4, CuSO4, ZnSO4, CaSO4, NaSO4, KCl, EDTA, SDS, DTT 및 mercaptoethanol을 첨가한 뒤 실온에서 15분 간 반응시켰다.
결과 및 고찰 β-Glucosidase 유전자선발, 발현 및 정제
Streptomyces coelicolor A(3)2 균주에서 분리한 유전자 DNA를 이용하여 PCR하여 1.5 kb의 PCR 산물을 증폭하였 다. 증폭된 PCR 산물의 염기서열을 조사하고, NCBI의 BLAST search를 하여 S. coelicolor A(3)2의 β-glucosidase 유전자의 염기서열임을 확인하고, 발현벡터 pET15b에 cloning 하였다. 발현벡터에 삽입이 확인된 plasmid pET15bScBGL 는 E. coli BL21(DE3) pLys에 형질전환시켜 재조합 단백질 발현에 사용하였다.
형질전환 된 E. coli BL21(DE3) pLys를 LB 배지에 접종 하여 발현시켰다. 적정 IPTG 농도를 조사하기 위하여 0, 0.01, 0.1, 0.5, 1 mM 농도로 배지에 첨가하여, 0.1 mM
IPTG 농도에서 가장 높게 발현되는 것을 확인하였고, 25oC
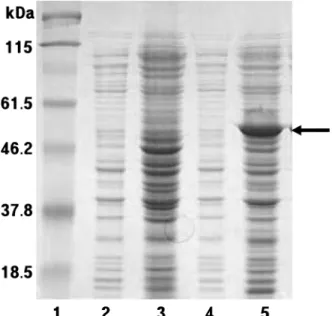
에서 가장 높게 발현되었다(Fig. 1). 이러한 β-glucosidase는 1377염기서열의 51 kDa 분자량의 단백질로 분리하기 위해 서 상등액을 여과하고 Histrap column을 이용하여 정제하였 다. pNPG 활성분획은 12% SDS-PAGE를 통해 확인 하였으 며 투석을 통해 imidazole을 제거하였다(Fig. 2).
β-Glucosidase의 최적활성조건
부분정제된 β-glucosidase(ScBGL1)의 활성은 β-glucosid- ase assay 법으로 조사하였으며 반응buffer의 조건에 따라 활 성이 다르게 나타났으며, 기질로 pNPG를 이용하였다. 최적
반응시간은 10-30분까지 10분 간격으로 측정한 결과 30분 이후에는 활성의 변화가 크게 나타나지 않았고 안정성이 유 지되었으며 15분 동안 반응시킬 때 최적활성을 나타냈다. 온 도에 따른 활성은 pH 5일 때 20oC, pH 6 일 때 60oC에서 가장 높은 활성을 나타냈다. 이러한 활성차이는 낮은 온도 에서 더욱 뚜렷한 차이를 보였다(Fig. 3). pH 변화에 따른 활성은 pH 3과 pH 9 이상에서는 낮은 활성을 나타냈으며, pH 7에서 가장 높은 활성을 나타냈다(Fig. 3).
β-Glucosidase의 기질특성
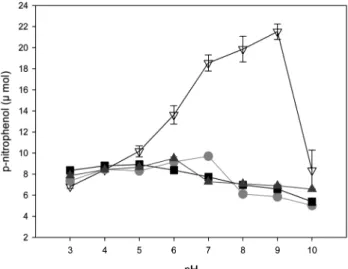
기질에 대한 β-glucosidase의 pH 및 온도에 따른 특성을 조사하였다. α-pNPG, β-pNPG, β-pNPF는 pH 3-10 까지 비 슷한 활성을 나타냈으며, α-pNPG가 pH 7에서 약간 높은 활 성을 보였다(Fig. 4). β-pNPGA는 다른 기질보다 pH 5-9까 지 높은 활성을 나타냈으며, 특히 pH 9에서는 약 3배 이상 높은 활성을 나타냈다(Fig. 4). α-pNPG, β-pNPG, β-pNPF 에 대한 온도에 따른 활성변화는 20-70oC까지 큰 변화폭을 보이지 않고 있으며, 60oC에서 β-pNPF의 활성이 다소 높게 나타난 반면, β-pNPGA는 30-50oC까지 활성이 증가하여 50oC에서 가장 높은 활성을 보였다(Fig. 5). 다른 화학구조 를 가지는 기질과의 활성을 조사하기 위하여 당이 결합된 flavonoids를 이용한 기질특이성을 조사하였으며 daidzin, genistin, glycitin, esculin, apigenin-7-glucose 등을 사용하 였다. β-pNPGA에 대한 상대활성은 daidzin 17%, glycitin 12%, genistin 8% 순으로 나타났으며 esculin과 apigenin-7- glucose 두 기질은 활성을 나타내지 않았다(Table 1).
금속이온 및 기타 영향
β-Glucosidase 활성은 EDTA와 DTT에서는 억제되었으며, SDS와 mercaptoethanol에서는 대조구와 비슷하였다(Fig. 6).
Fig. 1. Expression of recombinant β-glucosidase gene from Streptomyces coelicolor A3(2) in E. coli. Samples were run in 12% SDS-PAGE. lane 1, molecular size markers; lane 2, no induction of E. coli; lane 3, E. coli lysate without recombinant β- glucosidase after induction by 0.01 M IPTG; lane 4, E. coli lysate with recombinant β-glucosidase before induction by 0.01 M IPTG;
lane 5, soluble protein after induction.
Fig. 2. SDS-PAGE of the purified recombinant β-glucosidase in E. coli by liquid chromatography. lane 1, molecular size markers; lane 2, before expression; lane 3, after expression; lane 4- 10, active fractions.
1가 및 2가 금속이온으로 MnSO4, CaCl2, KCl, MgSO4에서 는 효소활성이 증가하였으며, 특히 Mn 이온이 효소의 활성 을 약 3.5배 증가시키는 것으로 나타났다. CuSO4, NaCl은 효소활성을 저해하였으며 특히, ZnSO4는 β-glucosidase 에 대하여 높은 저해활성을 나타냈다(Fig. 6).
Streptomyces 속에서 β-glucosidase(BGL)의 역할에 대해 서 밝혀진 것이 많지는 않지만 주로 BGL 효소의 활성에 초 점이 맞추어져 연구가 이루어지고 있다[13, 15, 21, 22, 28].
같은 family에 속하는 glycosyl hydrolases는 일반적으로 유전자의 염기서열과 발현된 단백질의 구조가 유사한 경우, 특성 또한 유사한 것으로 보고하고 있다[28]. ScBGL1에서 보이는 특성은 다른 종의 β-glucosidase특성과 비슷한 경향
을 보이고 있다. Glycosyl hydrolase family 1에 속하는 유 전자는 기질특이적 반응에 따라 (1) aryl β-glucosidases, (2) cellobiose와 함께 oligosaccharides만을 가수분해하는 cello- Fig. 3. Effects of temperature (A) and pH (B) on the recombinant β-glucosidase activity from Streptomyces coelicolor A3(2) in E.
coli. □, pH 5.0; ●, pH 6.0.
Fig. 4. Effects of substrate on the activity of recombinant β- glucosidase from Streptomyces coelicolor A3(2) in E. coli at different pHs. ●, ρ-nitrophenyl-α-D-glucopyranoside; ■, ρ-nitro- phenyl-β-D-glucopyranoside; ▲, ρ-nitrophenyl-β-D-fucopyrano- side; ▽, ρ-nitrophenyl-β-D-galcotopyranoside.
Fig. 5. Effects of substrates on the activity of recombinant β- glucosidase from Streptomyces coelicolor A3(2) in E. coli at different tempatures. ●, r-nitrophenyl-α-D-glucopyranoside; ■, ρ-nitrophenyl-β-D-glucopyranoside; ▲, ρ-nitrophenyl-β-D-fuco- pyranoside; ▽, ρ-nitrophenyl-β-D-galcotopyranoside.
Table 1. Relative activity of recombinant β-glucosidase from Streptomyces coelicolor A3(2) in E. coli on various flavonoid substrates.
Flavonoid substrates Relative activity(%)
β-pNPG 100
Daidzin 17
Genistin 8
Glycitin 12
Esculin 0
Apigenin-7-glucose 0
biases, (3) 위 두 가지 모두를 기질로 사용하는 β-glucosidases 로 구별하고 있다[6]. 또한 β-glucosidase는 기질특이성에 따 라 β-glucosidase A, B로 나누기도 한다[9]. ScBGL1의 경 우 nitro group을 가지는 기질에서 활성을 나타내는데, 다른 종의 BGL 유전자의 경우 nitro group을 가지는 기질에서 활 성반응을 보이지 않는 경우도 보고되고 있다[17]. ScBGL1 의 경우 cellobiose와 CMC에서 활성을 나타내지 않고 있어 1 group에 속한다고 보이나, β-glucosidase의 기질선택 폭이 넓은 점을 생각 한다면 보다 다양한 기질선택의 조사가 필 요하다고 본다. 이러한 기질선택의 다양성은 flavonoids를 이 용한 기질특이성 조사에서도 보이는데, 다른 종의 BGL효소 의 경우도 flavonoids 기질선택에서 다양한 선택을 보이고 있다. BGL를 물질전환을 위한 소재로 활용하기 위해서는 보 다 다양한 효소의 개발이 필요하다고 본다. 최근 당화관련 효소의 개발은 바이오에너지 개발에 맞추어 전처리단계에서 우수한 효소개발의 필요성이 요구되고 있다. 효소를 이용한 자원의 전처리 기술은 많은 발전을 이루었지만 우수한 효소 개발을 통한 생산비의 절감은 방향이 바이오에너지 개발의 한 축이기도 하다[30, 31]. 이러한 측면에서 우수한 효소의 개발은 물질전환 뿐만 아니고 에너지자원 개발의 소재로도 사용이 가능하다.
요 약
Streptomyces coelicolor A3(2)의 β-glucosidase 유전자를 분리하여 대장균에서 발현하여 특성을 조사하였다. 최적 활 성을 나타내는 온도는 pH 5에서는 20oC, pH 6에서는 60oC 에서 높은 활성을 나타냈다. pH에 따른 활성은 pH 3 이하 와 pH 9 이상의 범위에서는 낮은 활성을 나타냈으며 pH 7 에서 가장 높은 활성을 나타냈다.
α-pNPG(ρ-nitrophenyl-α-D-glucopyranoside), β-pNPG (ρ-
nitrophenyl-β-D-glucopyranoside), β-pNPF(ρ-nitrophenyl-β- D-fucopyranoside)는 pH 3-10까지 비슷한 활성을 나타냈으 며, α-pNPG가 pH 7에서 다소 높은 활성을 보였다. β-
pNPGA는 pH 5-9까지 높은 활성을 나타냈으며, 특히 pH 9
에서 3배 이상의 높은 활성을 나타냈다. 기질 α-pNPG, β- pNPG, β-pNPF의 온도에 따른 활성변화는 β-pNPF의 활성 이 60oC에서 증가하였고, β-pNPGA는 30-50oC까지 활성이 증가하여 50oC에서 최대활성을 나타내었다. 당화 flavonoid 를 이용한 기질특이성의 상대활성은 daidzin, glycitin, genistin, 순으로 나타났으며 esculin과 apigenin-7-glucose는 기질로 사용하지 않았다. β-Glucosidase 활성은 EDTA, DTT에 의해 억제되었으며, MnSO4, CaCl2, KCl, MgSO4 에 의해 증가하였고, 특히 Mn이온에 의해 증가하였다.
CuSO4, NaCl에 의해 효소활성이 저해되었으며, 특히 ZnSO4
의 경우 효소활성이 강하게 억제되었다.
감사의 글
본 연구는 농촌진흥청 바이오그린 21(과제번호:
20080401034052)에 의해 수행되었으며 연구비 지원에 감사
드립니다.
REFERENCES
1. Barras, F., J. P. Chambost, and M. Chippaux. 1984. Cello- biose metabolism in Erwinia: genetic study. Mol. Gen.
Genet. 197: 486-490.
2. Bentley, S. D., K. F. Chater, A. M. Cerdeno-Tarraga, G. L.
Challis, N. R. Thomson, K. D. James, D. E. Harris, M. A.
Quail, H. Kieser, D. Harper. et al. 2002. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature. 417: 141-147.
3. Bradford, M. M. 1976. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:
248-254.
4. Coutinho, P. M. and B. Henrissat. Carbohydrate-Active Enzymes Server. http://afmb.cnrs-mrs.fr/CAZY.
5. Dan, S., I. Marton, M. Dekel, B. Bravdo, S. He, S. G.
Withers, and O. Shoseyov. 2000.Cloning, expression, char- acterization, and nucleophile identification of family 3, Aspergillus niger β-Glucosidase. J. Biol. Chem. 275: 4973- 4980.
6. Enari, T. M. and M. L. Niku-paavola. 1987. Enzymatic hydrolysis of cellulose: Is the current theory of the mecha- nism of hydrolysis valid? Crit. Rev. Biotechnol. 5: 67-87.
7. Estruch, J. J., D. Chriqui, K. L. Grossmann, J. Schell, and A.
Spena. 1991. The plant oncogene rolC is responsible for the release of cytokinins from glucoside conjugates. EMBO J.
10: 2889-2895.
8. Grabnitz, F. and W. L. Staudenbauer. 1988. Characterization Fig. 6. Effects of metal ions and other chemical reagents on the
activity of recombinant β-glucosidase from Streptomyces coeli- color A3(2) in E. coli.
of two β-glucosidase genes from Clostridium thermocellum.
Biotechnol. Lett. 10: 73-78.
9. Grabnitz, F., M. Seiss, K. P. Rucknagel, and W. L.
Staudenbauter. 1991. Structure of the β-glucosidase gene A of Clostridium thermocellum. Eur. J. Biochem. 200: 301- 309.
10. Grimaldi, A., E. Bartowsky, and V. Jiranek. 2005. Screening of Lactobacillus spp. and Pediococcus spp. for glucosidase activities that are important in oenology. J. Appl. Microbiol.
99: 1061-1069.
11. Gunata, Y. Z., C. L. Bayonove, R. L. Baumes, and R. E.
Cordonier. 1985. The aroma of grapes I. Extraction and determination of free and glycosidically bound fractions of some grape aroma components. J. Chromatogr. 331: 83-90.
12. Henrissat, B. 1991. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 280:
309-316.
13. Jang, H. D. and K. S. Chang. 2005. Thermostable cellulases from Streptomyces sp.: scale-up production in a 50-l fer- menter. Biotechnol. Lett. 27: 239-242.
14. Jenkins, J. L., L. L. Leggio, G. Harris, and R. Pickersgill.
1995. β-Glucosidase, β-galactosidase, family A cellulases, family F xylanases and two barley glycanases form a superfamily of enzymes wit 8-fold ββ/αα architecture and with two conserved glutamates near the carboxy-terminal ends of ββ-strands four and seven. FEBS Letters 362: 281- 285.
15. Jiresová, M., Z. Dobrová, J. Náprstek, P. Rysavý, and J.
Janecek. 1983. Induction of beta-D-glucosidase in Streptomyces granaticolor. Folia Microbiol. (Praha) 28: 379-385.
16. Kadam, S., A. L. Demain, J. Millet, P. Biguin, and J. P.
Aubert. 1988. Molecular cloning of a gene for a thermostable β-glucosidase from Clostridium thermocellum into Escherichia coli. Enzyme Microb. Technol. 10: 9-13.
17. Kaur, J., B. S. Chadha, B. A. Kumar, G. S. Kaur, and H. S.
Saini. 2007. Purification and characterization of β-glu- cosidase from Melanocarpus sp. MTCC3922. Electronic Journal of Biotechnology 10: 260-270.
18. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 227:
680-685.
19. Otieno, D. O., J. F. Ashton, and N. P. Shah. 2005. Stability of β-glucosidase activity produced by Bifidobacterium and Lactobacillus spp. in fermented soymilk during processing and storage. J. Food Sci. 70: 236-241.
20. Parry, N. J., D. E. Beever, I. Vandenberghe, J. Van Beeumen, and M. K. Bhat. 2001. Biochemical characterization and mechanism of action of a thermostable β-glucosidase purified from Thermoascus aurantiacus. Biochem. J. 353:
117-127.
21. Perez-Pons, J. A., A. Cayetano, X. Rebordosa, J. Lloberas, A. Guasch, and E. Querol. 1994. A beta-glucosidase gene (bgl3) from Streptomyces sp. strain QM-B814. Molecular cloning, nucleotide sequence, purification and characteriza- tion of the encoded enzyme, a new member of family 1 glycosyl hydrolases. Eur. J. Biochem. 223: 557-565.
22. Pérez-Pons, J. A., X. Rebordosa, and E. Querol. 1995. Pro- perties of a novel glucose-enhanced beta-glucosidase purifi- ed from Streptomyces sp. (ATCC 11238). Biochim. Biophys.
Acta. 1251: 145-153.
23. Pisani, F. M., R. Rella, C. Raia, C. Rozzo, R. Nucci, A.
Cambacorta, D. M. Rosa, and M. Rossi. 1990. Thermostable β-galactosidase from the archaebacterium Sulfolobus sol- fataricus purification and properties. Eur. J. Biochem. 187:
321-328.
24. Rashid, M. H. and K. S. Siddiqui. 1997. Purification and characterization of a β-glucosidase from Aspergillus niger.
Folia Microbiol. 42: 544-550.
25. Romaniec, M. P. M., K. Davidson, and G. P. Hazlewood, 1987. Cloning and expression in Escherichia coli of Clostri- dium thermocellum DNA encoding beta-glucosidase activity.
Enzyme Microb. Technol. 9: 474-478.
26. Shewale, J. G. 1982. 3-Glucosidase: its role in cellulase synthesis and hydrolysis of cellulose. Int. J. Biochem. 14:
435-443.
27. Tamas J., K. Krisztina, S. Zsolt, and R. Kati. 2003. Pro- duction of β-glucosidases in mixed culture of Aspergillus niger BKMF 1305 and Trichoderma reesei RUT C30. Food Technol. Biotechnol. 41: 49-53.
28. Vallmitjana, M., M. Ferrer-Navarro, R. Planell, M. Abel, C.
Ausín, E. Querol, A. Planas, and J. A. Pérez-Pons. 2001.
Mechanism of the family 1 beta-glucosidase from Strepto- myces sp: catalytic residues and kinetic studies. Biochemistry.
40: 5975-5982.
29. Wen, Z., W. Liao, and S. Chen. 2005. Production of cellulase/
β-glucosidase by the mixed fungi culture Trichoderma reesei and Aspergillus phoenicis on dairy manure. Process Biochem.
40: 3087-3094.
30. Yoon, J. J., C. J. Cha, Y. S. Kim, and W. Kim. 2008. De- gradation of cellulose by the major endoglucanase produced from the brown-rot fungus Fomitopsis pinicola. Biotechnol Lett. 30: 1373-1378.
31. Zamocky, M., R. Ludwig, C. Peterbauer, B. M. Hallberg, C.
Divne, P. Nicholls, and D. Haltrich. 2006. Cellobiose de- hydrogenase a flavocytochrome from wood-degrading, phytopathogenic and saprotropic fungi. Curr. Protein Pept.
Sci. 7: 255-280.
(Received Jan. 12, 2009/Accepted May 25, 2009)